竹類植物分蘗相關基因的生物信息學分析
2021-05-06 08:45:58孫佳瑋郭振華
西南林業大學學報 2021年2期
孫佳瑋郭振華
(1.中國科學院昆明植物研究所,中國西南野生生物種質資源庫,云南 昆明 650201;2.中國科學院大學,北京 100049)
竹亞科(Bambusoideae)屬于禾本科(Poaceae)的12個亞科之一,共125屬1 670種,可分為草本竹與木本竹兩大類[1]。竹類的地下莖俗稱竹鞭,形態特征較穩定,不僅是竹亞科分類的重要依據之一,而且對竹類的生長發育起著關鍵的營養運輸的作用。根據竹類地下莖營養器官繁殖特點和形態特征可劃分為粗型和細型。粗型地下莖即合軸型,在地下沒有竹鞭,母竹側芽直接發育成竹,地上部表現為密集叢生;細型地下莖即單軸型,母竹側芽發育成竹鞭,且鞭上側芽可以再發育成筍或鞭,在地上表現為散生狀[2],形成龐大的地下莖網絡。粗型地下莖的竹類生長模式與禾本科其他類群的分蘗生長有相似之處,而細型地下莖的竹類則不產生或少量分蘗,形成獨特的地下莖系統。因此,探究竹子不同類型的地下莖的形成與禾草植物的分蘗之間的關系將有助于對竹類地下莖發育模式的理解。
前人分別在雷竹(Phyllostachys praecox)、美麗箬竹(Indocalamus decorus)等單一竹種中的研究表明,ABA、GA3、ZT、IAA等激素參與了竹類地下莖側芽的萌發過程[3-4],但相關基因的研究尚未見報道。在模式植物水稻中的分蘗相關基因已有了較系統的研究,且有研究表明,某些分蘗相關基因參與到這些激素的信號通路中,最具代表性的就是MOC1基因,其編碼的蛋白產物包含VHIID和SH2?like結構域,屬于植物特有的GRAS家族蛋白,參與了植物側枝形成、根輻射方向結構分布、光信號傳導、植物激素赤霉素的信號傳導和植株高度控制等多種重要的生理和發育過程[5]。因此本研究從水稻(Oryza sativa)中選取相關的候選基因,對毛竹(Phyllostachys heterocycla)和4種新測序的竹類植物[6]以及水稻、谷子(Setaria italica)和二穗短柄草(Brachypodium distachyum)的分蘗數相關基因和氨基酸序列進行比較研究,發現其中差異從而為后續不同地下莖類型的相關研究提供參考。
1 材料與方法
1.1 材料介紹
現已有5種竹子的基因組完成測序,分別代表了竹類的2種地下莖類型(圖1)和所有的倍性:地下莖為細型的四倍體毛竹;地下莖為粗型的四倍體瓜多竹(Guadua angustifolia)和六倍體蕓香竹(Bonia amplexicaulis),以及二倍體草本竹Olyra latifolia和Raddia guianensis。同時選取了模式植物水稻,以及禾本科的二穗短柄草和谷子為參考。
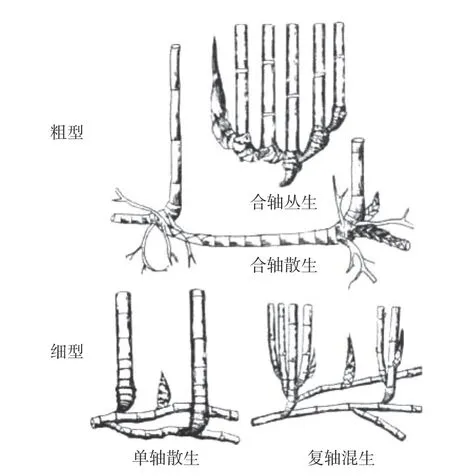
圖 1 木本竹地下莖類型[7]Fig.1 Rhizome types of woody bamboo
1.2 竹類分蘗基因的鑒定及序列分析
