大麗輪枝菌及激素處理后棉花酵母雙雜交文庫的構建
2021-04-30 09:03:12許艷李冉宋健王丹周雷李俊嬌張丹丹陳捷胤戴小楓楊星勇
植物保護 2021年2期
許艷 李冉 宋健 王丹 周雷 李俊嬌 張丹丹 陳捷胤 戴小楓 楊星勇

關鍵詞大麗輪枝菌;植物激素;酵母雙雜交;cDNA文庫;互作蛋白
黃萎病是由大麗輪枝菌Verticillium dahliae引起的世界性真菌病害,每年造成棉花、馬鈴薯、蔬菜等農作物損失數十億美元,嚴重威脅農業生產安全。感染黃萎病的植物通常表現出葉片萎蔫、黃化、卷曲或壞死癥狀,維管束變褐,并隨著病程進展最終枯萎死亡。
大麗輪枝菌分泌大量具有毒力的胞外蛋白是成功侵染寄主的重要策略之一,如其分泌的細胞壁降解酶類在侵染寄主過程中通常參與降解細胞壁的作用,表現為毒力功能。另外一類分泌蛋白則參與了病原與寄主互作過程,如大麗輪枝菌的無毒因子Avel能夠被番茄的抗性蛋白Vel識別而誘導寄主免疫反應;大麗輪枝菌中小分子量的富含半胱氨酸蛋白VdSCP41能與寄主植物的轉錄因子CBP60g互作而激發免疫反應。此外,通過在模式植物煙草中瞬時表達大麗輪枝菌分泌的胞外蛋白,還篩選出了多個參與寄主互作的毒力因子,如糖苷水解酶VdEG1和VdEG3、角質酶VdCUTll、小分子量富含半胱氨酸蛋白VdSCP7、壞死和乙烯誘導蛋白NLPs。目前,多個參與病原與寄主互作的效應蛋白已經被鑒定并確定了其在侵染過程中的毒力功能,但其是如何識別寄主或被寄主識別并發揮作用的機制仍不清楚。
酵母雙雜交系統(yeast two-hybrid system,Y2H)是研究蛋白相互作用的常用方法之一,最早在1989年由Fields和Song提出,其原理是利用真核生物轉錄因子的DNA結合結構域-BD和DNA轉錄激活結構域AD,如GAL4和GCN4等,將2個待研究的蛋白分別與BD和AD結構域連接并共同導人酵母菌中,若2個蛋白相互作用則轉錄因子的BD和AD結構域相互靠近,即可以轉錄激活下游報告基因的表達來檢測這種相互作用的發生。除應用于2個已知蛋白間的相互作用的檢測外,酵母雙雜交體系也廣泛應用于與已知蛋白具有潛在相互作用的蛋白篩選。但這一技術在大麗輪枝菌效應子與寄主植物相互作用的研究中很少報道,因此,在明確參與大麗輪枝菌與寄主互作效應子的前提下,通過構建植物寄主酵母雙雜交文庫并篩選與病原效應子互作的靶標蛋白,對揭示大麗輪枝菌侵染寄主過程的致病分子機理具有重要意義。
本研究以大麗輪枝菌及激素處理的棉花為材料,構建了用于大麗輪枝菌效應子互作蛋白篩選的酵母雙雜交文庫,并對文庫的質量進行評估;以實驗室篩選的具有誘導細胞壞死且定位于植物細胞核的效應子VCR1(未發表)為例,通過互作蛋白篩選、基因功能分析和驗證進一步評價了構建的酵母雙雜交文庫的應用價值。本研究建立的棉花酵母雙雜交cDNA文庫將為大麗輪枝菌效應子互作蛋白的篩選鑒定及深入研究病原與寄主互作分子機制的研究提供有力支撐。
1材料與方法
1.1試驗材料
植物材料:海島棉‘海7124Gossypium barba-dense‘Hai 7124,生長條件為L∥D=14 h∥10h,溫度28℃。
菌株:落葉型強致病力大麗輪枝菌Vd991、酵母菌株Y187和Y2H均由本實驗室保存,大腸桿菌感受態細胞Transl-T1購自北京全式金生物技術有限公司。
