龍膽苦苷對脂多糖誘導血管內皮細胞損傷的作用及機制
2021-05-27 11:57:28胡志良丁水印
中草藥 2021年10期
胡志良,池 豪,丁水印
1.駐馬店市中心醫院 心血管內科,河南 駐馬店 463000
2.鄭州市中心醫院 心血管內科,河南 鄭州 450000
血管內皮細胞具有分泌細胞因子、維持血管結構等多種功能,在腫瘤、嚴重感染、心血管系統疾病發生過程中均發現有血管內皮細胞損傷[1]。病理條件下,細胞分泌大量的炎性因子,合成活性氧自由基,細胞過度凋亡[2]。脂多糖(lipopolysaccharide,LPS)促進血管內皮細胞凋亡、氧化應激和炎性因子分泌[3]。龍膽苦苷提取自龍膽科植物,具有抗氧化、抗炎、殺菌等作用,臨床上主要用于抗炎、健胃、保肝以及抗病原微生物的治療[4]。研究發現,龍膽苦苷能夠改善氧化應激下血管內皮細胞氧化損傷,龍膽苦苷可能具有血管內皮損傷保護作用[5]。龍膽苦苷能夠通過抑制核因子-κΒ(nuclear factor-κΒ,NF-κΒ)信號通路,從而抑制氧化應激和炎性反應[6]。人體內腦、肝、心、肺等多種組織均表達NF-κΒ,NF-κΒ負責調控氧化應激、細胞凋亡和炎性反應[7]。LPS可激活血管內皮細胞NF-κΒ信號通路,抑制NF-κΒ信號通路能夠保護血管內皮細胞損傷[8]。本研究考察龍膽苦苷對LPS誘導的血管內皮細胞損傷的作用及機制,為其治療感染等因素誘導的血管內皮損傷提供依據。
1 材料
1.1 細胞
大鼠主動脈血管內皮細胞RAEC購自上海中喬新舟生物科技有限公司。
1.2 藥品與試劑
龍膽苦苷(批號L-005,質量分數>98%)購自成都瑞芬思生物科技有限公司;丙二醛檢測試劑盒(批號BC0025)、超氧化物歧化酶(superoxide dismutase,SOD)檢測試劑盒(批號BC0175)、膜聯蛋白V(Annexin V)-異硫氰酸熒光素(fluorescein isothiocyanate isomer,FITC)/碘化丙錠(propidium iodide,PI)細胞凋亡檢測試劑盒(批號CA1020)購自北京索萊寶科技有限公司;兔抗磷酸化p65(p-p65)抗體、兔抗剪切型半胱氨酸蛋白酶-3(cleaved Caspase-3)抗體、兔抗甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)抗體、HRP標記的山羊抗兔抗體購自英國Abcam公司;RPMI 1640培養基(批號R8758)、胎牛血清(批號F2442)購自美國Sigma公司;PMA(批號GT6290)購自北京華越洋生物科技有限公司;引物由武漢金開瑞生物工程有限公司合成。
1.3 儀器
Multiscan MK3酶標儀(美國Thermo Fisher Scientific公司);FC500流式細胞儀(美國Beckman公司);5424離心機、5810R離心機(德國Eppendrof公司);LightCycler480熒光定量PCR儀(美國Roche公司);PowerPac蛋白電泳儀、Gel Doc XR+凝膠顯示系統(美國Bio-Rad公司)。
2 方法
2.1 細胞培養
RAEC細胞用含10%胎牛血清的RPMI 1640培養基,于37 ℃、5% CO2培養箱中培養。
2.2 龍膽苦苷對RAEC細胞活力的影響
