廣藿香青枯病菌致病相關基因RsgidA的克隆及生物信息學分析
2021-05-27 11:57:38王亞琴張宇瑤黎廣衛
中草藥 2021年10期
李 巧,王亞琴,賀 紅*,張 泳,張宇瑤,黎廣衛
1.廣州中醫藥大學中藥學院,廣東 廣州 510006
2.廣州中醫藥大學 中藥資源科學與工程研究中心,廣東 廣州 510006
廣藿香Pogostemon cablin(Blanco) Benth.為唇形科刺蕊草屬植物,以干燥地上部分入藥,其味辛,性微溫,具有芳香化濁,和中止嘔,發表解暑等功能,為廣東道地藥材、“十大廣藥”之一。廣藿香在種植過程中,易感染青枯病。青枯病是一種細菌性土傳病害,病原菌在植物維管束內定殖和繁殖,破壞寄主植物的輸導系統,使植株無法正常吸收水分和養分導致枯萎死亡[1]。青枯病嚴重影響廣藿香的產量和質量,危及廣藿香的生存和發展。發現和研究病原菌致病相關基因,有助于了解病原菌致病機理、開發抗菌的靶標藥物及發展新的防病策略。
轉座子(transposon,Tn)是DNA插入因子的一種,是一類可以在基因組中移動的DNA片段[2]。目前細菌轉座子Tn5的轉座機制已闡明,利用Tn5轉座突變技術,將轉座子插入到細菌基因組中,使得插入位點處基因發生突變并導致其相關功能活性喪失,從而篩選出功能發生突變的菌株[3]。如果轉座子插入位點基因是病原菌致病相關基因,就可能導致病原菌致病力喪失或減弱,由此可發現致病相關基因[4-5]。本課題組前期從感染了青枯病的廣藿香植株中分離得到了青枯菌菌株Ralstonia solanacearumPRS-84[6],并利用Tn5轉座子插入突變及致病性篩選獲得了菌株PRS-84的低致病力突變株[7]。本研究以低致病力突變株PRS-84-4-7為實驗材料,對其進行致病性驗證,再以插入的Tn5轉座子序列為基礎,利用反向PCR擴增獲得轉座子插入位點的側翼序列,并對插入位點基因進行克隆及生物信息學分析,為了解廣藿香青枯病菌的致病機制、發展新的防病策略奠定基礎。
1 材料
廣藿香青枯菌野生菌株Ralstonia solanacearumPRS-84(簡稱PRS-84)由劉丹等[6]從感染了青枯病的廣藿香植株中分離獲得。廣藿香青枯菌低致病力突變株PRS-84-4-7由王亞琴等[7]構建及篩選獲得。
2 方法
2.1 青枯菌的培養
將廣藿香青枯菌菌株PRS-84及其突變株PRS-84-4-7從?80 ℃取出,采用劃線法將其接種至NA固體培養基培養,挑取單菌落并將其接種于NA液體培養基中,28 ℃、200 r/min恒溫振蕩培養24 h。
2.2 致病性測定
對野生菌株PRS-84及低致病力突變株PRS-84-4-7進行培養,將菌液稀釋至5.0×108cfu/mL;以無菌水為對照,采用傷根浸泡法,將稀釋后的菌液分別接種至廣藿香植株,再將實驗植株移至塑料棚中培養,濕度85%左右,溫度28~30 ℃。觀察植株的發病情況。
2.3 反向PCR擴增及序列測定
提取低致病力突變株PRS-84-4-7基因組DNA,用限制性內切酶Hind Ⅲ對基因組DNA進行酶切,用T4連接酶對酶切產物進行連接。根據Tn5轉座子序列設計的引物KAN-2 FP-1(5’-ACCTACAACAAAGCTCTCATCAACC-3’)及KAN-2 RP-1(5’-GCAATGTAACATCAGAGATTTTGAG-3’),對連接產物進行反向PCR擴增,以獲得突變株轉座子插入位點的側翼序列。反應體系:連接產物1.0 μL;引物(10 μmol/L)各1.0 μL;Ex Taq酶12.5 μL;ddH2O 9.5 μL。反應條件:94 ℃預變性5 min;94 ℃變性30 s,58 ℃退火30 s,72 ℃延伸2 min,35個循環;72 ℃延伸7 min。對PCR產物進行瓊脂糖凝膠電泳檢測,并對擴增產物進行序列測定。
