
中藥植物羌活種子休眠解除的代謝組分析
2021-05-20 09:36:54李愛花蔣順媛郭娜黃璐琦
李愛花, 蔣順媛, 郭娜, 黃璐琦
(1.北京市植物園, 北京市花卉園藝工程技術(shù)研究中心, 北京 100093; 2.中國(guó)中醫(yī)科學(xué)院中藥資源中心, 國(guó)家道地藥材重點(diǎn)實(shí)驗(yàn)室培育基地, 北京 100700; 3.四川省中藥科學(xué)研究院, 四川省中藥質(zhì)量和創(chuàng)新研究重點(diǎn)實(shí)驗(yàn)室, 成都 610041; 4.中國(guó)中醫(yī)科學(xué)院實(shí)驗(yàn)研究中心, 北京 100700)
羌活(NotopterygiumincisumTing ex H.T. Chang)傘形科(Umbelliferae)羌活屬(Notopterygium)植物,為我國(guó)特產(chǎn),多年生草本,生長(zhǎng)于海拔2 000—4 000 m的林緣及灌叢內(nèi)。羌活為我國(guó)傳統(tǒng)大宗藥材,以根及根莖入藥,主治外感風(fēng)寒、頭痛無汗、寒濕痹、上肢風(fēng)濕疼痛。長(zhǎng)期以來,羌活藥材資源主要依靠采挖野生資源為主,導(dǎo)致羌活資源已經(jīng)處于近危狀態(tài)[1]。目前,羌活已被甘肅省、西藏自治區(qū)、寧夏自治區(qū)列為省級(jí)重點(diǎn)保護(hù)藥用植物[2-4]。為了保證其在中醫(yī)藥領(lǐng)域的應(yīng)用,亟需對(duì)其進(jìn)行人工馴化栽培。
由于羌活種子存在形態(tài)生理休眠,自然發(fā)芽率低,休眠期長(zhǎng),限制了羌活的人工馴化和栽培種植,導(dǎo)致其人工種植發(fā)展緩慢。羌活種子成熟脫落時(shí),胚未發(fā)育完全,需要待胚發(fā)育完全后,再進(jìn)行低溫處理才能解除休眠,最終萌發(fā)[5]。楊旻[6]研究發(fā)現(xiàn),低溫層積能夠明顯解除羌活的種子休眠。
近年來,一些學(xué)者利用代謝組學(xué)解析作物種子發(fā)育、萌發(fā)過程的代謝物變化[7-11],但對(duì)于種子休眠解除過程的代謝組學(xué)分析少見,且對(duì)于羌活種子休眠解除的代謝機(jī)理研究還未見報(bào)道。本研究通過對(duì)不同休眠解除階段的羌活種子進(jìn)行取樣,利用UPLC(ultra performance liquid chromatography)對(duì)其進(jìn)行代謝物質(zhì)變化分析,以解析羌活種子休眠解除過程的物質(zhì)代謝變化規(guī)律,并試圖找出種子休眠解除的關(guān)鍵化學(xué)物質(zhì),以期為羌活的人工馴化栽培提供理論背景,為具相似休眠特征植物種子的栽培種植提供重要理論參考。