取處于對數生長期的RAEC細胞,以3×103/孔接種至96孔板中,培養12 h。設置對照組和龍膽苦苷(5、10、20、40、80 μmol/L)組,各給藥組加入相應藥物,對照組加入不含藥物的培養基,培養24 h。每孔加入10 μL CCK-8工作液,孵育2 h,采用酶標儀檢測490 nm處的吸光度(A)值。
2.3 龍膽苦苷對RAEC細胞增殖的影響
取處于對數生長期的RAEC細胞,以3×103/孔接種至96孔板中,培養12 h。設置對照組、模型組和龍膽苦苷(5、10、20 μmol/L)組,模型組和各給藥組加入LPS(100 μg/mL),各給藥組再加入相應藥物,對照組加入不含藥物的培養基,培養24 h。按“2.2”項下方法檢測各組細胞活力。
2.4 龍膽苦苷對RAEC細胞凋亡的影響
取處于對數生長期的RAEC細胞,以1×103/孔接種至6孔板中,培養12 h。設置對照組、模型組和龍膽苦苷(5、10、20 μmol/L)組,模型組和各給藥組加入LPS(100 μg/mL),各給藥組再加入相應藥物,對照組加入不含藥物的培養基,培養24 h。收集細胞,以PBS溶液重懸,制成5×105/mL細胞懸液,加入5 μL Annexin V-FITC和5 μL PI染色液,1 h內采用流式細胞儀測定各組細胞凋亡情況。
2.5 龍膽苦苷對RAEC細胞丙二醛水平和SOD活性的影響
按“2.4”項下方法處理細胞,按照試劑盒說明書檢測各組細胞丙二醛水平和SOD活性。
2.6 龍膽苦苷對RAEC細胞腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)、白細胞介素-6(interleukin-6,IL-6)和IL-1β mRNA表達的影響
按“2.4”項下方法處理細胞,按照試劑盒說明書提取細胞總RNA并合成cDNA,進行qRT-PCR分析。引物序列:TNF-α上游引物5’-TGACAAGCCTGTAGCCCATGTT-3’,下游引物5’-AGGGCAATGATCCCAAAGTAGA-3’;IL-6上游引物5’-CCAGTACCCCCAGGAGAAGAT-3’,下游引物5’-TTGCCTTTTTCTGCAGGAAC-3’;IL-1β上游引物5’-AGTTGCCTTCTTGGGACTGA-3’,下游引物5’-TCCACGATTTCCCAGAGAAC-3’;GAPDH上游引物5’-CCAGCAAAGAGACCAAGAGGAA-3’,下游引物5’-ATGGTACATAGACAAGGTGCGG-3’。
2.7 龍膽苦苷對RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響
按“2.4”項下方法處理細胞,收集細胞,加入RIPA裂解液,4 ℃、12 000×g離心10 min,收集上清液,采用BCA蛋白定量試劑盒測定蛋白質量濃度。蛋白樣品經10%十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳,轉至PVDF膜,于5%牛血清白蛋白中封閉1 h,分別加入p-p65和cleaved Caspase-3抗體(1∶600),4 ℃孵育過夜;加入HRP標記的山羊抗兔抗體(1∶4000),室溫孵育2 h,加入ECL發光液顯影。
2.8 NF-κB信號激活劑(PMA)對龍膽苦苷促進RAEC細胞增殖的影響
