龍須菜(Gracilariopsis lemaneiformis)海藻糖-6-磷酸合成酶(TPS)對逆境脅迫的響應(yīng)*
2021-05-26 08:08:36張瑩瑩宣雯燕張小倩徐年軍
海洋與湖沼 2021年3期
張瑩瑩 呂 燕 宣雯燕 張小倩 徐年軍 孫 雪
龍須菜()海藻糖-6-磷酸合成酶(TPS)對逆境脅迫的響應(yīng)*
張瑩瑩 呂 燕 宣雯燕 張小倩 徐年軍 孫 雪①
(寧波大學(xué)海洋學(xué)院 浙江省海洋生物工程重點實驗室 寧波 315211)
非還原性二糖海藻糖及其代謝物是調(diào)控植物生長發(fā)育和逆境響應(yīng)的信號分子。本研究以大型海藻龍須菜()為對象, 從基因轉(zhuǎn)錄、蛋白和酶活性3個水平探討了海藻糖合成酶——海藻糖-6-磷酸合成酶(TPS)對高溫、高鹽及滲透脅迫的響應(yīng)。龍須菜中4條TPS序列均具有TPS家族保守結(jié)構(gòu)域(Glyco-transf-20)和TPP家族保守結(jié)構(gòu)域(Trehalose-PPase), 且屬于Class I亞家族。在轉(zhuǎn)錄水平上, 高鹽脅迫主要促進(jìn)了1、2和4基因的表達(dá), 而滲透脅迫則總體抑制了1、2和3基因的表達(dá)。在高鹽脅迫48 h時, TPS1蛋白含量升高到對照組的2.03倍。在高溫和高鹽脅迫24 h時, TPS活性升高, 而在高鹽脅迫48 h及滲透脅迫條件下酶活性降低。可見海藻糖-6-磷酸合成酶參與了龍須菜抗高溫和高鹽脅迫的應(yīng)答, 但對滲透脅迫不敏感。該研究為提高龍須菜抗逆性及培育抗逆龍須菜品種提供了參考。
龍須菜; 海藻糖; 海藻糖-6-磷酸合成酶; 非生物脅迫
龍須菜()隸屬紅藻門(Rhodophyta)、江蘺目(Gracilariales), 是一種生長在潮間帶的大型經(jīng)濟(jì)海藻。龍須菜不僅可用作瓊膠提取原料、鮑的餌料及食材, 而且在去除氮磷、降低海水富營養(yǎng)化、維持海洋生態(tài)平衡方面也具有重要的環(huán)境效益(Chen, 2015)。野生型龍須菜主要分布在山東沿海, 其生長主要集中在春秋兩季, 生長的適溫范圍為12—23°C (張學(xué)成等, 2009)。隨著耐高溫981品系等的成功培育, 大規(guī)模的龍須菜栽培逐漸從原來的北方低溫海區(qū)擴(kuò)展到了南方高溫海區(qū)。但南方夏季海水溫度高, 龍須菜無法成功度夏, 并且潮汐以及采收運輸過程中藻體還可能會受到滲透脅迫的影響。因此, 研究龍須菜抗逆脅迫對其夏季栽培及運輸?shù)染哂幸欢ǖ膮⒖純r值。
海藻糖是一種非還原性二糖, 廣泛存在于植物、細(xì)菌、真菌和無脊椎動物中。海藻糖不僅可用于生物體內(nèi)碳的存儲和運輸, 還可作為壓力保護(hù)劑使生物更好地抵抗不利環(huán)境脅迫(Schluepmann, 2004; O’Hara, 2013; Tang, 2016)。高含量海藻糖對于許多沙漠植物如折扇葉()、卷柏()等的生存是必需的(Goddijn, 1999; Paul, 2008)。在熱激條件下, 釀酒酵母()中海藻糖積累量可達(dá)到細(xì)胞干重的30% (Bell, 1998)。擬南芥()中海藻糖含量的增加會提高其對鹽脅迫的耐受性(Krasensky, 2014)。可見生物體內(nèi)海藻糖的積累與逆境脅迫存在著密切關(guān)系。此外, 海藻糖及其代謝物海藻糖-6-磷酸(Trehalose-6- phosphate, T6P)還參與一些信號調(diào)控, 它們可能作為中央代謝調(diào)節(jié)器, 協(xié)調(diào)植物中多種代謝通路。
自然界中有5條海藻糖合成途徑, 其中OtsA-OtsB途徑在生物中分布最廣泛, 也是植物中發(fā)現(xiàn)的唯一的海藻糖合成途徑(Paul, 2008)。在植物中, 海藻糖合成由兩步反應(yīng)組成, 首先海藻糖-6-磷酸合成酶(Trehalose-6-phosphate synthase, TPS)催化尿苷二磷酸-葡萄糖與葡萄糖-6-磷酸合成T6P, 然后T6P在海藻糖磷酸磷酸酶(Trehalose-6-phosphate phosphatase, TPP)催化下脫磷酸形成海藻糖。海藻糖-6-磷酸合成酶在調(diào)控碳水化合物代謝、生長發(fā)育及脅迫應(yīng)答響應(yīng)中起著重要作用。如即使在合適的開花條件下, 缺失1的擬南芥開花也會推遲到很晚(Wahl, 2013)。與非轉(zhuǎn)基因水稻()相比, 過表達(dá)基因的轉(zhuǎn)基因水稻在高鹽、干旱和低溫條件下能夠更好地維持生長和抗光氧化損傷(Li, 2011)。在高溫下,1基因敲除的酵母菌不生長, 而1過表達(dá)的菌株則生長良好(Jiang, 2016)。