質粒:酵母雙雜交所用的誘餌質粒pGBKT7、文庫質粒pGADT7、對照質粒pGBKT7-Lam、pGBKT7-53和pGADT7-T購自美國賽默飛公司。
1.2試驗方法
1.2.1海島棉樣品處理及RNA提取
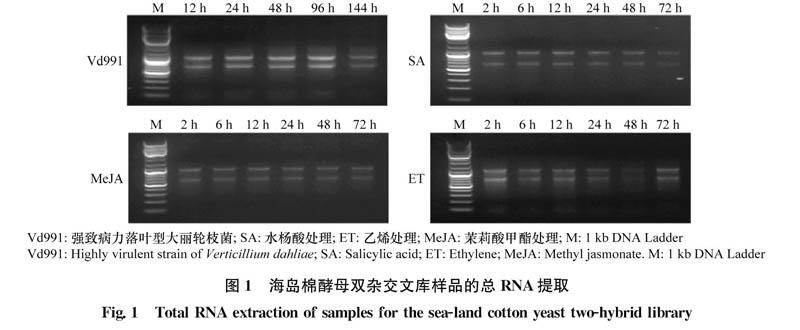
大麗輪枝菌于馬鈴薯葡萄糖瓊脂(potato Dex-trose agar,PDA)培養基25℃條件下培養7d,無菌水收集孢子并制備成2×10個/mL孢子懸浮液。取20株2周齡海島棉幼苗(2片真葉完全展平)浸泡于50mL上述孢子懸浮液,30min后移栽回含有滅菌土的營養缽,處理后分別于12、24、48、96 h和144 h取根部樣品液氮速凍,-80℃保存。制備10 mmol/L水楊酸(salicylic acid,SA)、5 mmol/L乙烯利溶液(ethephon,ET)和100mol/L茉莉酸甲酯(methyl jasmonate,MeJA),其中乙烯利用水溶,水楊酸和茉莉酸甲酯用乙醇促溶后用水稀釋到目標濃度,3種激素分別噴施于20株上述海島棉幼苗,進行單獨處理,處理后于2、6、12、24、48 h和72 h后取葉片迅速置于液氮中速凍,-80℃保存。RNA提取使用EASYspin植物RNA快速提取試劑盒(北京原平皓生物技術有限公司),通過1.0%瓊脂糖電泳檢測提取RNA的完整性,使用Nano-Drop2000檢測提取RNA的質量與濃度。
1.2.2酵母雙雜交cDNA文庫構建
根據檢測濃度將大麗輪枝菌和激素處理的所有植株RNA樣品等量混合,利用SMARTTM cDNA文庫構建試劑盒(美國賽默飛公司)構建酵母雙雜交cDNA文庫。以上述2μL的RNA為模板合成第1條cDNA鏈,然后通過LD-PCR擴增cDNA得到第2條鏈,并利用柱層析剔除250 bp以下的雙鏈cDNA。利用PEG/LiAc法將所得cDNA產物和6μL的pGADT7-Rec線性質粒共轉入Y187酵母感受態細胞內,用15mL的NaCl溶液(0.9%)重懸菌體并涂布于缺少亮氨酸(Leu)的酵母培養基(syn-thetic dropout nutrient medium,SD)SD/-Leu培養基,30℃倒置培養3~4d后收集菌體,得到的酵母細胞即為處理樣品的酵母雙雜交cDNA文庫,按1.0mL/管分裝于1.5mL離心管中,液氮速凍存于μ80℃備用。
1.2.3 cDNA文庫轉化效率及滴度檢測
從上述重懸菌液吸取100μL并用0.9%NaCl10倍梯度稀釋至10,涂布于SD/-Leu培養基后于30℃條件下倒置培養3~4d,通過單菌落數計算轉化效率。將無菌水收集的菌體稀釋至10、10、10和10,分別吸取50,μL涂布于SD/-Leu培養基上,30℃倒置培養3d,根據單菌落數計算文庫滴度。
1.2.4cDNA文庫平均插入片段大小及重組率分析
在SD/-Leu平板上隨機挑取23個單菌落,使用上游引物(5,-TAATACGACTCACTATAGGGO3)和下游引物(5-AGATGGTGCACGATGCA-CAG-3)進行菌落PCR擴增。擴增條件:95℃預變性3 min;95℃變性30s,55℃退火30 s,72℃延伸2min,擴增34個循環;72℃延伸5min。擴增產物使用1.0%的瓊脂糖凝膠電泳檢測,依據擴增產物片段大小計算cDNA文庫的平均插入片段大小,陽性克隆比率以及cDNA文庫的重組率。