在funRiceGenes網站(https://funricegenes.github.io/)找到了51個與水稻分蘗數相關的基因,通過NCBI數據庫(https://www.ncbi.nlm.nih.gov/)以 及Rice Genome Annotation Project(http://rice.plantbiology.msu.edu/)對這51個基因進行再次確認。將以上基因作為候選基因,在NCBI網站使用BLAST工具,用blastp與blastn方法,在5種竹子的基因組中獲取這些基因的同源序列,將獲得的同源序列進行建樹、篩選,從而更準確獲取同源序列,用DNAMAN軟件對其DNA和氨基酸序列進行比對。
1.3 進化樹構建與同源序列分析
使用MEGA X軟件對水稻、二穗短柄草、谷子、毛竹、瓜多竹、蕓香竹、草本竹Olyra latifolia和Raddia guianensis的各基因比對結果進行分析,運用最大似然法(ML),設置1 000次檢驗構建進化樹。候選基因的氨基酸序列的分子量、等電點、序列長度、脂溶性系數等信息通過網站(http://www.expasy.org)獲取。在MEME(http://meme-suite.org/index.html)網站對候選基因進行motif預測,并通過TBtools軟件作基因結構圖。
2 結果與分析
2.1 基因比對結果
從獲得的51個與水稻分蘗數相關的基因,通過序列比對分析發現其中17個基因在竹類與水稻、二穗短柄草及谷子存在明顯差異,包括:D10、DLT、Gn1a、HTD1、LAX1、MOC1、Os-AAP3、OsCYP19?4、OsEATB、OsIAGLU、OsPIN1、OsPIN5b、OsSPL14、OsSUT2、OsTB1、RFL、THIS1。基因序列以Osa代表水稻,Bdi代表二穗短柄草,Sit代表谷子,Ped代表毛竹,Bam代表蕓香竹,Gan代表瓜多竹,Ola代表Olyra latifolia,Rgu代表Raddia guianensis。(正文中只列出D10、OsCYP19?4、OsEATB、DLT、Gn1a、LAX1、MOC1、OsPIN5b、OsSPL14、OsTB1等部分基因,其余請見電子附圖1~7。)
2.2 基因功能分類
2.2.1 使分蘗數增加的基因
竹子D10基因的氨基酸序列信息如表1,D10基因的氨基酸序列比對結果如圖2所示,相似度為77.22%。D10基因的進化樹(圖3)及motif信息(表2)如下。motif 1、2、3、4、5、9、10對應類胡蘿卜素裂解雙加氧酶的編碼區。草本竹Olyra latifolia和蕓香竹的序列較為保守;RguD10b序 列 缺 失motif 4、5、6、7,毛竹PedD10a序列缺失motif 4、5,PedD10b序列缺失motif 4、6、7,這3條序列較其他序列在motif 2前多1個motif 8;瓜多竹的兩條序 列缺少motif 3。D10基因編碼類胡蘿卜素裂解雙加氧酶(CCD8),參與獨角金內酯(SL)的生物合成以及側根、側芽的萌發,在根中柱、葉芽、莖、花序和圓錐花序的木質部薄壁細胞中表達[8],植株表現為矮化,分枝和分蘗數增加。D10蛋白定位于葉綠體。Raddia guianensis和毛竹序列中所缺失的片段為CCD8的功能域。
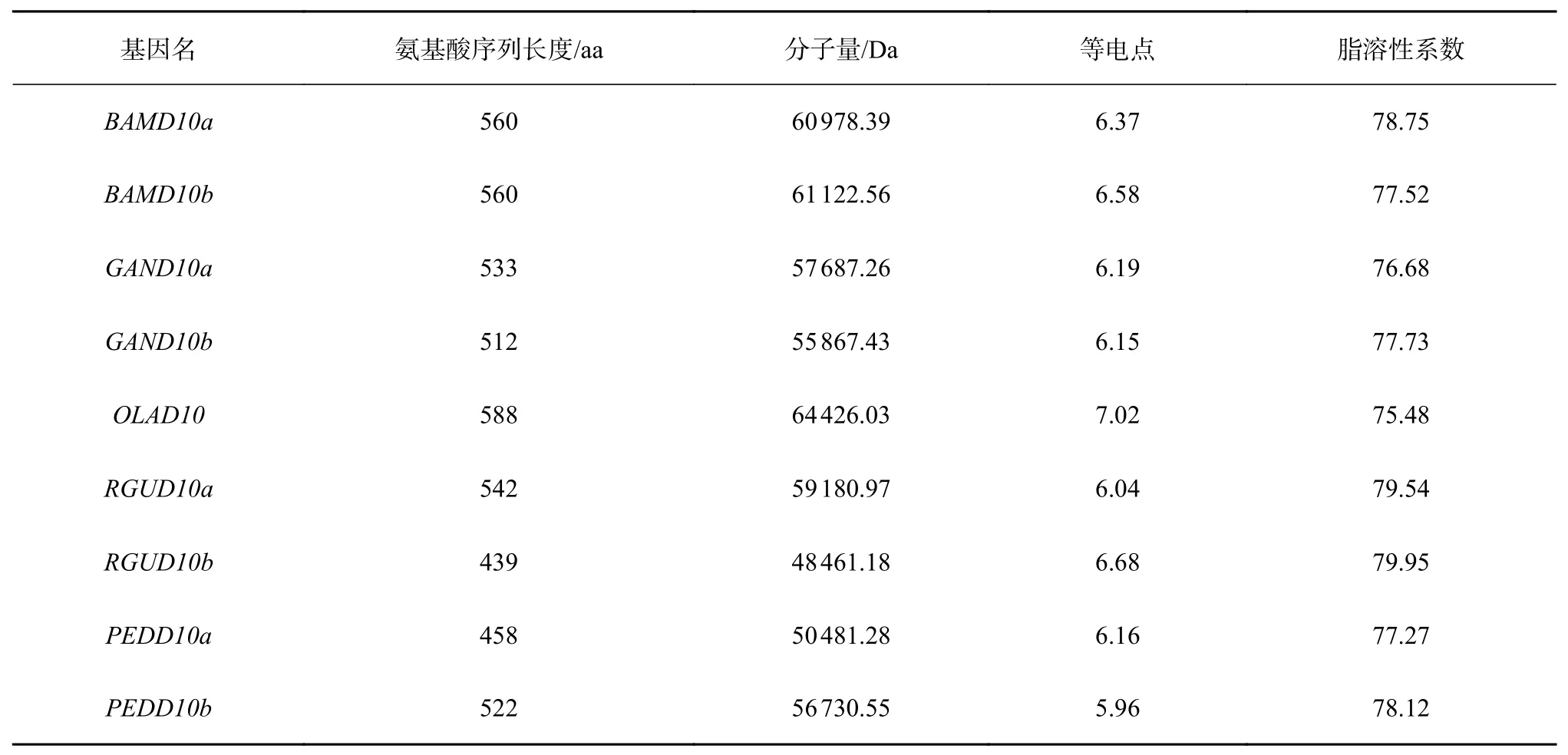
表 1 竹子D10基因的氨基酸序列信息Table 1 Amino acid sequence information of bambooD10gene