1 材料與方法
1.1 試驗(yàn)材料及試劑
羌活(N.incisum)種子采自四川省甘孜藏族羌族自治州小金縣。
瓊脂為分析純,氟啶酮(fluridone,F(xiàn)L)、赤霉素(gibberellin 3,GA3)均為分析純,均購(gòu)自上海生工生物工程有限公司。
1.2 試驗(yàn)設(shè)計(jì)
1.2.1預(yù)處理成熟干種子1∶1拌細(xì)河沙,在15/5 ℃條件下,進(jìn)行暖-冷層積預(yù)處理,以完成羌活種子胚的生長(zhǎng),即解除其形態(tài)休眠(morphological dormancy,MD)。預(yù)處理過程中,每隔15 d取樣20粒,直至150 d結(jié)束。取樣種子在奧林巴斯解剖鏡(SZX7,日本奧林巴斯)下進(jìn)行解剖、形態(tài)觀察及拍照;并進(jìn)行胚長(zhǎng)和種子長(zhǎng)的測(cè)量。同時(shí)將部分種子速凍后保存在-80 ℃冰箱。選取預(yù)處理0(0MD)、15(1MD)、60(3MD)、90(5MD)、120(7MD) d進(jìn)行后續(xù)檢測(cè)。
胚率=胚長(zhǎng)/種子長(zhǎng)×100%
(1)
1.2.2生理休眠(physiologicaldormancy,PD)解除處理將經(jīng)過預(yù)處理的種子進(jìn)行以下4種處理:分別用蒸餾水(對(duì)照,CK)、100 mg·L-1氟啶酮(FL)、400 mg·L-1赤霉素(GA)浸泡24 h后,流水沖洗,播種到1%瓊脂培養(yǎng)基。另將部分種子繼續(xù)進(jìn)行5 ℃混沙低溫處理(cold stratification, CS)3個(gè)月,分別在預(yù)處理+低溫層積(CS)150 d(0PD)、150+30 d(3PD)、150+ 60 d(5PD)、150+110 d(10PD)進(jìn)行取樣。低溫層積結(jié)束的種子播種到1%瓊脂培養(yǎng)基觀察種子萌發(fā)情況,萌發(fā)試驗(yàn)每個(gè)處理3個(gè)重復(fù),每個(gè)重復(fù)100粒種子。萌發(fā)條件如下:15 ℃,光照12 h/黑暗12 h,光源為冷光源,1 000 lx。 每隔一周觀察一次種子萌發(fā)情況。同時(shí),將休眠解除處理0PD(CK)、FL、GA和CS 4個(gè)處理的種子取樣,速凍保存在-80 ℃冰箱,進(jìn)行后續(xù)代謝物檢測(cè)分析。
1.3 代謝物檢測(cè)