取處于對數生長期的RAEC細胞,以3×103/孔接種至96孔板中,培養12 h。設置對照組、模型組、龍膽苦苷(10 μmol/L)組和龍膽苦苷(10 μmol/L)聯合PMA(1 μmol/L)組,模型組和各給藥組加入LPS(100 μg/mL),各給藥組再加入相應藥物,對照組加入不含藥物的培養基,培養24 h。按“2.2”項下方法檢測各組細胞活力。
2.9 PMA對龍膽苦苷抑制RAEC細胞凋亡的影響
取處于對數生長期的RAEC細胞,以1×103/孔接種至6孔板中,培養12 h。設置對照組、模型組、龍膽苦苷(10 μmol/L)組和龍膽苦苷(10 μmol/L)聯合PMA(1 μmol/L)組,模型組和各給藥組加入LPS(100 μg/mL),各給藥組再加入相應藥物,對照組加入不含藥物的培養基,培養24 h。按“2.4”項下方法檢測各組細胞凋亡情況。
2.10 PMA對龍膽苦苷抑制RAEC細胞丙二醛水平、升高SOD活性的影響
按“2.8”項下方法處理細胞,按“2.5”項下方法檢測各組細胞丙二醛水平和SOD活性。
2.11 PMA對龍膽苦苷抑制RAEC細胞TNF-α、IL-6和IL-1β mRNA表達水平的影響
按“2.8”項下方法處理細胞,按“2.6”項下方法檢測各組細胞TNF-α、IL-6和IL-1βmRNA表達情況。
2.12 PMA對龍膽苦苷抑制RAEC細胞p-p65和cleaved Caspase-3蛋白表達水平的影響
按“2.8”項下方法處理細胞,按“2.7”項下方法檢測各組細胞p-p65和cleaved Caspase-3蛋白表達情況。
2.13 統計學分析
采用SPSS 21.0軟件分析數據,計量資料以±s表示,用獨立樣本t檢驗和單因素方差比較。
3 結果
3.1 龍膽苦苷對RAEC細胞活力的影響
如表1所示,與對照組比較,龍膽苦苷(5、10、20 μmol/L)組細胞活力無明顯變化,龍膽苦苷(40、80 μmol/L)組細胞活力顯著降低(P<0.05)。因此,選擇5、10、20 μmol/L龍膽苦苷進行后續研究。
表1 龍膽苦苷對RAEC細胞活力的影響 (±s ,n=3)Table 1 Effect of gentiopicroside onviability of RAEC cells (±s ,n=3)

表1 龍膽苦苷對RAEC細胞活力的影響 (±s ,n=3)Table 1 Effect of gentiopicroside onviability of RAEC cells (±s ,n=3)
與對照組比較:*P<0.05*P < 0.05 vs control group
組別 劑量/(μmol·L?1) A490對照 — 0.65±0.05龍膽苦苷 5 0.63±0.07 10 0.60±0.08 20 0.61±0.05 40 0.50±0.04*80 0.32±0.03*
3.2 龍膽苦苷對RAEC細胞增殖和凋亡的影響
如圖1和表2所示,與對照組比較,模型組細胞活力顯著降低(P<0.05),細胞凋亡率顯著升高(P<0.05);與模型組比較,龍膽苦苷組細胞活力顯著升高(P<0.05),細胞凋亡率顯著降低(P<0.05),呈劑量相關性,表明龍膽苦苷可促進LPS誘導的RAEC細胞增殖并抑制其凋亡。

圖1 龍膽苦苷對RAEC細胞凋亡的影響Table 1 Effect of gentiopicroside on apoptosis of RAEC cells