目前許多生物中基因序列信息已經(jīng)闡明。盡管高等植物中不會大量積累海藻糖, 但是基因組測序結(jié)果顯示植物中存在著基因家族。如水稻、小麥()和大豆()中分別有11條、12條和13條基因, 木豆()中則有28條序列(Zang, 2011; Singh, 2012; Xie, 2015)。但是在藻類中關(guān)于基因信息及抗逆性的工作較少。在本研究中, 我們分析了從龍須菜基因組中篩選到的4條基因序列, 并在轉(zhuǎn)錄、蛋白及酶活性3個水平探究了TPS對高溫、高鹽及滲透脅迫的響應(yīng)。該研究豐富了海藻糖在藻類抗逆脅迫中的工作, 為提高龍須菜抗逆性提供了資料。
1 材料與方法
1.1 材料與培養(yǎng)條件
龍須菜耐高溫981品系采自福建霞浦縣長春鎮(zhèn)養(yǎng)殖場(119°59′E, 26°43′N)。首先用毛刷清洗去除藻體表面附著物, 然后挑取生長狀態(tài)良好的龍須菜, 用鑷子截取5—10 cm長的藻體尖部, 置于光照培養(yǎng)箱中進(jìn)行培養(yǎng)。培養(yǎng)條件為: 溫度23 °C, 光強約40 μmol/(m2×s), 光周期12L : 12D, 培養(yǎng)基使用鹽度30的無菌海水。
1.2 材料處理
實驗共設(shè)置4個組: 對照組(CK: 23 °C, 鹽度30)、高溫組(HS: 30 °C, 鹽度30)、高鹽組(SS: 23 °C, 鹽度45)和滲透脅迫組(OS: 23 °C, 鹽度30, 5% PEG6000), 其他培養(yǎng)條件與1.1相同, 每組3個重復(fù)。分別在培養(yǎng)0、3、6、12 h取約100 mg樣品用于實時熒光定量PCR(RT-qPCR)分析; 分別在0、12、24、48 h取約200 mg樣品用于蛋白免疫印跡(Western blot)分析或酶活性測定。
1.3 TPS序列的生物信息學(xué)分析
結(jié)合龍須菜基因組測序和cDNA末端快速擴(kuò)增(RACE)技術(shù), 我們得到4條龍須菜TPS基因序列, 分別記為1—4。利用ProtParam tool (https: //web. expasy.org/protparam/)進(jìn)行TPS蛋白基本理化性質(zhì)預(yù)測。利用SMART (http: //smart.embl-heidelberg.de/)分析TPS蛋白結(jié)構(gòu)和功能域。使用TMHMM (http://www. cbs.dtu.dk/services/TMHMM/)對TPS蛋白中跨膜結(jié)構(gòu)進(jìn)行預(yù)測。分別利用SignalP (http://www.cbs.dtu.dk/ services/SignalP/)和Plant-mPLoc (http://www.csbio. sjtu.edu.cn/bioinf/plant-multi/)對TPS蛋白進(jìn)行信號肽和亞細(xì)胞定位分析。用MEGA 7.0軟件的鄰接法構(gòu)建TPS系統(tǒng)進(jìn)化樹(饒莉萍等, 2020), bootstrap值設(shè)置為1000。
1.4 TPS基因轉(zhuǎn)錄分析
參考林麗春等(2019)的方法, 將1.2中的樣品進(jìn)行總RNA提取和cDNA的合成。以n和為內(nèi)參基因, 利用Primer Premier 5.0軟件設(shè)計和內(nèi)參基因序列的RT-qPCR引物(表1), 引物序列交由華大基因有限公司合成。所有基因片段均經(jīng)過T-A克隆和測序驗證為正確的序列。在Eppendorf熒光定量PCR儀(Mastercycler ep realplex, Germany)中進(jìn)行RT-qPCR擴(kuò)增, 所用試劑和反應(yīng)程序等參照孫鵬等(2019)的方法進(jìn)行。采用2?ΔΔCT法分析目的基因的相對表達(dá)水平(Livak, 2001)。
1.5 抗體制備與Western blot分析
根據(jù)抗體制備原則及TPS氨基酸序列設(shè)計多肽序列, 交由杭州華安生物技術(shù)有限公司完成抗體制備。經(jīng)過特異性驗證和檢測, 只有TPS1蛋白制得了合格抗體, 其多肽序列為NDDPNRIRTTPRSIHC。
根據(jù)呂燕等(2019)的方法進(jìn)行蛋白提取和濃度測定。總蛋白經(jīng)過SDS-PAGE電泳后, 將膠轉(zhuǎn)移到聚偏二氟乙烯(PVDF)膜上, 用5%牛血清白蛋白于室溫封閉2 h, 然后用封閉液稀釋(1:200)的一抗溶液于4 °C孵育過夜, 洗膜3次, 加入1:8000稀釋的HRP標(biāo)記的羊抗兔IgG二抗(康為世紀(jì), 貨號CW0103)溶液孵育1.5 h, 洗膜, 最后加入Western Bright ECL化學(xué)發(fā)光液(Advansta)反應(yīng)后, 在Tanon 5200全自動化學(xué)發(fā)光圖像分析系統(tǒng)中檢測、成像, 用Image J軟件分析目的蛋白條帶的灰度值。
表1 RT-qPCR的引物

Tab.1 Primers used for RT-qPCR
1.6 酶的提取及TPS活性分析