圖 2D10基因的氨基酸序列比對結果Fig.2 Amino acid sequence alignment results ofD10gene

圖 3D10基因的進化樹及結構域Fig.3 Evolutionary tree and motif ofD10gene

表 2D10基因的motif信息Table 2 Motif information ofD10gene
OsCYP19?4基因的氨基酸序列比對結果顯示相似度為21.55%。竹子的氨基酸序列信息如表3。OsCYP19?4基因進化樹及motif如下(圖4)。motif 2、3、5對應的結構域為Pro-isomerase。除水稻和Raddia guianensis外,其他序列中都存在motif5,草本竹Olyra latifolia、Raddia guianensis、瓜多竹、蕓香竹以及毛竹PedCYP19?4c序列缺失motif 7。OsCYP19?4編碼一種包含208個氨基酸的蛋白,編輯的肽脯氨酰順反異構酶加速蛋白質折疊,催化脯氨酸酰亞胺肽鍵在寡肽中的順反式異構化,基因過表達使分蘗增加[9],此結構域在竹類中較為保守。
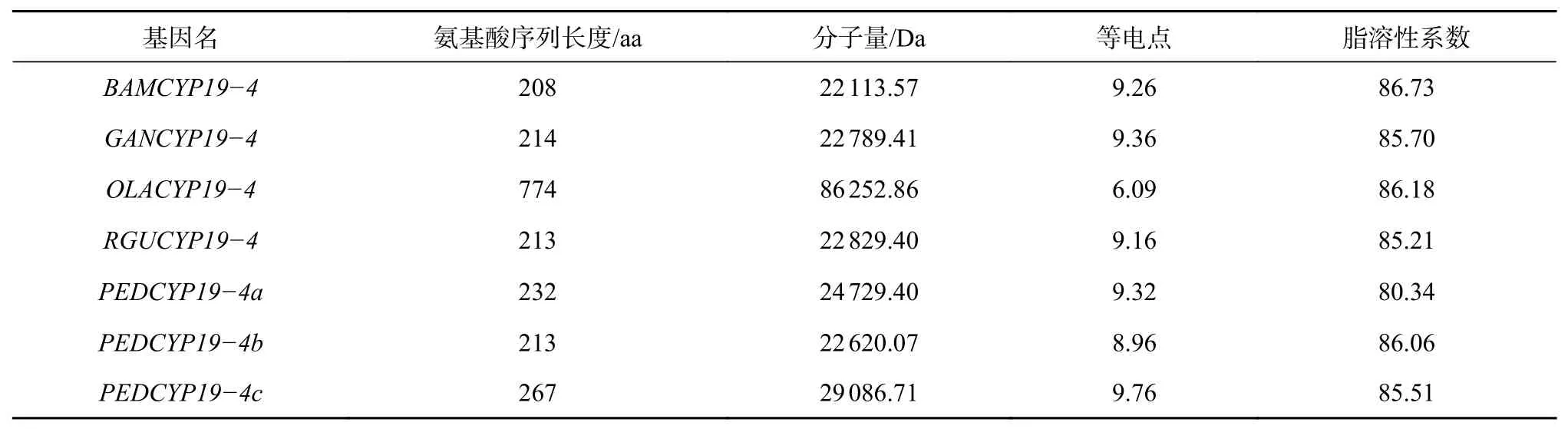
表 3 竹子OsCYP19?4氨基酸序列信息Table 3 BambooOsCYP19?4amino acid sequence information