表2 龍膽苦苷對RAEC細胞增殖和凋亡的影響(±s ,n=3)Table 2 Effectof gentiopicroside on proliferation and apoptosis of RAEC cells (±s ,n=3)
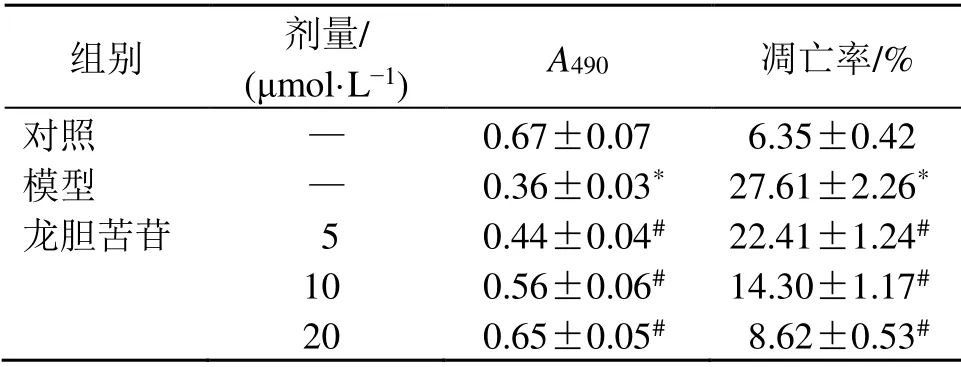
表2 龍膽苦苷對RAEC細胞增殖和凋亡的影響(±s ,n=3)Table 2 Effectof gentiopicroside on proliferation and apoptosis of RAEC cells (±s ,n=3)
與對照組比較:*P<0.05;與模型組比較:#P<0.05,下表同*P < 0.05 vs control group; #P < 0.05 vs model group,same as below tables
組別 劑量/(μmol·L?1) A490 凋亡率/%對照 — 0.67±0.07 06.35±0.42模型 — 0.36±0.03* 27.61±2.26*龍膽苦苷 5 0.44±0.04# 22.41±1.24#10 0.56±0.06# 14.30±1.17#20 0.65±0.05# 08.62±0.53#
3.3 龍膽苦苷對RAEC細胞丙二醛水平和SOD活性的影響
如表3所示,與對照組比較,模型組細胞丙二醛水平顯著升高(P<0.05),SOD活性顯著降低(P<0.05);與模型組比較,龍膽苦苷組細胞丙二醛水平顯著降低(P<0.05),SOD活性顯著升高(P<0.05),呈劑量相關性,表明龍膽苦苷可抑制LPS誘導的RAEC細胞氧化應激。
表3 龍膽苦苷對RAEC細胞丙二醛水平和SOD活性的影響 (±s ,n=3)Table 3 Effect of gentiopicroside on malondialdehyde level and SOD activity in RAEC cells (±s ,n=3)
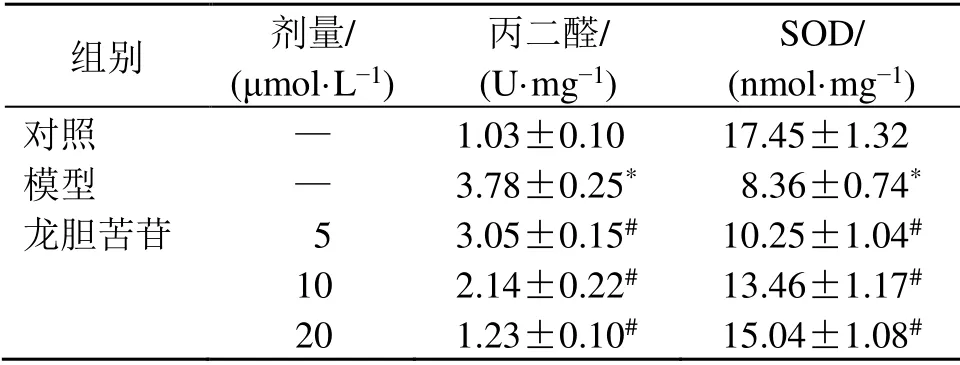
表3 龍膽苦苷對RAEC細胞丙二醛水平和SOD活性的影響 (±s ,n=3)Table 3 Effect of gentiopicroside on malondialdehyde level and SOD activity in RAEC cells (±s ,n=3)
組別 劑量/(μmol·L?1)丙二醛/(U·mg?1)SOD/(nmol·mg?1)對照 — 1.03±0.10 17.45±1.32模型 — 3.78±0.25* 08.36±0.74*龍膽苦苷 5 3.05±0.15# 10.25±1.04#10 2.14±0.22# 13.46±1.17#20 1.23±0.10# 15.04±1.08#
3.4 龍膽苦苷對RAEC細胞TNF-α、IL-6和IL-1β mRNA表達的影響
如表4所示,與對照組比較,模型組細胞TNF-α、IL-6和IL-1βmRNA表達水平顯著升高(P<0.05);與模型組比較,龍膽苦苷組細胞TNF-α、IL-6和IL-1βmRNA表達水平顯著降低(P<0.05),呈劑量相關性,表明龍膽苦苷可抑制LPS誘導的RAEC細胞炎性因子的表達。