酶提取方法參考Pramanik等(2005)并進(jìn)行了一些修改。將200 mg龍須菜在液氮中研磨成粉末, 然后加入50 mmol/L的Tris-HCl緩沖液(含100 mmol/L的NaCl, DTT和PMSF, pH 7.5)提取。最后將提取物以13000×(4 °C)離心5 min, 上清液即為酶粗提液。
TPS活性測定方法參考Hottiger等(1987)。該方法檢測了在葡萄糖-6-磷酸存在下UDP-葡萄糖中UDP的釋放, 以340 nm吸光度的降低來計算UDP的釋放量。將每g樣品每min形成1 nmol UDP定義為一個酶活性單位(U), 酶活性表示為U/g藻體鮮重(FW)。
1.7 數(shù)據(jù)分析
采用Excel 2013進(jìn)行數(shù)據(jù)處理和作圖, 實驗數(shù)據(jù)均以平均值±標(biāo)準(zhǔn)偏差(mean±SD)表示。用SPSS 13.0中的One-way ANOVA和Tukey多重比較進(jìn)行統(tǒng)計學(xué)分析, 以<0.05設(shè)為差異顯著。對于RT-qPCR的數(shù)據(jù), 把每個組在0 h時的基因表達(dá)量記為1, 將基因表達(dá)倍數(shù)上調(diào)2倍或下調(diào)0.5倍設(shè)為差異顯著(Chang, 2014)。
2 結(jié)果與分析
2.1 TPS序列和結(jié)構(gòu)特征
龍須菜4條基因編碼的蛋白特征如表2所示, 其在GenBank中的序列號分別為MT919220—MT919223。其中3基因序列最長, 其次是1, 兩者分別編碼1140和1007個氨基酸, 而2和4開放閱讀框較短, 其編碼蛋白分子量分別為84.9 ku和100.6 ku。4條序列中, TPS1、TPS2和TPS4為酸性蛋白(pI為5.60—6.14), 而TPS3則偏弱堿性(pI為7.94)。4條TPS蛋白的不穩(wěn)定指數(shù)在43.75—52.98之間, 以≤40為穩(wěn)定性蛋白的標(biāo)準(zhǔn), 龍須菜TPS蛋白均為不穩(wěn)定性蛋白; 平均親水系數(shù)在–0.382和–0.229之間, 均為親水性蛋白。
對龍須菜TPS蛋白序列進(jìn)行結(jié)構(gòu)域分析(圖1), 發(fā)現(xiàn)4條TPS的N端都有TPS家族保守結(jié)構(gòu)域(Glyco-transf-20), C端都包含TPP家族保守結(jié)構(gòu)域(Trehalose-PPase)。除了以上兩個結(jié)構(gòu)域之外, TPS1蛋白的N端還包含一個CBM-2保守結(jié)構(gòu)域(carbohydrate-binding module), 而TPS3蛋白的C端764—882 aa和933—1121 aa各有1個TPP家族保守結(jié)構(gòu)域。跨膜區(qū)和信號肽分析發(fā)現(xiàn)TPS1—TPS4蛋白均無跨膜區(qū), 也無信號肽序列。亞細(xì)胞定位分析結(jié)果表明TPS1、TPS2可能位于細(xì)胞質(zhì)或液泡中, TPS3、TPS4則可能位于葉綠體或液泡中。
表2 龍須菜TPS蛋白序列的基本特征
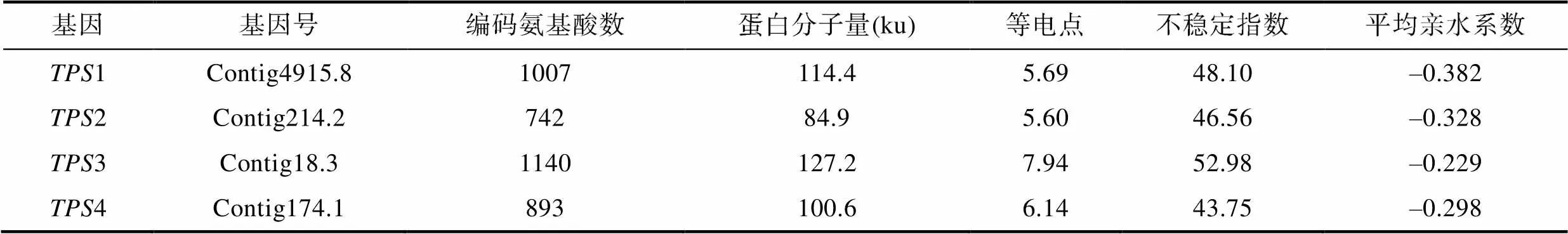
Tab.2 Characteristics of TPSsequences in G. lemaneiformis
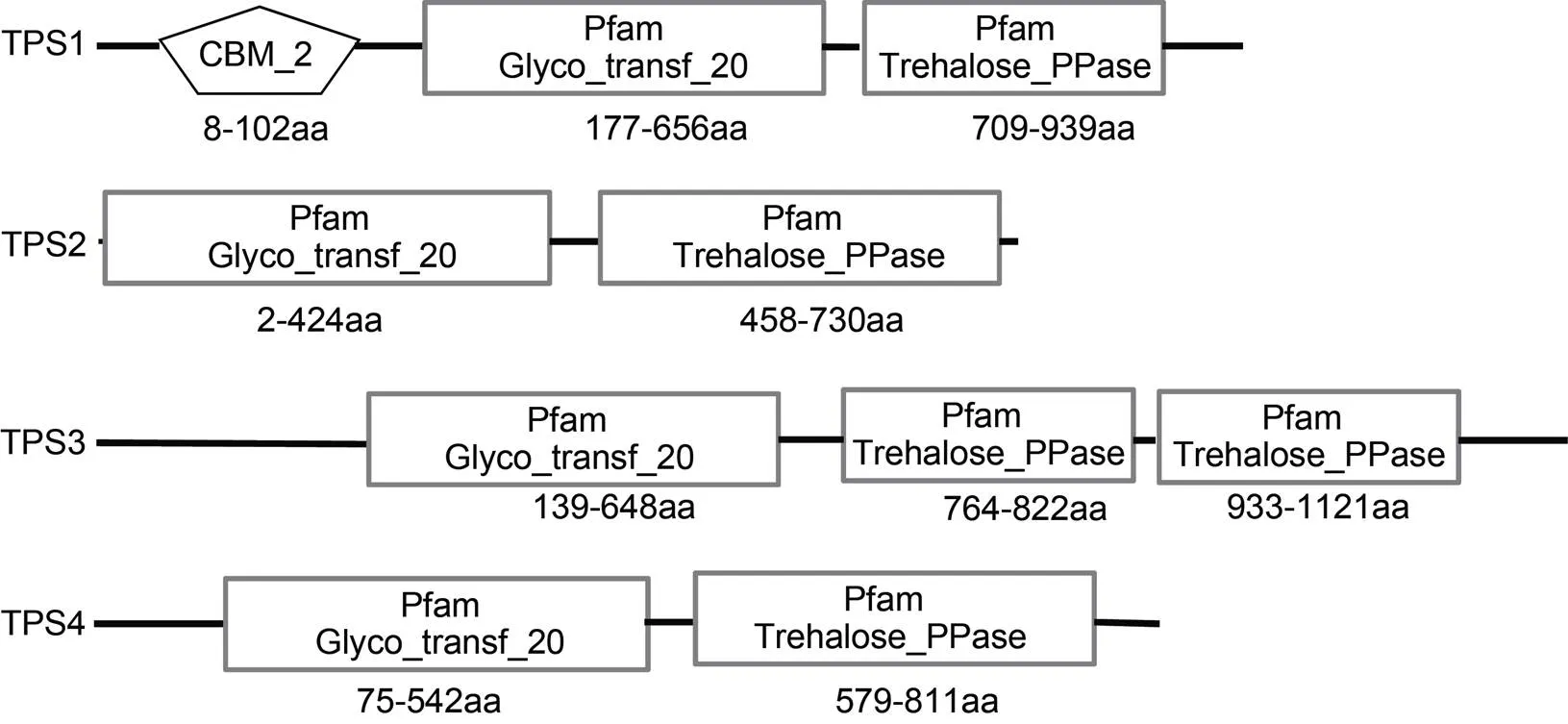
圖1 龍須菜TPS蛋白的結(jié)構(gòu)域組成
2.2 龍須菜TPS蛋白序列系統(tǒng)進(jìn)化分析
在GenBank數(shù)據(jù)庫中選了24條藻類TPS蛋白序列, 將這些TPS與4條龍須菜TPS序列一起構(gòu)建系統(tǒng)進(jìn)化樹(圖2a)。在進(jìn)化樹中, 28條TPS分列在4個簇中, 各個簇中分別包含1條龍須菜TPS序列。在每一個簇中, 龍須菜TPS與繩狀龍須菜()TPS距離最近, 相似性均在99%以上, 與皺波角叉菜()TPS相似性為51%—77%, 3者的4條TPS分別一一對應(yīng)。其次較為相似的為壇紫菜()、溫泉紅藻()等紅藻。
為了進(jìn)一步分析龍須菜TPS所屬亞家族, 我們又將龍須菜中4條TPS與釀酒酵母()TPS序列一起構(gòu)建了進(jìn)化樹(圖2b)。結(jié)果顯示龍須菜的4條TPS與TPS1聚在一個簇中,TPS2、TPS3和TSL1分布在外圍, 說明龍須菜中4條TPS蛋白均與TPS1的關(guān)系較近。再結(jié)合Leyman等(2001)對擬南芥中11條TPS的分析結(jié)果, 即TPS1—TPS4與TPS1相似度最大, 屬于Class I亞家族, 而TPS5-TPS11與TPS2相似度最大, 屬于Class II亞家族, 我們推測龍須菜中4條TPS序列屬于Class I亞家族。
2.3 龍須菜TPS基因?qū)?種脅迫條件的響應(yīng)