圖 4OsCYP19?4基因的進化樹、motif及基因結構Fig.4 Evolutionary tree,motif and gene structure ofOsCYP19?4gene
OsEATB基因的氨基酸序列的比對結果顯示相似度為63.20%。竹子的氨基酸序列信息如表4。OsEATB基因的進化樹及motif如下(圖5)。motif 1對 應AP2結 構 域。motif 1、3、4、5在8個物種中保守。竹類中都包含motif 7,其余3種中不存在。毛竹和蕓香竹中有motif 6,Olyra latifolia的motif 1和5之 間 有motif 10插 入,缺失motif 9。OsEATB基因降低成熟時的株高[10],促進分蘗和小穗分枝發生。

表 4 竹子OsEATB氨基酸序列信息Table 4 BambooOsEATBamino acid sequence information

圖 5OsEATB基因的進化樹、motif及基因結構Fig.5 Evolutionary tree,motif and gene structure ofOsEATBgene
OsPIN1基因的進化樹及motif見電子附圖1。除毛竹外,其他4種竹類的序列與水稻、二穗短柄草、谷子的差異極小,毛竹的PedOsPIN1a序列缺失motif 3、4、7、9,PedOsPIN1b序列只有motif 3、4、8。OsPIN1基因在維管組織和根原基中表達,參與分蘗的產生,低表達時會使分蘗數增加[11]。
OsSUT2基因的進化樹及motif見電子附圖2。此基因在水稻和竹類中相似性較高,motif 1、3、4在8個物種中均保守,motif 5、7在除瓜多竹和草本竹Raddia guianensis外的幾個物種中保守。Raddia guianensis、瓜多竹和毛竹中均無motif 2和6。OsSUT2基因編碼的蛋白質參與蔗糖代謝的途徑,并且減少分蘗的產生[12]。
RFL基因的進化樹及motif見電子附圖3。蕓香竹BamRFLb序列缺失motif 4,瓜多竹序列缺失motif 1和6,在末端多一段motif 9;草本竹Raddia guianensis缺失motif 6、7、8、10,毛竹序列中缺失motif 8、10。RFL基因與開花相關,除調控水稻抽穗期、穗部枝梗發育外,它還可以通過調節一些激素信號分子和轉錄因子等影響分蘗的形成[13]使分蘗數減少。毛竹缺失的部分為轉錄因子區域,包含甘氨酸多聚體。

圖 6DLT基因的氨基酸序列比對結果Fig.6 Amino acid sequence alignment results ofDLTgene
2.3 使分蘗數減少的基因
DLT基因的氨基酸序列比對結果如圖6所示,相似度為83.68%。竹子的氨基酸序列信息如表5。DLT基因的進化樹及motif如下(圖7,表6)。motif 1、2、4、6、7、8、9對應GRAS結構域。此基因在8個物種中均較為保守,只有毛竹序列缺少motif 4。DLT基因編碼一個由617個氨基酸組成植物特有的GRAS家族蛋白,包含leucine heptad Ⅰ、VHIID、leucine heptad Ⅱ、PFYRE和SAW等結構域,是少分蘗矮稈基因[14]。毛竹序列中缺失的片段為PFYRE基序。