表4 龍膽苦苷對RAEC 細胞TNF-α、IL-6和IL-1β mRNA表達的影響 (±s ,n=3)Table 4 Effect of gentiopicroside on mRNA expressions of TNF-α,IL-6 and IL-1β in RAEC cells (±s ,n=3)

表4 龍膽苦苷對RAEC 細胞TNF-α、IL-6和IL-1β mRNA表達的影響 (±s ,n=3)Table 4 Effect of gentiopicroside on mRNA expressions of TNF-α,IL-6 and IL-1β in RAEC cells (±s ,n=3)
組別 劑量/(μmol·L?1) mRNA相對表達量TNF-α IL-6 IL-1β對照 — 1.00±0.09 1.00±0.11 1.00±0.11模型 — 5.17±0.42* 9.65±0.93* 4.16±0.32*龍膽苦苷 5 4.12±0.36# 7.51±0.62# 2.95±0.23#10 3.01±0.21# 5.04±0.52# 2.20±0.14#20 1.78±0.12# 3.71±0.28# 1.64±0.17#
3.5 龍膽苦苷對RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響
如圖2和表5所示,與對照組比較,模型組細胞p-p65和cleaved Caspase-3蛋白表達水平顯著升高(P<0.05);與模型組比較,龍膽苦苷組細胞p-p65和cleaved Caspase-3蛋白表達水平顯著降低(P<0.05),呈劑量相關性,表明龍膽苦苷可抑制LPS誘導的RAEC細胞NF-κΒ信號通路的激活,并抑制細胞凋亡。
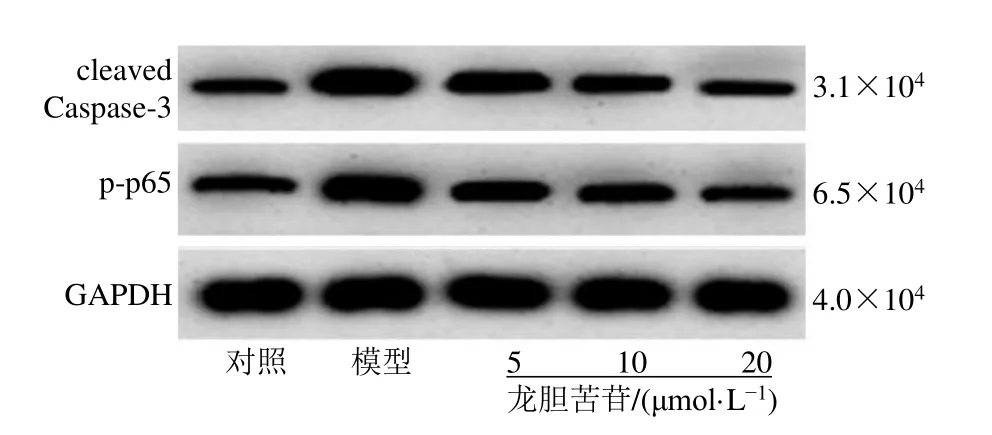
圖2 龍膽苦苷對RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響Fig.2 Effect of gentiopicroside on expressions of p-p65 and cleaved Caspase-3 in RAEC cells
表5 龍膽苦苷對RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響 (±s ,n=3)Table 5 Effect of gentiopicroside on expressions of p-p65 and cleaved Caspase-3 in RAEC cells (±s ,n=3)
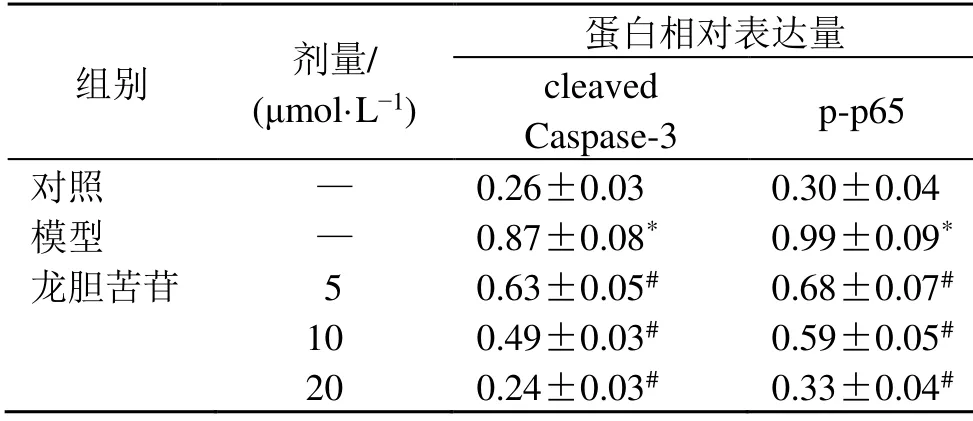
表5 龍膽苦苷對RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響 (±s ,n=3)Table 5 Effect of gentiopicroside on expressions of p-p65 and cleaved Caspase-3 in RAEC cells (±s ,n=3)
組別 劑量/(μmol·L?1)蛋白相對表達量cleaved Caspase-3 p-p65對照 — 0.26±0.03 0.30±0.04模型 — 0.87±0.08* 0.99±0.09*龍膽苦苷 5 0.63±0.05# 0.68±0.07#10 0.49±0.03# 0.59±0.05#20 0.24±0.03# 0.33±0.04#