利用UPLC-MS/MS平臺(tái)檢測(cè)羌活種子內(nèi)代謝物。提取檢測(cè)方法參考Evans等[12],并稍作修改,簡(jiǎn)要描述如下:將儲(chǔ)藏在-80 ℃的樣品研磨成粉。稱取0.1 g研磨樣品,加入到1 mL甲醇,超聲提取30 min。離心取上清液,反復(fù)提取3次,合并提取液。氮?dú)獯蹈桑婵崭稍铩?0 μL甲醇超聲溶解,上機(jī)檢測(cè)。
色譜條件:Waters Acquity UPLC-I-Class system (Waters Corporation, Milford, MA),Acquity BEH C18色譜柱,柱溫40 ℃,流速350 μL·min-1。流動(dòng)相:0.1%甲酸水為水相,甲醇為有機(jī)相。線性洗脫梯度:(0—0.5) min,100%水;(0.5—6.0) min,60%甲醇;(6.0—6.5) min, 98%甲醇;(6.5—8.0) min,100%甲醇;進(jìn)樣量1 μL。
質(zhì)譜條件:電噴霧離子源(ESI),采取ESI (+)離子模式,離子源溫度分別為100 ℃和450 ℃,毛細(xì)管電壓分別為0.5 kV和2 kV,錐孔電壓為40 V,低能量碰撞電壓6 eV,高能量碰撞電壓分別為6 590 eV和40~70 eV。掃描范圍 質(zhì)荷比50~1 000 m·z-1。
1.4 數(shù)據(jù)處理與分析
用Origin 8.0的Boltzmann模型進(jìn)行羌活種子胚的生長(zhǎng)曲線擬合。代謝物檢測(cè)數(shù)據(jù)用Progenesis QI軟件進(jìn)行分析。原始數(shù)據(jù)進(jìn)行Z-score歸一化處理后,用SPSS 11.0進(jìn)行PCA和聚類分析。
2 結(jié)果與分析
2.1 羌活種子預(yù)處理過程的內(nèi)部結(jié)構(gòu)變化
圖1顯示,羌活種子成熟散布時(shí),種子內(nèi)部大部分為胚乳,僅在果疤一端有一團(tuán)未分化的透明物質(zhì)為原胚, 0.1~0.2 mm,原胚的頂端有1個(gè)白色胚結(jié)構(gòu)。在形態(tài)休眠解除的處理過程中,胚不斷長(zhǎng)大,至150 d時(shí),胚長(zhǎng)至約為種子長(zhǎng)度的三分之二,胚乳軟化。在胚發(fā)育過程中,隨著處理時(shí)間的推移,胚率的變化表現(xiàn)為S曲線,處理80 d左右時(shí),胚率的變化速度最快,即羌活種子的胚生長(zhǎng)最快,這是加速胚發(fā)育速度的關(guān)鍵時(shí)間點(diǎn)。
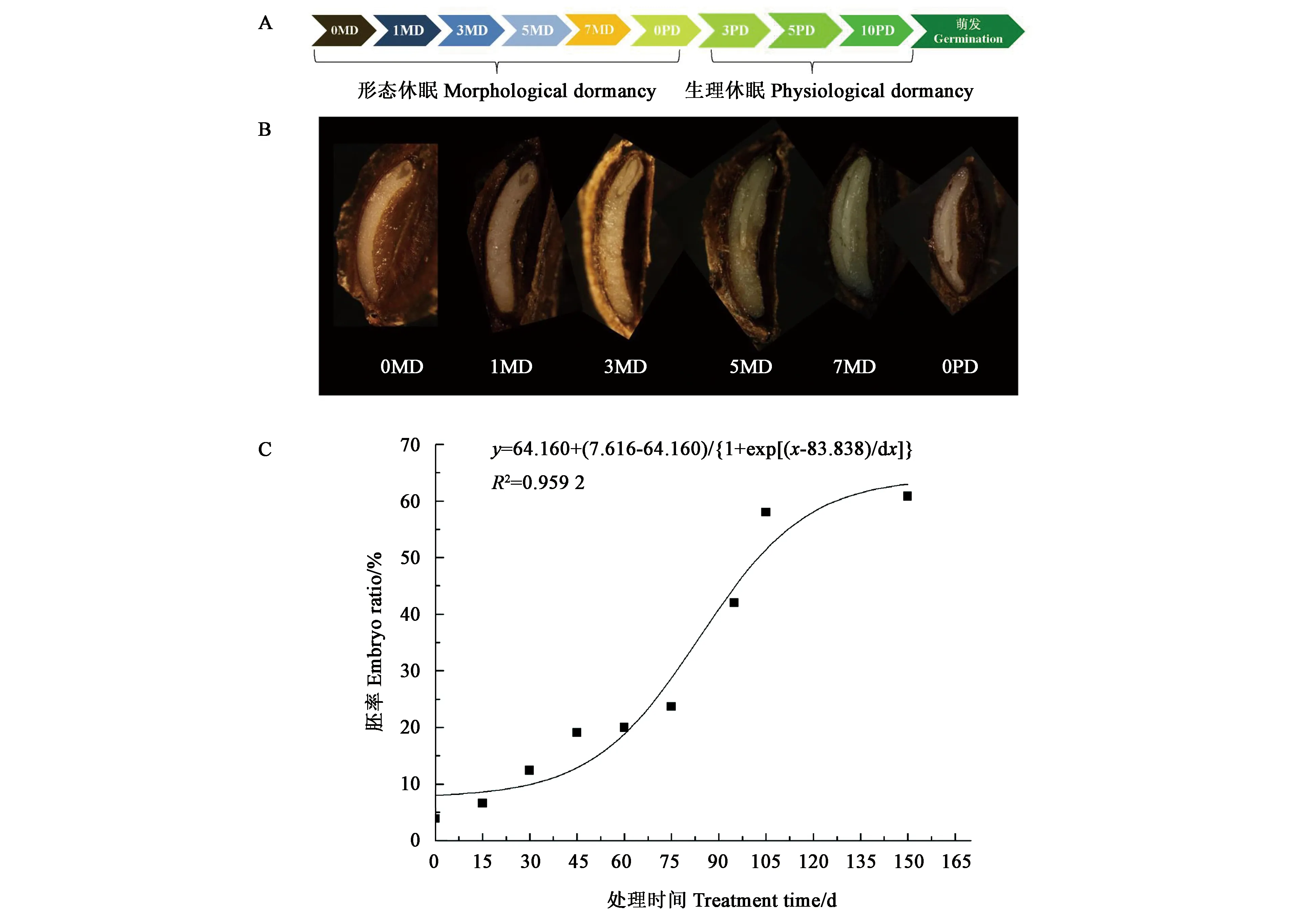