表 5 竹子DLT氨基酸序列信息Table 5 BambooDLTamino acid sequence information
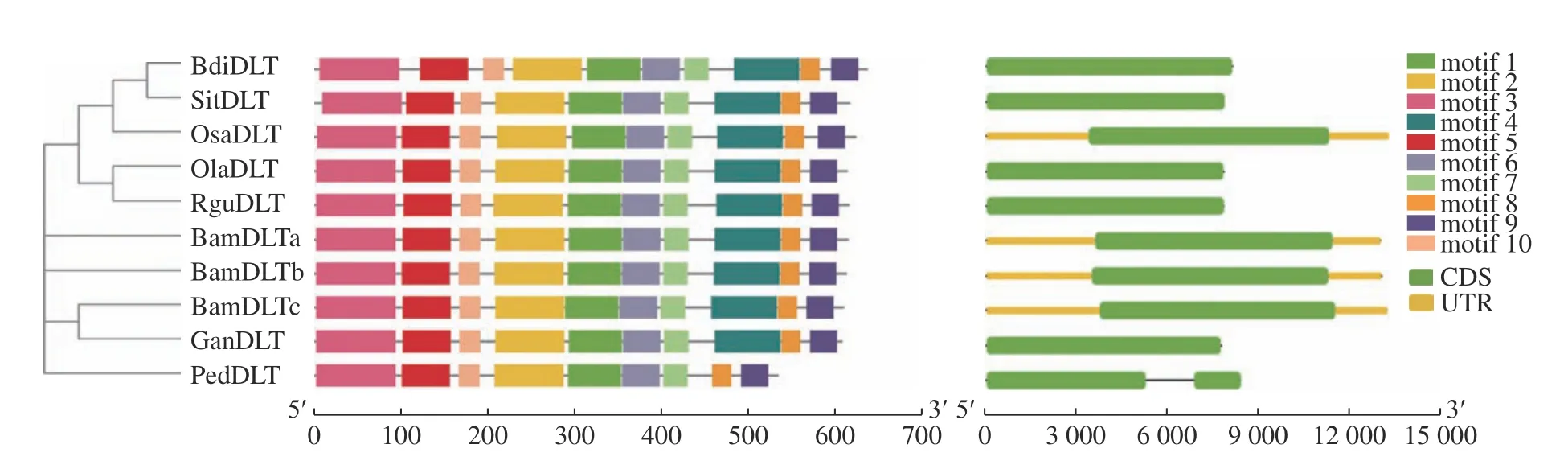
圖 7DLT基因的進化樹、motif及基因結構Fig.7 Evolutionary tree,motif and gene structure ofDLTgene

表 6DLT基因的motif信息Table 6 Motif information ofDLTgene
MOC1基因的氨基酸序列比對結果顯示相似度為48.88%。竹子的氨基酸序列信息如表7。MOC1基因的進化樹及motif如下(圖8)。motif 1、2、3、4對應的結構域為GRAS,motif 5對應一段信號肽。該基因在竹子與水稻中有較高的相
似性,但Raddia guianensis的motif缺失很多。毛竹序列中缺少motif 4和5。MOC1定位于水稻第6號染色體的長臂,編碼GRAS蛋白,為單分蘗基因,不產生分蘗,只有一個主莖[5]。

表 7 竹子MOC1氨基酸序列信息Table 7 BambooMOC1amino acid sequence information
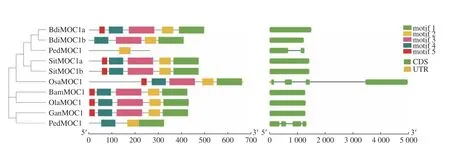
圖 8MOC1基因的進化樹、motif及基因結構圖Fig.8 Evolutionary tree,motif and gene structure ofMOC1gene
LAX1基因的氨基酸序列比對結果顯示相似度為36.85%。竹子的氨基酸序列信息如表8,LAX1基因的進化樹及motif見電子附圖4。motif 6對應HLH(helix-loop-helix)結 構;motif 2、3在8種植物中都保守,水稻、谷子、二穗短柄草中無motif7而竹類中均存在。蕓香竹BamLAX1b序列缺失motif 4、6,Raddia guianensis序列缺失motif 1、4、6,缺失HLH結構。LAX1基因編碼植物特有的bHLH轉錄因子,對基因的轉錄發揮調控作用,可以控制水稻的腋芽原基形成,減少分枝分蘗的產生[15]。