3.6 PMA對龍膽苦苷促進RAEC細胞增殖以及抑制RAEC細胞凋亡的影響
如圖3和表6所示,與龍膽苦苷組比較,龍膽苦苷聯合PMA組細胞活力顯著降低(P<0.05),細胞凋亡率顯著升高(P<0.05)。
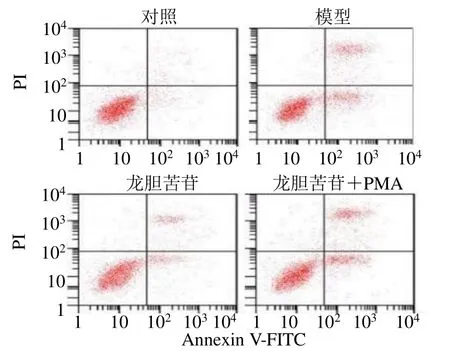
圖3 PMA對龍膽苦苷抑制血管內皮細胞凋亡的影響Fig.3 Effect of PMA on inhibiting RAEC cells apoptosis by gentiopicroside
表6 PMA對龍膽苦苷促進血管內皮細胞增殖以及抑制細胞凋亡的影響 (±s ,n=3)Table 6 Effect of PMA on promoting RAEC cells proliferation and inhibiting cells apoptosis by gentiopicroside (±s ,n=3)
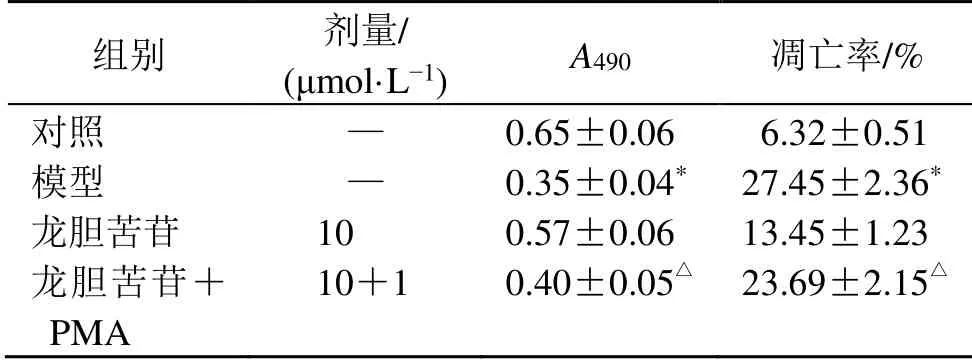
表6 PMA對龍膽苦苷促進血管內皮細胞增殖以及抑制細胞凋亡的影響 (±s ,n=3)Table 6 Effect of PMA on promoting RAEC cells proliferation and inhibiting cells apoptosis by gentiopicroside (±s ,n=3)
與對照組比較:*P<0.05;與龍膽苦苷組比較:△P<0.05,下表同*P < 0.05 vs control group; △P < 0.05 vs gentiopicroside group,same as below tables
組別 劑量/(μmol·L?1) A490 凋亡率/%對照 — 0.65±0.06 06.32±0.51模型 — 0.35±0.04* 27.45±2.36*龍膽苦苷 10 0.57±0.06 13.45±1.23龍膽苦苷+PMA 10+1 0.40±0.05△ 23.69±2.15△
3.7 PMA對龍膽苦苷抑制RAEC細胞丙二醛水平、升高SOD活性的影響
如表7所示,與龍膽苦苷組比較,龍膽苦苷聯合PMA組細胞丙二醛水平顯著升高(P<0.05),SOD活性顯著降低(P<0.05)。
表7 PMA對龍膽苦苷抑制RAEC細胞丙二醛水平、升高SOD活性的影響 (±s ,n=3)Table 7 Effect of PMA on inhibiting malondialdehyde level and increasing SOD activity of RAEC cells by gentiopicroside (±s ,n=3)
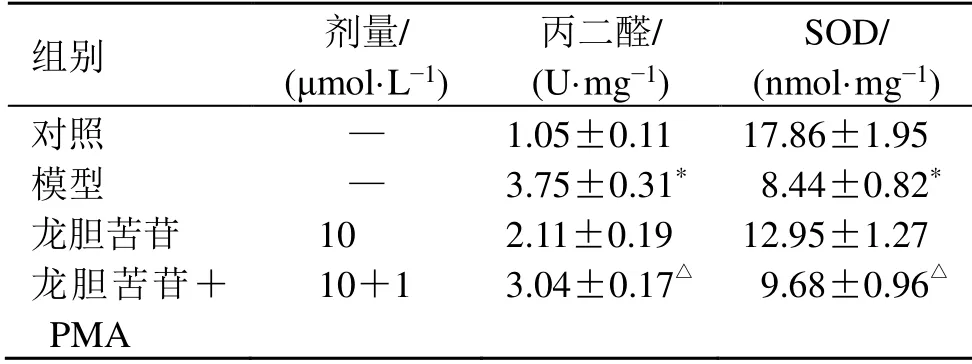
表7 PMA對龍膽苦苷抑制RAEC細胞丙二醛水平、升高SOD活性的影響 (±s ,n=3)Table 7 Effect of PMA on inhibiting malondialdehyde level and increasing SOD activity of RAEC cells by gentiopicroside (±s ,n=3)