1.2.5誘餌載體構建
以實驗室篩選的定位于寄主植物細胞核、引起細胞壞死的大麗輪枝菌效應蛋白VCR1(未發表)為研究對象,構建誘餌載體并開展酵母雙雜交文庫的篩選。利用帶EcoR I酶切位點的特異性引物(上游引物:5,-CATATGGCCATGGAGGC-CGAATTCATGGCACCAGTCCCGGCT-3;下游引物:5,-AGGTCGACGGATCCCCGGGAATTCG-CATGTGTTGAGCATGCT-3)對VCR1基因進行PCR擴增,EcoR I對誘餌表達載體pGBKT7酶切消化,再通過ClonExpress II One Step Cloning Kit(諾唯贊,南京)將VCR1基因片段連接到pGBKT7上;獲得重組質粒pGBKT7-VCRl轉入Trans1-T1大腸桿菌感受態細胞,使用pGBKT7載體通用引物(上游引物:5,-TAATACGACTCACTATAGGGC-3;下游引物:5,-TAAGAGTCACTTTAAAATTT-GTAT-3)進行PCR擴增驗證。
1.2.6重組誘餌載體自激活及毒性檢測
將上述構建的誘餌載體pGBKT7-VCR1及pGBKT7質粒分別轉入Y2H酵母感受態細胞,涂布于SD/-Trp、SD/-Trp-His和SD/-Trp-His-Ade培養基,通過轉化后酵母細胞在上述培養基上的生長情況檢測VCR1的自激活性。將上述轉化體系涂布SD/-Trp和酵母膏胨葡萄糖瓊脂(YPDA)培養基,通過觀察其在平板上生長情況鑒定VCR1對酵母是否具有毒性。
1.2.7酵母雙雜交文庫的篩選
酵母文庫篩選:上述攜帶有誘餌載體的菌株于30 mL SD/-Trp液體培養基中擴大培養,離心收集菌體并用5mL的SD/-Trp液體培養基重懸,隨后與構建的酵母雙雜交文庫混勻,并于45mL的2×YPDA培養基(卡那霉素,50μL/L)中混勻融合培養,30℃、45r/min振蕩培養20h。待觀察到有合子出現后繼續培養4h,典型的合子呈現三耳突狀,分別代表兩個母細胞和一個子細胞。之后收集菌體,用0.5×YPDA(卡那霉素,50μL/L)培養液重懸菌體2次,最后定容至10mL。將上述重懸的酵母細胞涂布于直徑15cm的SD/-Trp-Leu-His(TDO)培養基上,每個涂布200μL,共50個平板,30℃倒置培養3~5d。將TDO培養基上長勢較好的陽性克隆轉接到SD/-Trp-Leu-His-Ade(QDO)培養基上,30℃倒置培養觀察其生長隋況。
結合效率測定:取上述重懸的酵母菌體100μL,按10倍梯度稀釋至10后每個梯度都分別涂布于SD/-Trp、SD/-Leu、SD/-Trp-Leu(DDO)培養基平板上,每種培養基涂5個平板,30℃倒置培養3~5d。待長出菌落后,統計二倍體、文庫和誘餌克隆數,在相同稀釋倍數的前提下,計算結合效率。結合效率=二倍體克隆數/文庫或誘餌克隆數×100%。
潛在互作轉化子測序:挑取在SD/-Trp-Leu-His-Ade培養基上正常生長的酵母細胞,于SD/-Trp-Leu-His-Ade液體培養基進行擴大培養,利用pGADT7載體通用引物(上游引物:5,-TAATAC-GACTCACTATAGGGC-3;下游引物:5,-AGAT-GGTGCACGATGCACA-3)擴增轉化子的cDNA插入片段并測序分析。