龍須菜中4條基因?qū)Σ煌{迫條件的響應(yīng)不同(圖3)。高鹽脅迫顯著促進(jìn)了1、2和4的轉(zhuǎn)錄表達(dá), 而滲透脅迫則多抑制1、2和3的表達(dá)。各基因的表達(dá)如下:
1基因的表達(dá)模式: 在對照組中,1基因的轉(zhuǎn)錄水平基本穩(wěn)定(>0.05)。在高溫脅迫組中,1表達(dá)量呈先上升后下降的趨勢, 在6 h和12 h 時1基因表達(dá)量分別為0 h的1.90倍(>0.05)和0.48倍(<0.05), 但與對照組中相應(yīng)時間點之間差異不顯著。在高鹽脅迫組中,1表達(dá)量在3 h和6 h時分別是0 h的2.39倍和2.74倍(<0.05), 且與對照組中差異顯著。在滲透脅迫組中,1基因表達(dá)變化幅度較小, 并且在6 h時低于對照組。
2基因的表達(dá)模式: 在對照組中,2基因的表達(dá)量隨時間的增加而逐漸上升。在高溫條件下,2基因表達(dá)量在6 h時最高, 但與對照組無顯著差異。高鹽促進(jìn)了2基因的表達(dá), 在3、6和12 h時其表達(dá)量分別為0 h的7.45、7.19和11.21倍, 但僅在3 h時高于對照組(<0.05)。與1類似, 滲透脅迫條件對2基因表達(dá)量影響較小, 僅在12 h時低于對照組(<0.05)。
3基因的表達(dá)模式: 對照組中3基因表達(dá)變化穩(wěn)定。高溫處理后3基因表達(dá)量在6 h時達(dá)到最高, 但與對照組差異不顯著。除滲透脅迫12 h時3表達(dá)量稍有下降之外, 在其余時間點高鹽和滲透脅迫處理對3基因表達(dá)量均無顯著影響。
4基因的表達(dá)模式: 在高溫及滲透脅迫條件下, 與前面3條基因表達(dá)變化不同,4基因在3 h后表達(dá)量均下調(diào)。且與對照組各時間點無顯著差異。而高鹽脅迫時,4基因在3 h和6 h時表達(dá)量高于對照組。
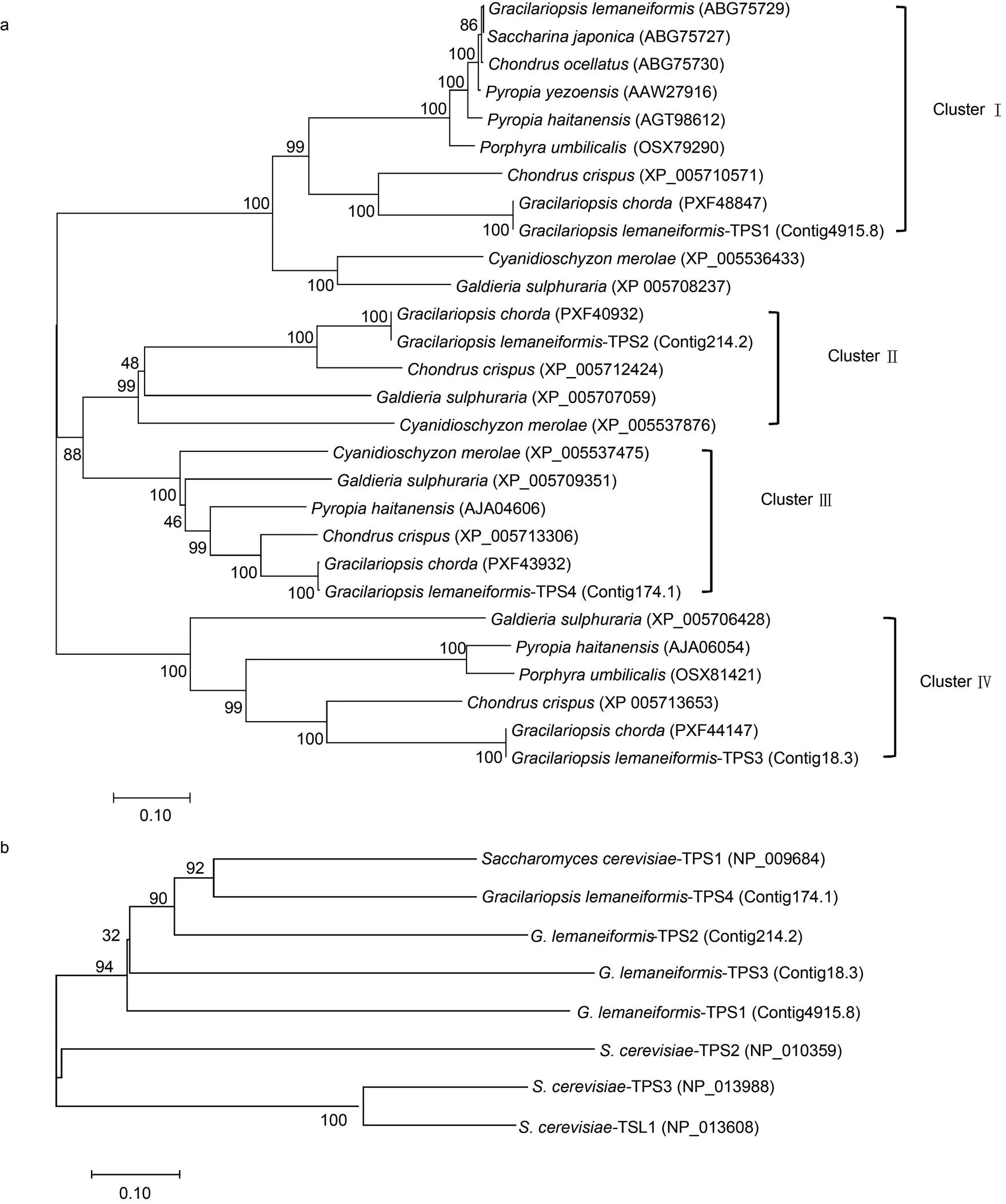
圖2 基于TPS蛋白序列構(gòu)建的系統(tǒng)進(jìn)化樹
注: a. 藻類TPS; b. 龍須菜和釀酒酵母TPS
2.4 龍須菜TPS1蛋白對3種脅迫條件的響應(yīng)
龍須菜TPS1蛋白水平變化與1轉(zhuǎn)錄變化類似(圖4)。在對照組、高溫和滲透脅迫組中, TPS1蛋白表達(dá)量在48 h內(nèi)比較穩(wěn)定, 與對照組之間也無顯著差異。而在高鹽脅迫組中, TPS1蛋白表達(dá)水平緩慢上升, 在48 h時其表達(dá)量達(dá)到對照組的2.03倍(<0.05)。可見高鹽處理可促進(jìn)龍須菜TPS1轉(zhuǎn)錄和蛋白水平的升高。
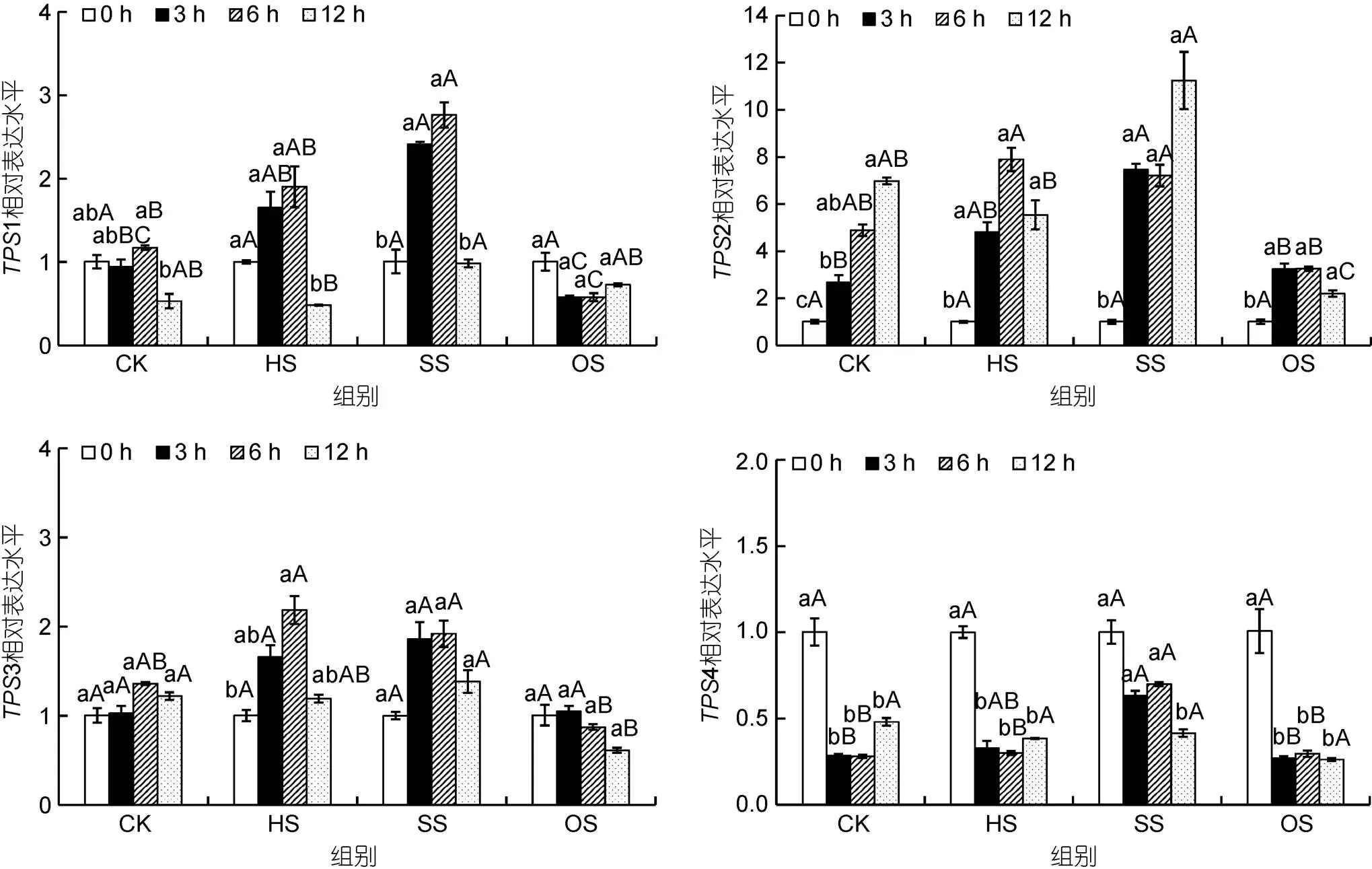
圖3 不同脅迫條件對龍須菜TPS基因表達(dá)的影響
注: CK. 對照組, HS. 高溫組, SS. 高鹽組, OS. 滲透脅迫組; 不同小寫字母表示同組中不同時間點差異顯著(<0.05), 不同大寫字母表示同一時間點不同組差異顯著(<0.05)。下同
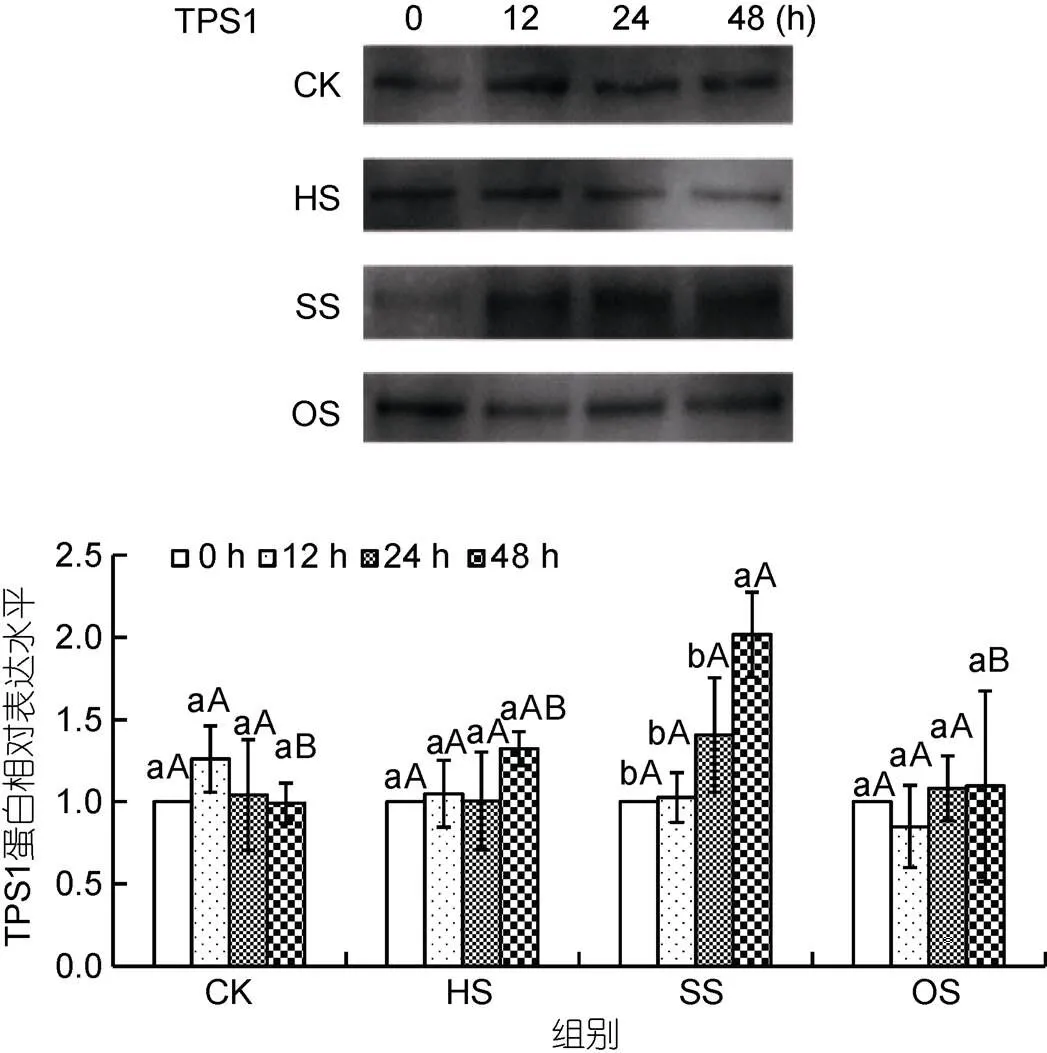
圖4 不同脅迫條件對龍須菜TPS1蛋白表達(dá)的影響
2.5 龍須菜TPS酶對3種脅迫條件的響應(yīng)
3種脅迫處理顯著影響了龍須菜TPS活性(圖5)。在對照組中, 龍須菜TPS活性比較穩(wěn)定。在高溫組中, TPS活性在24 h達(dá)到最高(1.28倍,<0.05), 并且高于對照組。在高鹽組中, 龍須菜TPS活性在24 h時升高為對照組(24 h)的1.11倍(<0.05), 在48 h時其活性降低至對照組(48 h)的66.34% (<0.05)。在滲透脅迫組中, TPS活性逐漸下降, 且在12—48 h與對照組差異顯著。可見, 高溫脅迫可以短時促進(jìn)TPS活性, 高鹽脅迫則先促進(jìn)、后抑制TPS活性, 而滲透脅迫則會顯著抑制TPS活性。
3 討論
海藻糖具有高度的化學(xué)穩(wěn)定性, 是天然雙糖中最穩(wěn)定的, 素有“生命之糖”之稱。海藻糖對生物膜、蛋白質(zhì)和核酸等生物大分子具有非特異的保護(hù)作用(Richards, 2002), 其代謝及信號途徑在植物生長和抗逆等方面發(fā)揮了重要作用。
目前越來越多的全基因組測序工作已完成, 人們在已測序高等植物基因組中發(fā)現(xiàn)了龐大的基因家族(Yang, 2012)。根據(jù)與酵母1或2的相似性,基因可以被歸為Class I和Class Ⅱ兩個亞家族(Leyman, 2001)。近年來, 藻類基因序列也相繼被報道。Wang等(2010)克隆了包括海帶()、條斑紫菜()、壇紫菜等在內(nèi)的10種大型海藻的基因序列, 并且發(fā)現(xiàn)它們在核苷酸和氨基酸組成上具有高度保守性。大型紅藻繩狀龍須菜和皺波角叉菜基因組中各有4條基因(Collén, 2013; Lee, 2018), 本課題組在龍須菜基因組中也發(fā)現(xiàn)了4條基因(Sun, 2018), 且4條基因均屬于Class I亞家族。TPS氨基酸序列高度保守, 如龍須菜與繩狀龍須菜TPS相似性達(dá)99%以上, 而基因序列決定功能, 推測基因在進(jìn)化過程中功能也高度保守。