A:種子休眠解除處理過程;B:關(guān)鍵時(shí)間點(diǎn)的胚形態(tài);C:胚生長(zhǎng)曲線
2.2 羌活種子預(yù)處理過程的代謝組分析
對(duì)羌活種子的整個(gè)預(yù)處理及低溫層積過程取樣,進(jìn)行代謝物檢測(cè)。對(duì)正離子模式原始檢測(cè)結(jié)果歸一化處理后,進(jìn)行H聚類分析,結(jié)果(圖2)發(fā)現(xiàn),0MD、1MD、3MD聚類到一組,5MD、7MD、0PD、3PD聚類到一組,而5PD、10PD聚類到一組。可見,總體來看,在羌活種子的整個(gè)處理過程中,種子內(nèi)化合物的代謝模式,分為3個(gè)階段:0—60 d、90—(150+30) d和(150+60)—(150+110)d。其中第3個(gè)階段的兩個(gè)處理5PD和10PD的羌活種子已具備萌發(fā)能力,5PD的種子通過ABA抑制劑處理已可以萌發(fā),而10PD的種子可以直接萌發(fā)[13]。而0PD、3PD甚至之前處理階段的羌活種子不萌發(fā),且經(jīng)過氟啶酮處理后也基本不萌發(fā)。

注:色度條表示代謝物的豐度,以相對(duì)于所有處理平均值的變化來表示。
對(duì)檢測(cè)結(jié)果進(jìn)行PCA分析,結(jié)果見圖3。可見,羌活種子處理整個(gè)過程的不同階段可以有很好的區(qū)分度,隨著時(shí)間的推移,不同處理的代謝物表現(xiàn)出順時(shí)針的旋轉(zhuǎn)模式,0MD、1MD、3MD、5MD、7MD沿著t[1] 主成分軸由小到大排列,0PD、3PD、5PD、10PD按照t[2] 主成分軸由大到小排列。即t[1] 主成分所代表的化合物能夠很好地將0MD、1MD、3MD、5MD、7MD區(qū)分開,而t[2]主成分所代表的化合物對(duì)0PD、3PD、5PD、10PD能夠很好地分開。那么,t[1]、t[2]主成分所代表的化合物即是這些處理之間發(fā)生代謝變化的關(guān)鍵化合物。
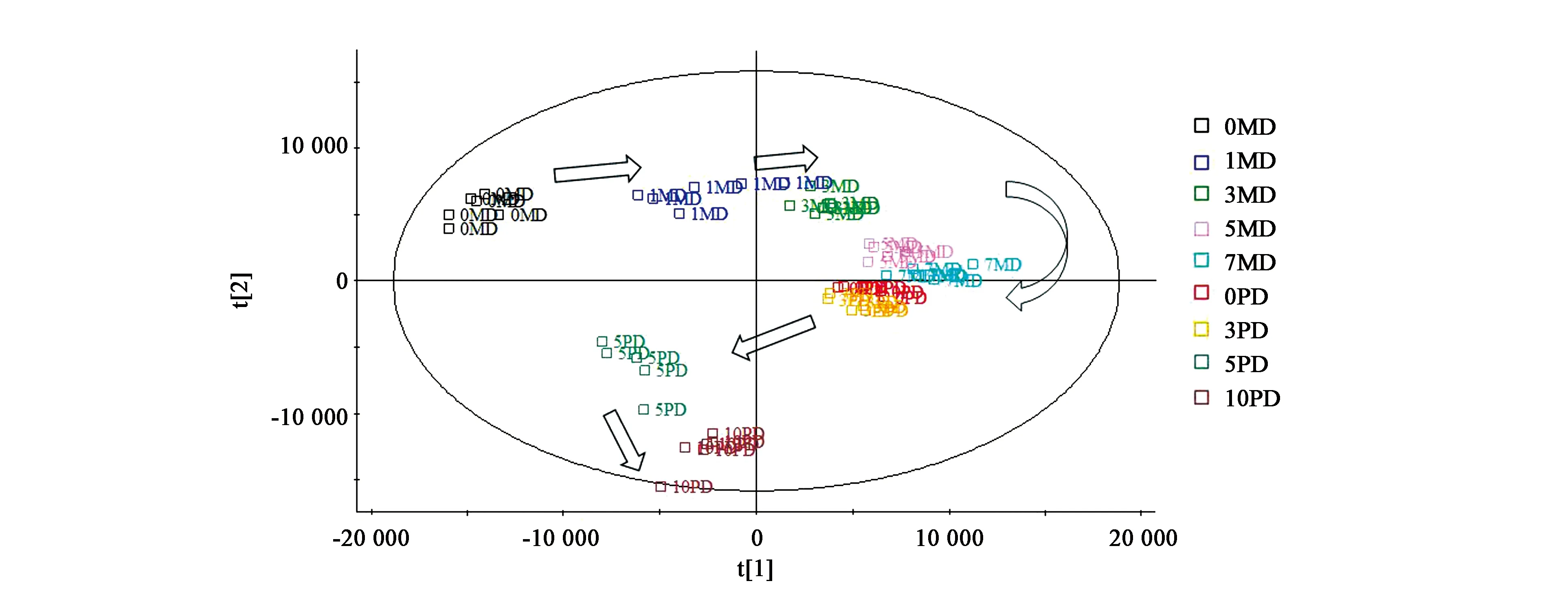
圖3 羌活種子處理過程代謝物的PCA分析
2.3 羌活種子休眠解除處理的代謝組分析
對(duì)羌活種子休眠解除處理的代謝物檢測(cè)結(jié)果進(jìn)行PCA分析,結(jié)果(圖4)顯示,對(duì)完成形態(tài)休眠羌活種子進(jìn)行的休眠解除處理(5PDFL、5PDGA和10PD)的代謝物的PCA分析結(jié)果,與5PD處理可以顯著區(qū)分。而3個(gè)解除休眠處理間,5PDFL與5PDGA和10PD可以明顯區(qū)分,但是5PDGA與10PD不能區(qū)分。t[1] 主成分所代表的化合物能夠很好地將5PD、5PDFL、5PDGA和10PD區(qū)分開,而不能將5PDGA和10PD 區(qū)分。萌發(fā)試驗(yàn)顯示,5PDFL和10PD的種子可以萌發(fā)[13],而5PDGA的種子萌發(fā)率很低(未發(fā)表)。