1.2.8潛在互作蛋白基本特性分析
對上述獲得的潛在互作轉化子序列進行初步生物信息學分析:通過NCBI數據的BlastP比對推測潛在互作蛋白的編碼序列;利用SMART(http://smart.embl-heidelberg.de/smart/set-mode.cgi?NORMAL=1)預測編碼蛋白的保守結構域;采用WoLF PSORT(https://wolfpsort.hgc.jp/)預測編碼蛋白的亞細胞定位;通過SignalP 4.1(http://WWW.cbs.dtu.dk/services/SignalP-4.1/)預測編碼蛋白是否具有信號肽序列特征。
1.2.9互作蛋白驗證
對篩選的陽性克隆與本研究的大麗輪枝菌效應蛋白VCR1進行驗證:隨機選取5個可能定位于細胞核的潛在互作基因,合成全長cDNA序列并構建于pGADT7載體上,將其分別與誘餌載體pGBKT7-VCRl共轉化至Y2H酵母感受態細胞中,以pGADT7-T和pGBKT7-53、pGADT7-T和pGBKT7-Lam分別作為陽性和陰性對照,涂布SD/-Trp-Leu(DDO)培養基,30℃倒置培養3~5d。觀察SD/-Trp-Leu培養基上酵母的生長情況,將陽性克隆轉接到SD/-Trp-Leu-His-Ade(QDO)和SD/-Trp-Leu-His-Ade/X-a-gal(QDO/x)培養基上,30℃倒置培養5~7d后觀察生長和顯色情況,驗證篩選的基因與靶標效應蛋白的互作情況。
2結果與分析
2.1構建酵母雙雜交文庫的樣品處理及RNA提取
‘海7124是抗黃萎病棉花品種,落葉型強致病力大麗輪枝菌Vd991侵染5~6d后病原仍停留在皮層,病原菌無法繁殖擴展;同時,激素SA、ET和MeJA處理下可誘導植物抗病相關基因表達。為構建有效的抗病相關基因誘導表達文庫,本研究收集了大麗輪枝菌Vd991侵染以及激素SA、ET和MeJA處理的‘海7124樣品,并提取了所有樣品的總RNA,1.0%瓊脂糖凝膠電泳檢測結果表明28S及18S條帶清晰,完整性好。依據紫外吸收光譜檢測發現上述提取的RNA樣品的ODe60/ODe80在1.97~2.02之間,濃度在500.4~2 086.4ng/μL(圖1),表明RNA質量較好,可以用于文庫構建。
2.2海島棉酵母雙雜交eDNA文庫構建及評價
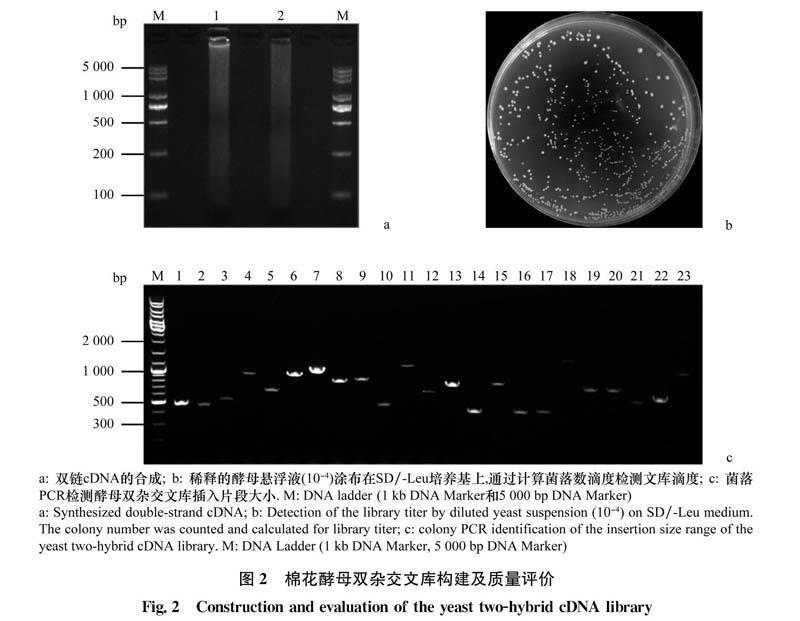
上述4組處理樣品(大麗輪枝菌侵染及3種激素處理)的總RNA等量混合,反轉錄合成cDNA第一鏈,使用此cDNA合成雙鏈cDNA(圖2a)并利用柱層析剔除250 bp以下的雙鏈cDNA(ds-cDNA)。將獲得的ds-cDNA與酵母雙雜交文庫載體pGADT7獲得共轉化Y187酵母菌株進行同源重組,最終獲得海島棉在上述4種處理條件下的酵母雙雜交文庫。用SD/-Leu培養基收集cDNA文庫的菌體,得到約130mL菌懸液并分裝于1.5 mL離心管,-80℃凍存備用。