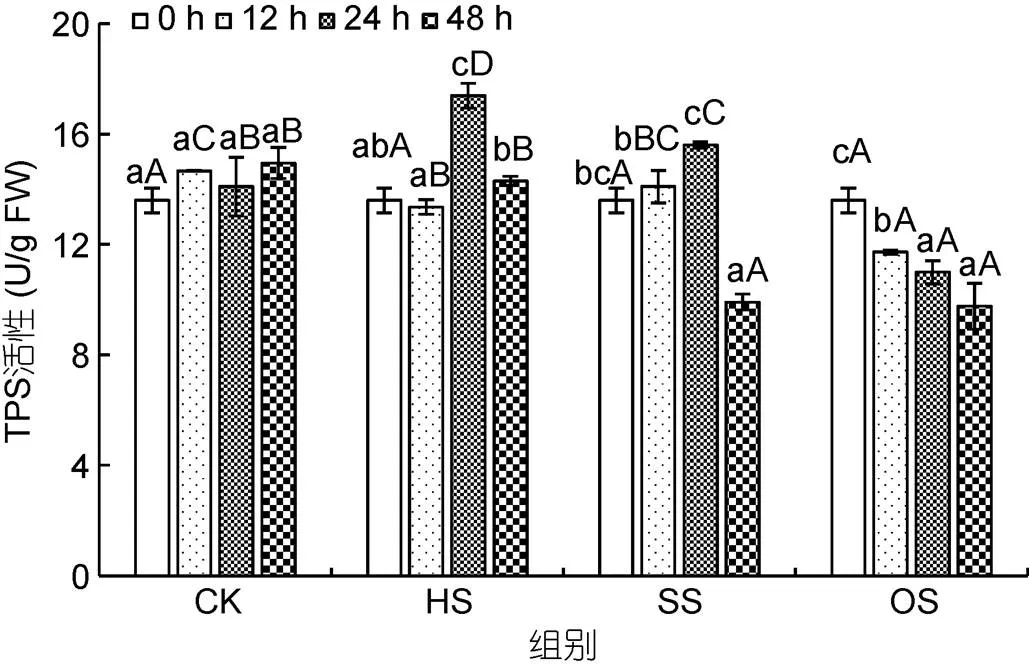
圖5 不同脅迫條件對龍須菜TPS活性的影響
大量基因家族在高等植物中擔(dān)負(fù)著不同功能, 隨著組織和環(huán)境的不同而具有不同的表達(dá)模式。如毛白楊()中只有10條基因在所有組織及氯化鈉、干旱等條件下能夠表達(dá); 擬南芥中2和3不表達(dá),4只在根和花中表達(dá), 其余8條基因在根、莖、葉等組織中均能表達(dá)(Yang, 2012)。相比于高等植物, 藻類中基因家族成員較少, 但這些基因在藻類的抗逆脅迫中也發(fā)揮了重要作用。如失水條件下海帶基因表達(dá)量在3 h最高, 可達(dá)到0 h的300倍(Deng, 2014); 壇紫菜中3條基因?qū)?9 °C高溫脅迫無應(yīng)答, 而在高度失水脅迫中發(fā)揮了應(yīng)激調(diào)節(jié)作用(史健志等, 2015)。本文中1—3 3條龍須菜基因在滲透脅迫下多被抑制, 與海帶、壇紫菜不同, 推測其原因可能與海帶、壇紫菜本身具有較強的脫水性有關(guān)。而對于高溫脅迫,1-3表達(dá)量雖上升但與對照組差異不顯著, 即對高溫?zé)o顯著應(yīng)答, 該結(jié)果與史健志等(2015)的研究結(jié)果類似, 可能是由于本文設(shè)置的30 °C高溫較低的原因。而且, 本文中TPS1蛋白與1基因在應(yīng)對高溫和高鹽脅迫時的響應(yīng)趨勢較為類似, 即對高溫?zé)o明顯響應(yīng), 而受高鹽脅迫的誘導(dǎo)。
在生物體內(nèi)海藻糖及其代謝物的含量變化與TPS等代謝酶的活性變化密切相關(guān)。如受熱激處理時金針菇()中TPS和TPP活性升高, 海藻糖酶活性下降, 海藻糖快速積累(Liu, 2016)。在干旱處理14 d的Desf中TPS和TPP活性上升, 同時海藻糖酶活性降低, 海藻糖含量增加(Ilhan, 2015)。與以上結(jié)果一致, 本文中TPS活性在高溫脅迫24 h時升高, 而海藻糖酶活性降低, 24 h時海藻糖含量開始顯著增加(Lv, 2019)。但在高鹽和滲透脅迫處理下, TPS活性與海藻糖含量無正相關(guān)性, 可能是因為海藻糖合成還受TPP的調(diào)控。
4 結(jié)論
海藻糖不僅是生物體內(nèi)碳儲存物質(zhì), 還可作為保護(hù)劑來抵抗不利環(huán)境脅迫。相比于大多數(shù)高等植物和紫菜, 龍須菜中海藻糖含量十分豐富, 因此海藻糖在龍須菜中可能具有重要的作用。龍須菜中4條基因序列高度保守, 推測為Class I亞家族。在基因轉(zhuǎn)錄、蛋白和酶活性水平上, TPS主要對高鹽和高溫脅迫響應(yīng)較大, 而對滲透脅迫不敏感或受其抑制, 該結(jié)果有助于更深入地了解海藻糖及其代謝酶在藻類抵御非生物脅迫中的作用, 為提高藻類抗逆性提供了理論資料。
史健志, 徐 燕, 紀(jì)德華等, 2015. 壇紫菜6-磷酸海藻糖合成酶(TPS)家族基因的克隆及表達(dá)特征分析. 水產(chǎn)學(xué)報, 39(4): 485—495
呂 燕, 汪芳俊, 林麗春等, 2019. 龍須菜中rbcL和hsp70對高溫和植物激素的響應(yīng). 水產(chǎn)學(xué)報, 43(4): 886—894
孫 鵬, 杜 宇, 呂 燕等, 2019. 龍須菜鹵代烷烴脫鹵酶基因的轉(zhuǎn)錄及原核表達(dá). 水產(chǎn)學(xué)報, 43(12): 2468—2475
張學(xué)成, 費修綆, 王廣策等, 2009. 江蘺屬海藻龍須菜的基礎(chǔ)研究與大規(guī)模栽培. 中國海洋大學(xué)學(xué)報, 39(5): 947—954
林麗春, 呂 燕, 孫 鵬等, 2019. 龍須菜()中兩種分支酸代謝酶對溫度和水楊酸的響應(yīng)及其原核表達(dá)研究. 海洋與湖沼, 50(1): 220—227
饒莉萍, 蘇文瑾, 劉 意等, 2020. 甘薯苯丙氨酸解氨酶基因IbPAL的克隆與表達(dá)分析. 植物科學(xué)學(xué)報, 38(3): 360—368
Bell W, Sun W N, Hohmann S, 1998. Composition and functional analysis of thetrehalose synthase complex. The Journal of Biological Chemistry, 273(50): 33311—33319