表 8 竹子LAX1氨基酸序列信息Table 8 BambooLAX1amino acid sequence information
Gn1a基因的氨基酸序列比對結果顯示相似度為67.42%。竹子的氨基酸序列信息如表9。Gn1a基因的進化樹及motif見電子附圖5。motif 2、6、9對應FAD結合區,motif 1、3、4、5、7、8為Cytokin(細胞因子)結合區。motif 1、3、4在8個物種中保守。瓜多竹GanGn1a?1序列缺失motif 8,GanGn1a?2序 列 缺 失motif 5、6、9、10,Raddia guianensis序 列 缺 失motif 6、8、9,毛 竹PedGn1a?1序 列 缺 失motif 2、6、9、10,PedGn1a?2序列缺失motif 6、9、10。Gn1a基因在葉、莖及花和花序分生組織中表達,另外可調控減少分蘗產生,基因沉默會使分蘗增加[16]。

表 9 竹子Gn1a氨基酸序列信息Table 9 BambooGn1aamino acid sequence information
OsPIN5b基因的氨基酸序列比對結果顯示相似度為51.89%。竹子的氨基酸序列信息如表10。OsPIN5b基因的進化樹及motif見電子附圖6。motif 3、6對應一個Mem_trans結構域。motif 1、2、3、4在所有序列中保守。毛竹序列和BamPIN5b?1序列缺失motif 2、5、6。OsPIN5b基因過表達會使分蘗減少,降低表達量使分蘗增加,可能作為生長素外排載體的組成部分,在根和莖頂端表達[17]。

表 10 竹子OsPIN5b氨基酸序列信息Table 10 BambooOsPIN5bamino acid sequence information
OsSPL14基因的氨基酸序列比對結果顯示相似度為63.82%。竹子的氨基酸序列信息如表11。OsSPL14基因的進化樹及motif見電子附圖7。motif 1對應的結構域為SBP,含有1個鋅指結構。motif 2、4、5、6、7、8在所有序列中保守,草本竹Raddia guianensis和毛竹與其他物種的序列差異較大,缺失motif 1、3、9,無SBP結構域。OsSPL14基因可能與穗的發育有關,使分蘗數減少[18]。起始84個氨基酸序列為富含甘氨酸區域,毛竹缺失的為此結構域。所有序列中都保守的區域為雙組分核定位信號區。
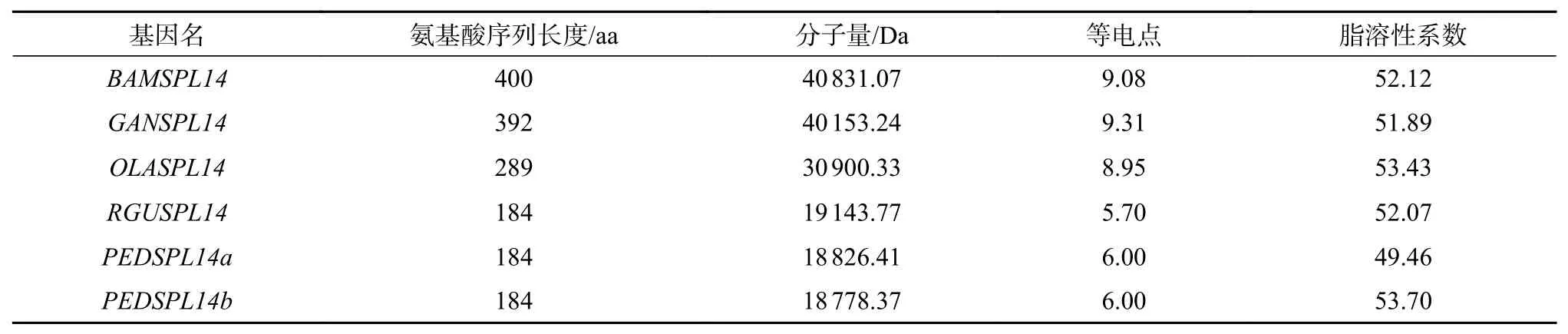
表 11 竹子OsSPL14氨基酸序列信息Table 11 BambooOsSPL14amino acid sequence information
OsTB1基因的氨基酸序列比對結果顯示相似度為65.71%。竹子的氨基酸序列信息如表12。OsTB1基因的進化樹及motif見電子附圖8。motif 1、8對應的是TCP結構域。谷子和竹類的基因序列中包含motif 3而水稻和二穗短柄草序列中不存在。毛竹的兩條序列中均缺失motif 2,Raddia guianensis中還有motif 1、5、6缺失。OsTB1調節側芽的生長,編碼一個由388個氨基酸組成的蛋白,包括TCP和R結構域[19]。與其他7種植物相比毛竹的同源序列中缺少的為R結構域。