圖4 休眠解除過程羌活種子代謝物的PCA分析
2.4 羌活種子休眠解除處理的關(guān)鍵化合物
通過分析可萌發(fā)處理5PDFL和10PD的共有代謝物,及二者與不能萌發(fā)處理(5PD)的差別代謝物,結(jié)果(圖5)發(fā)現(xiàn),正離子模式的加荷粒子 411.1607:[M+Na]+和389.1416:[M+H]+,碎片離子包括307: [M+Na-CH3COOH-CO2]+、351: [M + Na-CH3COOH]+、335: [[M+Na-CH3CO-H2O]+和371: [M+H-H2O]+。表明該化合物的分子式可能為C31H42O17。負(fù)離子模式的質(zhì)譜圖,顯示加荷粒子387.1652:[M-H]-,表明該化合物的經(jīng)驗(yàn)分子式為C31H42O17。這與獨(dú)腳金內(nèi)酯(strigolactones,SLs)化合物列當(dāng)醇乙酸酯(orobanchyl acetate)的分子式一致(圖6)。 離子片斷分析表明,列當(dāng)醇乙酸酯脫下一個(gè)糖基和1個(gè)分子H2O (180 Da)。基于UPLC-MS/MS檢測(cè)的列當(dāng)醇乙酸酯的峰面積,計(jì)算5PDFL和10PD處理相對(duì)于5PD的列當(dāng)醇乙酸酯變化,發(fā)現(xiàn)FL和CS處理的列當(dāng)醇乙酸酯水平分別顯著升高12.1和 28.5倍(圖7)。
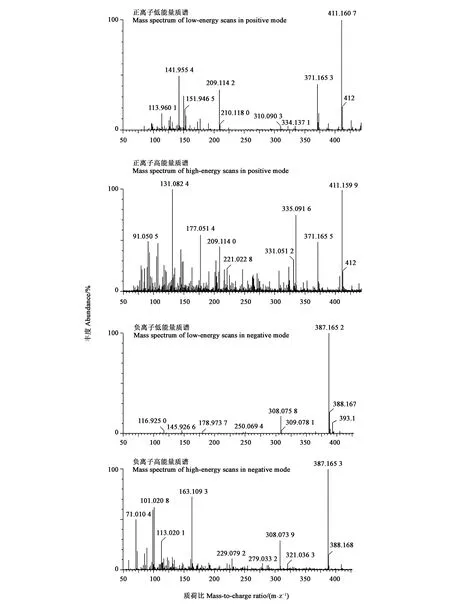
圖5 羌活種子中列當(dāng)醇乙酸酯的質(zhì)譜圖

圖6 列當(dāng)醇乙酸酯的結(jié)構(gòu)式
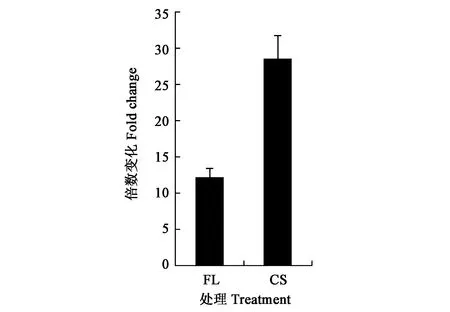
圖7 羌活種子中列當(dāng)醇乙酸酯的含量變化
3 討論
研究發(fā)現(xiàn),完成形態(tài)發(fā)育的羌活種子胚仍然不能萌發(fā),低溫層積3個(gè)月(CS)處理可以達(dá)到73.5%的萌發(fā)率;用氟啶酮溶液(FL)浸泡后,也可以達(dá)到81.3%的萌發(fā)率;不經(jīng)過處理的種子(Con)僅有12.8%萌發(fā);GA3溶液浸泡(GA)的種子萌發(fā)情況與未處理(Con)種子萌發(fā)情況類似[13]。可見,羌活種子通過預(yù)處理完成種子胚的發(fā)育,但是發(fā)育完全的胚仍然不能全部萌發(fā),需要通過低溫層積3個(gè)月解除休眠而萌發(fā),氟啶酮也可以有效解除完成胚發(fā)育種子的休眠。