Chang L P, Sui Z H, Fu F, 2014. Relationship between gene expression of UDP-glucose pyrophosphorylase and agar yield in(Rhodophyta). Journal of Applied Phycology, 26(6): 2435—2441
Chen B B, Zou D H, Jiang H, 2015. Elevated CO2exacerbates competition for growth and photosynthesis betweenand. Aquaculture, 443: 49—55
Collén J, Porcel B, Carré W, 2013. Genome structure and metabolic features in the red seaweedshed light on evolution of the Archaeplastida. Proceedings of the National Academy of Sciences of the United States of America, 110(13): 5247—5252
Deng Y Y, Wang X L, Guo H, 2014. A trehalose-6-phosphate synthase gene from(Laminariales, Phaeophyceae). Molecular Biology Reports, 41(1): 529—536
Goddijn O J M, van Dun K, 1999. Trehalose metabolism in plants. Trends in Plant Science, 4(8): 315—319
Hottiger T, Schmutz P, Wiemken A, 1987. Heat-induced accumulation and futile cycling of trehalose in. Journal of bacteriology, 169(12): 5518—5522
Ilhan S, Ozdemir F, Bor M, 2015. Contribution of trehalose biosynthetic pathway to drought stress tolerance ofDesf. Plant Biology, 17(2): 402—407
Jiang H, Liu N N, Liu G L, 2016. Melanin production by a yeast strain XJ5-1 ofisolated from the Taklimakan desert and its role in the yeast survival in stress environments. Extremophiles, 20(4): 567—577
Krasensky J, Broyart C, Rabanal F A, 2014. The redox-sensitive chloroplast trehalose-6-phosphate phosphatase AtTPPD regulates salt stress tolerance. Antioxidants & Redox Signaling, 21(9): 1289—1304
Lee J M, Yang E C, Graf L, 2018. Analysis of the draft genome of the red seaweedprovides insights into genome size evolution in Rhodophyta. Molecular Biology and Evolution, 35(8): 1869—1886
Leyman B, Van Dijck P, Thevelein J M, 2001. An unexpected plethora of trehalose biosynthesis genes in. Trends in Plant Science, 6(11): 510—513
Li H W, Zang B S, Deng X W, 2011.Overexpression of the trehalose-6-phosphate synthase gene1 enhances abiotic stress tolerance in rice. Planta, 234(5): 1007—1018
Liu J H, Shang X D, Liu J Y, 2016. Changes in trehalose content, enzyme activity and gene expression related to trehalose metabolism inunder heat shock. Microbiology, 162(8): 1274—1285
Livak K J, Schmittgen T D, 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2?ΔΔCTmethod. Methods, 25(4): 402—408