組別 劑量/(μmol·L?1)丙二醛/(U·mg?1)SOD/(nmol·mg?1)對照 — 1.05±0.11 17.86±1.95模型 — 3.75±0.31* 08.44±0.82*龍膽苦苷 10 2.11±0.19 12.95±1.27龍膽苦苷+PMA 10+1 3.04±0.17△ 09.68±0.96△
3.8 PMA對龍膽苦苷抑制RAEC細胞TNF-α、IL-6和IL-1β mRNA表達水平的影響
如表8所示,與龍膽苦苷組比較,龍膽苦苷聯合PMA組細胞TNF-α、IL-6和IL-1βmRNA表達水平顯著升高(P<0.05)。
表8 PMA對龍膽苦苷抑制RAEC細胞TNF-α、IL-6和IL-1β mRNA表達水平的影響 (±s ,n=3)Table 8 Effect of PMA on inhibiting mRNA expressions of TNF-α,IL-6 and IL-1β in RAEC cells by gentiopicroside(±s ,n=3)

表8 PMA對龍膽苦苷抑制RAEC細胞TNF-α、IL-6和IL-1β mRNA表達水平的影響 (±s ,n=3)Table 8 Effect of PMA on inhibiting mRNA expressions of TNF-α,IL-6 and IL-1β in RAEC cells by gentiopicroside(±s ,n=3)
組別 劑量/(μmol·L?1) mRNA相對表達量TNF-α IL-6 IL-1β對照 — 1.00±0.12 1.00±0.10 1.00±0.09模型 — 4.78±0.41* 9.32±0.74* 4.11±0.36*龍膽苦苷 10 2.95±0.25 4.84±0.41 2.26±0.21龍膽苦苷+PMA 10+1 3.20±0.21△ 6.98±0.15△ 3.25±0.10△
3.9 PMA對龍膽苦苷抑制RAEC細胞p-p65和cleaved Caspase-3蛋白表達水平的影響
如圖4和表9所示,與龍膽苦苷組比較,龍膽苦苷聯合PMA組細胞p-p65和cleaved Caspase-3蛋白表達水平顯著升高(P<0.05)。以上結果表明,PMA能夠抑制龍膽苦苷改善RAEC細胞損傷的作用。

圖4 PMA對龍膽苦苷抑制RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響Fig.4 Effect of PMA on inhibiting expressions of p-p65 and cleaved Caspase-3 in RAEC cells by gentiopicroside
表9 PMA對龍膽苦苷抑制RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響 (±s ,n=3)Table 9 Effect of PMA on inhibiting expressions of p-p65 and cleaved Caspase-3 in RAEC cells by gentiopicroside(±s ,n=3)
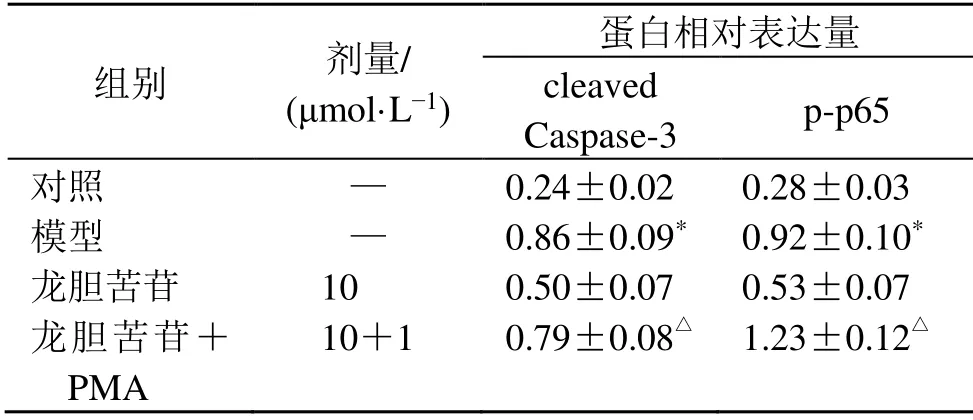