文庫轉化完成后,通過梯度稀釋涂布SD/-Leu培養基對酵母文庫的轉化效率和滴度進行檢測。通過統計,稀釋至10平板平均菌落數為590個(圖2b),計算得酵母文庫的轉化效率為5.9×10/3μgpGADT7-VCR1,高于1×10/3μg pGADT7-Rec的試驗要求,并且文庫滴度達到1.18×10cfu/mL,同樣高于1×10cfu/mL的文庫構建標準。為驗證本研究酵母文庫插入片段情況,從上述計算轉化效率的平板中隨機挑取23個菌落進行PCR擴增,結果顯示,挑取的克隆菌落均可擴增出插入片段,重組率達100%,并且擴增片段大小分布在300~2000bp之間(圖2c)。綜上結果說明,本研究構建的大麗輪枝菌及激素處理下棉花酵母雙雜交cDNA文庫質量良好,可用于后續大麗輪枝菌效應蛋白的篩選鑒定。
2.3大麗輪枝菌效應蛋白VCR1誘餌載體構建
使用pGBKT7通用引物對連接轉化后平板上的陽性克隆進行分子鑒定,并以空載體為對照。結果顯示,構建的誘餌載體擴增出的條帶與空載體擴增出的條帶差值大小與VCR1片段大小相近,約580bp,進一步提取構建好的誘餌載體質粒,用EcoRI內切酶進行酶切驗證,并進行測序,結果與預期一致,表明大麗輪枝菌VCR1基因成功整合到誘餌載體pGBKT7(pGBKT7-VCR1)(圖3a,3b)。為驗證VCR1的自激活性,將pGBKT7-VCR1轉化至Y2H酵母并涂布于相應培養基上,結果顯示轉化pGBKT7-VCR1誘餌載體與pGBKT7載體的酵母在SD/-Trp培養基上正常生長,在SD/-Trp-His和SD/-Trp-His-Ade培養基上不生長(圖3c),表明VCRl基因在酵母菌株中沒有自激活性。同時,含有pGBKT7-VCR1誘餌載體和pGBKT7載體的酵母菌株在YPDA培養基上生長一致(圖3d),表明VCR1對Y2H酵母菌株沒有毒性。因此,大麗輪枝菌VCR1可以用于下一步與酵母雙雜交文庫互作蛋白的篩選。
2.4大麗輪枝菌效應蛋白VCR1的候選互作蛋白篩選
使用上述誘餌載體pGBKT7-VCR1篩選文庫,經統計SD/-Leu培養基上稀釋100倍的菌落數為415個,1000倍的48個,SD/-Trp-Leu培養基上稀釋10倍的菌落數為55個,100倍的10個,計算得出酵母雙雜交結合效率約為1.33%(圖4a)。進一步從SD/-Trp-Leu-His培養基上篩選出生長正常的克隆480個接種于SD/-Trp-Leu-His-Ade培養基上二次篩選,最終獲得生長正常的克隆422個,使用pGADT7通用引物對422個克隆進行PCR鑒定,獲得插入片段大于500 bp的克隆201個(圖4b)。
2.5大麗輪枝菌效應蛋白VCR1的候選互作蛋白驗證
根據上述克隆測序結果分析發現,本研究共篩選到了與大麗輪枝菌效應蛋白VCRl潛在互作的蛋白27個(表1)。通過序列比對與功能注釋表明,其中18個為假定蛋白(功能未知)。結果顯示,效應蛋白VCR1與其中4個潛在互作蛋白(VCR1-IP001、VCR1-IP005、VCR1-IP009和VCR1-IP027)與陰性對照(pGADT7-T+pGBKT7-Lam)僅在SD/-Trp-Leu培養基上正常生長,在SD/-Trp-Leu-His-Ade和SD/-Trp-Leu-His-Ade/X-a-gal培養基上皆不生長;而效應蛋白VCRl和VCRl一IP003與陽性對照(pGADT7-T+pGBKT7-53),在SD/-Trp-Leu、SD/-Trp-Leu-His-Ade和SD/-Trp-Leu-His-Ade/X-a-gal培養基上均能夠正常生長,表明VCRl可以與VCR1-IP003互作(圖5)。