Lv Y, Sun P, Zhang Y Y, 2019. Response of trehalose, its degrading enzyme, sucrose, and floridoside/isofloridoside under abiotic stresses in(Rhodophyta). Journal of Applied Phycology, 31(6): 3861—3869
O’Hara L E, Paul M J, Wingler A, 2013. How do sugars regulate plant growth and development? New insight into the role of trehalose-6-phosphate. Molecular Plant, 6(2): 261—274
Paul M J, Primavesi L F, Jhurreea D, 2008. Trehalose metabolism and signaling. Annual Review of Plant Biology, 59: 417—441
Pramanik M H R, Imai R, 2005. Functional identification of a trehalose 6-phosphate phosphatase gene that is involved in transient induction of trehalose biosynthesis during chilling stress in rice. Plant Molecular Biology, 58(6): 751—762
Richards A B, Krakowka S, Dexter L B, 2002. Trehalose: a review of properties, history of use and human tolerance, and results of multiple safety studies. Food and Chemical Toxicology, 40(7): 871—898
Schluepmann H, Van Dijken A, Aghdasi M, 2004. Trehalose mediated growth inhibition ofseedlings is due to trehalose-6-phosphate accumulation. Plant Physiology, 135(2): 879—890
Singh N K, Gupta D K, Jayaswal P K, 2012. The first draft of the pigeonpea genome sequence. Journal of Plant Biochemistry and Biotechnology, 21(1): 98—112
Sun X, Wu J, Wang G C, 2018. Genomic analyses of unique carbohydrate and phytohormone metabolism in the macroalga(Rhodophyta). BMC Plant Biology, 18(1): 94
Tang B, Wei P, Zhao L N, 2016. Knockdown of five trehalase genes using RNA interference regulates the gene expression of the chitin biosynthesis pathway in. BMC Biotechnology, 16(1): 67
Wahl V, Ponnu J, Schlereth A, 2013. Regulation of flowering by trehalose-6-phosphate signaling in. Science, 339(6120): 704—707
Wang G L, Zhao G, Feng Y B, 2010. Cloning and comparative studies of seaweed trehalose-6-phosphate synthase genes. Marine Drugs, 8(7): 2065—2079
Xie D W, Wang X N, Fu L S, 2015. Identification of the trehalose-6-phosphate synthase gene family in winter wheat and expression analysis under conditions of freezing stress. Journal of Genetics, 94(1): 55—65