2.4 RsgidA基因的克隆及序列測定
根據Tn5轉座子插入位點側翼序列BLAST比對獲得的同源基因序列設計特異性引物:4-7-F為CCCTCGAGTGTTCCGTCTTTTTTTTCTGGT(下劃線為XhoⅠ酶切位點),4-7-R:CGAGCTCTGATGCTTAGGAAAATTCCAGATA(下劃線為SacⅠ酶切位點)。分別以野生菌株及突變菌株基因組DNA為模板,以4-7-F及4-7-R為引物對目的基因進行PCR擴增。反應體系:高保真預混合酶45 μL;引物(10μmol/L)各2.0 μL;DNA模板1.0 μL。反應條件:98 ℃預變性2 min;98 ℃變性10 s,58 ℃退火15 s,72 ℃延伸1 min,35個循環;72 ℃延伸3 min。
取PCR產物進行瓊脂糖凝膠電泳檢測,用凝膠成像儀觀察,并對擴增產物進行切膠回收。將純化產物與T載體(pClone007 simple vector)連接,構建重組質粒,采用熱激法將重組質粒轉化至大腸桿菌感受態細胞,將轉化后的菌液涂布于含有氨芐青霉素(100 μg/mL)的LB固體培養基,過夜培養。挑取抗性菌落,進行PCR鑒定,進一步提取陽性克隆質粒并經XhoⅠ及SacⅠ雙酶切鑒定和測序。
2.5 生物信息學分析
將測序獲得的核苷酸序列通過ExPASy工具翻譯成氨基酸序列;以ProtParam在線工具對蛋白質進行理化性質預測;以ProtScale軟件對蛋白質進行親疏水性預測;以SignalP 5.0軟件對蛋白質進行信號肽預測;以TMHMM軟件對蛋白質進行跨膜結構預測;以SOPMA軟件對蛋白質進行二級結構預測;以SWISS-MODEL對蛋白質進行三級結構預測;使用NCBI中的BLAST工具搜索并下載其他植物病原中的同源氨基酸序列,利用MEGA5.1軟件中的鄰接法構建系統進化樹。
3 結果與分析
3.1 低致病力突變菌株對廣藿香的致病性表型
分別用野生菌株PRS-84及突變株PRS-84-4-7對廣藿香植株進行傷根浸泡處理,以用無菌水浸泡的植株為對照,觀察植株在7 d內的發病情況。圖1為接種第3天的植株表型,可以看出,無菌水處理的植株保持健康,接種野生菌株PRS-84的廣藿香植株出現葉片下垂、萎蔫等典型的青枯病癥狀,而接種突變株PRS-84-4-7的植株無明顯的發病癥狀。結果表明,與野生菌株PRS-84相比,突變株PRS-84-4-7的致病性明顯減弱,符合低致病力突變株的表型特征。

圖1 低致病力突變菌株與野生菌株對廣藿香的致病性表型(接種第3天)Fig.1 Pathogenicity assays of low virulent mutant and wild-type strain on patchouli plants (at 3rd day postinoculation)
3.2 低致病力突變株Tn5轉座子插入位點側翼序列的擴增及序列測定
根據Tn5轉座子序列設計的特異性引物對突變株PRS-84-4-7基因組DNA進行反向PCR擴增,電泳結果顯示(圖2),該菌株擴增出特異性條帶,說明Tn5轉座子成功插入至廣藿香青枯菌PRS-84的基因組中。對Tn5轉座子插入位點側翼序列進行測序及分析,結果顯示,轉座子插入位點基因與gidA(glucose-inhibited division protein A)基因(Genbank accession No.CP049363.1)的相似度最高,將該基因命名為廣藿香青枯病菌RsgidA基因。