3討論
植物在病原菌侵染時會誘導抗性基因表達來抵御病原菌的侵染。此外,植物激素也可以作為免疫信號分子激活防御反應,如茉莉酸、水楊酸、乙烯等。水楊酸能夠激活針對活體、半活體營養寄生物的抗性防衛反應;茉莉酸能夠與乙烯協同發揮作用,激活對死體營養病原菌的防御反應。定位于細胞膜上的棉花枯草桿菌酶(subtilase)GbSBT1同時受大麗輪枝菌、茉莉酸和乙烯誘導,并進入細胞質中發揮功能。因此,參與病原與寄主互作的基因往往在病原侵染、激素處理條件下被大量誘導表達。海島棉對黃萎病具有較高的抗性,部分品種對黃萎病菌免疫,與陸地棉相比,更適用于篩選抗病基因;‘海7124是抗病基因鑒定與功能研究的常用材料。因此,本研究綜合采用大麗輪枝菌侵染、激素處理等條件誘導‘海7124抗病相關基因表達,對處理的樣品提取RNA并混合構建酵母雙雜文庫。本研究所構建的文庫可以覆蓋多種誘導條件下的抗病相關基因,文庫抗性相關基因種類和豐度優于單一處理條件下構建的酵母雙雜交文庫,從而提高了篩選到與大麗輪枝菌效應子互作蛋白的概率。應用本研究構建的酵母文庫,成功篩選到誘餌蛋白VCR1的27個潛在互作蛋白。
高質量cDNA文庫構建是利用酵母雙雜交篩選互作蛋白的關鍵,酵母文庫的好壞直接影響互作蛋白的篩選效率。良好的酵母文庫需要有較高的轉化效率(>1×10。/3μg pGADT7-Rec)、重組率、較長的插入片段以及足夠的文庫滴度(>1×10cfu/mL)。有研究報道利用水稻構建的酵母雙雜交文庫,轉化效率達到6.65×10/3μg pGADT7-Rec,重組率為100%。經質量檢測,本研究構建酵母文庫轉化效率為5.9×10/3μg pGADT7-VCRl,連接片段檢測重組率為100%,且插入片段分布在300~2000bp;此外,本研究構建的酵母雙雜交cDNA文庫滴度為1.18×10cfu/mL,約為標準量的10倍。綜上結果,本研究利用大麗輪枝菌及激素處理下的混合樣品,成功建立了高效的酵母雙雜交cDNA文庫,這為后續篩選與大麗輪枝菌效應子互作的靶標蛋白提供了重要的材料基礎。
酵母雙雜交系統是在真核酵母細胞內研究蛋白之問相互作用的一種方法,此方法靈敏度高,應用廣泛。本研究應用酵母雙雜交技術成功篩選到27個與VCR1潛在互作的蛋白,這表明構建的酵母雙雜交文庫具有較好的篩選效果。利用棉花測序的基因組信息,分析獲得對應基因的全長序列并進行功能預測,發現鑒定的27個潛在互作蛋白中,18個為未知蛋白,其功能尚不明確;其中24個蛋白質均無信號肽。16個潛在互作蛋白預測定位于細胞核,與效應子VCR1在寄主植物的亞細胞定位(細胞核)相同,具有很高的可信度。隨機選取5個定位于細胞核的潛在互作蛋白進行回轉驗證,確證了與效應子VCR1互作的候選蛋白(VCR1-IP003)。上述結果一定程度上客觀反映了酵母雙雜交技術自身存在假陽性高的局限性,但也說明本研究構建的酵母雙雜交文庫可以有效應用于病原與寄主互作蛋白的篩選。
綜上,本試驗成功構建了海島棉‘海7124在大麗輪枝菌侵染和激素處理下的酵母雙雜交cDNA文庫,文庫具有較高的轉化效率和重組效率,并通過大麗輪枝菌效應子VCR1互作蛋白的篩選系統評價了構建的棉花酵母雙雜交文庫,為后續篩選與大麗輪枝菌胞外蛋白(效應子)互作的靶標蛋白及其互作機理研究奠定了基礎。