Yang H L, Liu Y J, Wang C L, 2012. Molecular evolution of trehalose-6-phosphate synthase (TPS) gene family in,and rice. PLoS One, 7(8): e42438
Zang B S, Li H W, Li W J, 2011. Analysis of trehalose-6-phosphate synthase (TPS) gene family suggests the formation of TPS complexes in rice. Plant Molecular Biology, 76(6): 507—522
RESPONSE OF TREHALOSE-6-PHOSPHATE SYNTHASE (TPS) TO ABIOTIC STRESS IN
ZHANG Ying-Ying, LYU Yan, XUAN Wen-Yan, ZHANG Xiao-Qian, XU Nian-Jun, SUN Xue
(School of Marine Sciences, Ningbo University, Key Laboratory of Marine Biotechnology of Zhejiang Province, Ningbo 315211, China)
The non-reducing disaccharide trehalose and its metabolites are signaling molecules that regulate the growth, development, and stress response. In this paper, the responses of trehalose synthase (TPS) to high-temperature, high-salinity, and osmotic stresses were investigated at three levels of gene transcription, protein, and enzyme activity of seaweed. Four TPS sequences ofcontained both TPS-family conserved domains (Glyco-Transf-20) and TPP-family conserved domains (Trehalose-PPase), and belong to the Class I subfamily of TPS. At the transcriptional level, high-salinity stress mainly promoted the expression of1,2,and4, while1,2,and3 were almost suppressed under osmotic stress. At the protein level, TPS1 was promoted a 2.03-fold increment at 48 h under high-salinity compared with the control. TPS activity was increased at 24 h under high-temperature and high-salinity stresses, and was inhibited at 48 h under high-salinity stress and at 24—48 h under osmotic stress. In summary, the above results indicate that TPS was involved in the response of.to high-temperature and high-salinity stresses, but was not sensitive to osmotic stress. This study will provide a reference for improving the anti-adversity or breeding the stress-resistant varieties of.
; trehalose; trehalose-6-phosphate synthase; abiotic stress
* 國家重點研發(fā)計劃項目, 2018YFD0901502號; 國家自然科學(xué)基金項目, 31672674號; 浙江省自然科學(xué)基金項目, LY19C190003號, LQ20C190002號; 寧波市科技局項目, 2019B10009號。張瑩瑩, 碩士研究生, E-mail: 1113973798@qq.com
孫 雪, 博士, 研究員, E-mail: sunxue@nbu.edu.cn
2020-07-31,
2020-09-25
S968.43
10.11693/hyhz20200700225