本研究對(duì)羌活種子取樣進(jìn)行代謝物檢測(cè)后發(fā)現(xiàn),羌活種子整個(gè)處理過程分為3個(gè)階段:0—60 d(0MD、1MD、3MD)、90—180 d(5MD、7MD、0PD、3PD)、210—260 d(5PD、10PD),第三個(gè)階段的兩個(gè)處理5PD和10PD種子已具備萌發(fā)能力。t[1] 主成分所代表的化合物能夠很好地將形態(tài)休眠階段(即0MD、1MD、3MD、5MD、7MD)區(qū)分開,表明這些化合物是0MD、1MD、3MD、5MD、7MD胚發(fā)育處理過程發(fā)生主要變化的化合物。t[2] 主成分所代表的化合物對(duì)生理休眠階段(0PD、3PD、5PD、10PD)很好地區(qū)分,表明這些化合物主要是在生理休眠解除過程中發(fā)生變化的。而t[1]、t[2]主成分所代表的主要代謝化合物的分析鑒定,還需要進(jìn)一步研究。
獨(dú)腳金內(nèi)酯是一種來源于類胡蘿卜素的植物激素,能夠誘導(dǎo)寄生植物Orobanchesp.和Strigasp.種子萌發(fā)[14],促進(jìn)叢枝真菌分枝、抑制地上分枝的作用[15],然而其是否能夠誘導(dǎo)非寄生植物的種子萌發(fā),尚未見有報(bào)道。本研究發(fā)現(xiàn),在ABA抑制劑(FL)處理和低溫層積處理誘導(dǎo)羌活種子休眠解除過程中,列當(dāng)醇乙酸酯,一種SLs化合物,發(fā)生顯著積累。FL處理后SLs的生物合成水平升高12.1倍,顯著低于CS處理后SLs的合成水平升高倍數(shù)(28.5倍)。推測(cè)FL處理抑制八氫番茄紅素脫氫酶(PDS)活性[16],也降低了種子休眠解除過程SLs的生物合成水平。Li等[13]研究也發(fā)現(xiàn),SLs的合成酶基因NiD27、NiCCD8、NiCYP711A1和NiDAD2在5PDFL和CS處理中均顯著上調(diào)表達(dá)。因此,推測(cè)SLs的含量變化可能是羌活種子休眠解除過程是關(guān)鍵環(huán)節(jié)。
猜你喜歡
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
海峽科技與產(chǎn)業(yè)(2016年3期)2016-05-17 04:32:12
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