表9 PMA對龍膽苦苷抑制RAEC細胞p-p65和cleaved Caspase-3蛋白表達的影響 (±s ,n=3)Table 9 Effect of PMA on inhibiting expressions of p-p65 and cleaved Caspase-3 in RAEC cells by gentiopicroside(±s ,n=3)
組別 劑量/(μmol·L?1)蛋白相對表達量cleaved Caspase-3 p-p65對照 — 0.24±0.02 0.28±0.03模型 — 0.86±0.09* 0.92±0.10*龍膽苦苷 10 0.50±0.07 0.53±0.07龍膽苦苷+PMA 10+1 0.79±0.08△ 1.23±0.12△
4 討論
龍膽苦苷為裂環環烯醚萜化合物,能夠抑制氧化應激和炎性反應[4]。龍膽苦苷能夠抑制氧化應激誘導的血管內皮細胞損傷和細胞凋亡[5]。本研究結果顯示,與模型組比較,龍膽苦苷組細胞活力升高,細胞凋亡率降低,提示龍膽苦苷可能具有改善LPS誘導的血管內皮細胞損傷的作用。
LPS刺激血管內皮細胞損傷與氧化應激、炎性反應有關[9]。LPS誘導的血管內皮細胞中抗氧化酶的活性下降,抗氧化酶活性降低能夠直接誘導氧自由基的大量聚集,過量的氧自由基能夠刺激細胞,誘導脂質發生過氧化,導致氧化損傷;還能夠激活細胞內的凋亡反應,促進細胞凋亡[10-11]。丙二醛水平可直接反應細胞氧化損傷程度[12]。SOD是氧自由基清除劑,也是主要的抗氧化酶之一[13]。Caspase蛋白家族被激活后形成凋亡級聯反應,誘導細胞凋亡;Caspase-3是Caspase凋亡級聯反應的下游執行因子,其被活化后是細胞凋亡的標志[14-15]。LPS誘導血管內皮細胞分泌炎性因子,促進炎性反應的產生并誘導細胞凋亡[16]。血管內皮細胞分泌TNF-α、IL-6、IL-1β等炎性因子,誘導炎性損傷[17-19]。氧自由基過度積累可誘導炎性反應,并且炎性因子也可促進氧化應激[20-21]。本研究結果顯示,模型組細胞丙二醛水平升高,SOD活性降低,cleaved Caspase-3蛋白表達水平降低,TNF-α、IL-6和IL-1βmRNA表達水平升高;龍膽苦苷能夠顯著抑制LPS誘導的血管內皮細胞炎性因子表達和氧化損傷,抑制細胞凋亡。
龍膽苦苷能夠通過抑制NF-κΒ信號通路的激活發揮抗損傷作用[6]。本研究結果顯示,龍膽苦苷組細胞p-p65蛋白表達水平降低。p65是NF-κΒ信號通路的關鍵亞單位,其表達量與NF-κΒ信號通路的激活水平有關[22]。NF-κΒ在炎性反應、細胞生長和氧化應激等過程中均有調控作用[23]。血管內皮損傷中NF-κΒ信號異常激活,抑制NF-κΒ信號通路能夠緩解LPS誘導的血管內皮細胞損傷[7]。本研究結果顯示,PMA能夠抑制龍膽苦苷改善RAEC細胞損傷的作用,表明龍膽苦苷能夠通過NF-κΒ信號通路發揮血管內皮細胞損傷保護作用。
綜上,龍膽苦苷能夠改善LPS誘導的血管內皮細胞損傷,其機制與抑制NF-κΒ信號通路有關。
利益沖突所有作者均聲明不存在利益沖突
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
媽媽寶寶(2017年3期)2017-02-21 01:22:28
光學精密工程(2016年6期)2016-11-07 09:07:19
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
核科學與工程(2015年4期)2015-09-26 11:59:03