表 12 竹子OsTB1氨基酸序列信息Table 12 BambooOsTB1amino acid sequence information
HTD1基因的進化樹及motif見電子附圖9。瓜多竹序列中只有motif 7、10,Raddia guianensis序列缺失motif5、9,毛竹PedHTD1a序列缺失motif 9,PedHTD1b序 列 缺失motif 5、8、9。其余序列較為保守。HTD1基因是多分蘗矮稈基因,編碼胡蘿卜素裂解雙加氧酶CCD7,通過從休眠期釋放腋芽而不是啟動更多的腋芽來增加分蘗數[20]。
OsAAP3基因的進化樹及motif見電子附圖10。此基因在8種植物中較為保守,差異在毛竹兩條序列中缺失motif 5,蕓香竹序列中也缺失motif 5。OsAAP3基因調控有效分蘗數的增加[21]。
OsIAGLU基因的進化樹及motif見電子附圖11。motif 1、2、3、5、9、10在8個物種中均保守,瓜多竹缺失motif 4、6、7,Raddia guianensis缺失motif 6、7、8,蕓香竹缺失motif 4。OsIAGLU基因過表達,分蘗數和葉片角明顯增加,株高和穗長同時降低[22]。
THIS1基因的進化樹及motif見電子附圖12。motif 1、2、3、6在8個物種中均保守,瓜多竹、毛竹、草本竹Raddia guianensis中缺失motif 4、8,而這3種竹中都存在motif 7。此基因編碼一個第Ⅲ類的脂肪酶,調控株高變矮、小穗不育以及多分蘗產生[23]。
3 結論與討論
以上基因中,MOC1是單分蘗基因,DLT、Gn1a、LAX1、OsTB1、OsPIN5b、OsSPL14、Os-SUT2、OsTB1、RFL基因抑制分蘗產生,D10、HTD1、OsAAP3、OsCYP19?4、OsEATB、OsIAGLU、THIS1基因調控分蘗數目增加。從以上的基因進化樹可以看到,無論是使分蘗增加還是減少的基因中,2種草本竹基本都與水稻、谷子、二穗短柄草聚在一起,與3種木本竹類區分開。將草本竹Olyra latifolia和Raddia guianensis地下莖類型也作為粗型(即合軸型),竹類植物中地下莖類型為粗型的竹種與細型的竹種(即毛竹)在DLT、Gn1a、MOC1、OsPIN1、OsPIN5b、OsSPL14、OsSUT2、D10、OsAAP3基因中結構差異很大。調控分蘗增加的基因有結構域的缺失,可能導致不分蘗;而抑制分蘗生長的基因有結構域的缺失,其突變體分蘗減少,這兩種情況都有可能抑制毛竹的合軸生長,從而使其進行地下莖的延伸,形成真鞭,分析結果提示,粗型地下莖與細型地下莖的形成可能與調控分蘗數目的基因相關。
竹類地下莖的生長與內源激素也有著密切關系[24],分蘗數的變化(增加或減少)與各個激素的相互作用也存在聯系,如獨腳金內酯、生長素等植物激素的信號通路。PIN、AUX/LAX家族類蛋白參與調控植物生長素運輸,AUX/LAX調控生長素輸入,PIN調控生長素輸出[24-25],參與這些通路的基因包含上述的OsPIN1、OsPIN5b、LAX1;另外,游離IAA水平的降低可能促進水稻分蘗形成[21]。可見植物激素的相互作用對分蘗的產生起著重要的作用,由此推測,除分蘗基因外,竹類不同地下莖類型的形成與這些植物激素也有關聯之處。以上這些假設尚需后續研究來證實。