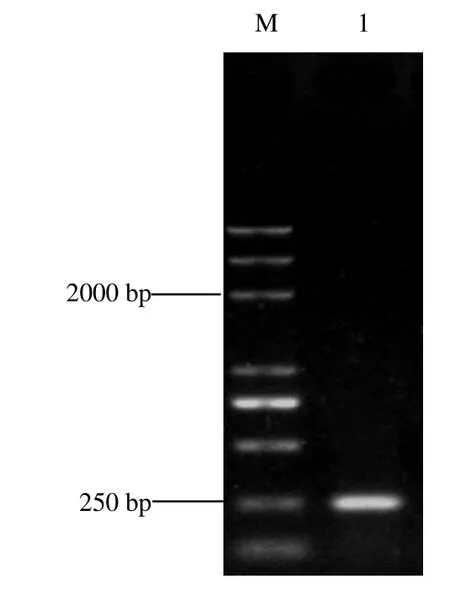
圖2 低致病力突變株Tn5轉座子插入位點側翼序列的反向PCR檢測Fig.2 Gel electrophoresis of inverse PCR products of flanking sequences adjacent to Tn5 transposon insertion site in low virulent mutant
3.3 廣藿香青枯病菌RsgidA基因的克隆
根據Tn5轉座子插入位點側翼序列BLAST比對獲得的同源基因序列設計特異性引物4-7-F/4-7-R,以野生菌株PRS-84和突變株PRS-84-4-7基因組DNA為模板,對插入突變基因進行PCR擴增,電泳結果顯示(圖3),突變株擴增的條帶約為3000 bp,較野生菌株長1000 bp左右,因Tn5轉座子全長為1221 bp,與預期片段大小一致。將菌株PRS-84的PCR擴增產物進行切膠純化回收,將其與T載體連接后轉化至大腸桿菌感受態細胞,經菌落PCR及雙酶切鑒定獲得陽性克隆子,表明成功克隆了廣藿香青枯病菌RsgidA基因。
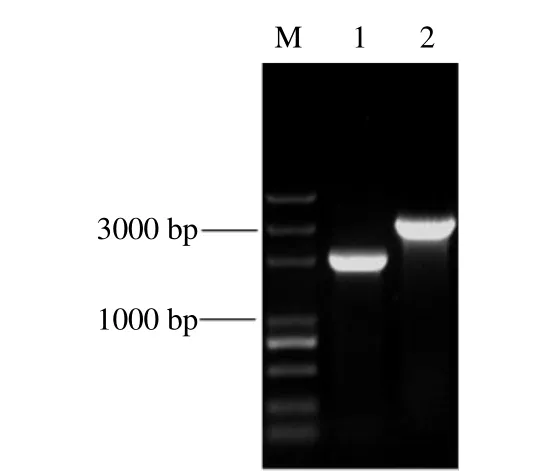
圖3 低致病力突變株Tn5轉座子插入突變基因的PCR檢測Fig.3 Gel electrophoresis of PCR products of Tn5 transposon insertion-site gene in low virulent mutant PRS-84-4-7
3.4 廣藿香青枯病菌RsgidA基因的生物信息學分析
3.4.1RsgidA基因的序列分析 對“3.3”項中獲得的陽性克隆子進行培養,提取質粒,測序獲得RsgidA基因的全長序列。該基因全長從起始密碼子ATG到終止密碼子TAA為1881 bp,共編碼626個氨基酸(圖4)。

圖4 RsgidA基因的核苷酸序列及推測的氨基酸序列Fig.4 Nucleotide sequence and deduced amino acid sequence of gene RsgidA
3.4.2 RsGidA蛋白的理化性質及結構預測 利用ProtParam軟件對RsgidA基因的編碼蛋白進行理化性質預測。RsGidA蛋白相對分子質量為69 071.73,理論等電點為6.21,分子式為C3038H4863N867O923S24,脂肪系數為86.04,不穩定指數為38.76,親水性平均系數為?0.319。利用ProtScale軟件對RsGidA蛋白進行親疏水性預測,由圖5可看出,該蛋白親水性氨基酸數量多于疏水性氨基酸,預測為親水性蛋白。利用SignalP 5.0及TMHMM軟件分別對RsGidA蛋白進行信號肽及跨膜結果預測,結果顯示該蛋白不含信號肽,無跨膜區。利用SOPMA軟件進行蛋白質的二級結構預測,結果顯示,RsGidA蛋白二級結構中包含256個α螺旋,216個無規卷曲,98個延伸鏈,56個β-轉角,分別占氨基酸總數的40.89%、34.5%、15.65%和8.95%。以大腸桿菌GidA的晶體結構(PDB ID:3ces)為模板,利用SWISS-MODEL軟件對RsGidA蛋白的三級結構進行預測,其空間結構主要由α螺旋和無規則卷曲組成,與二級結構預測結果相符合(圖6)。
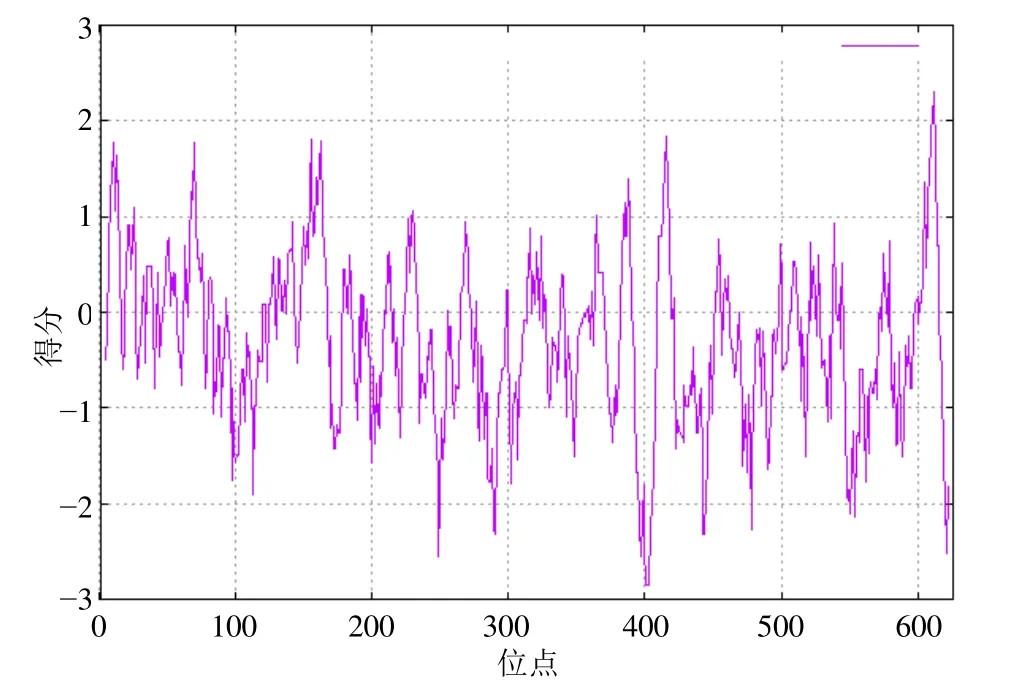
圖5 RsGidA蛋白的疏水性分析Fig.5 Hydrophobicity analysis of RsGidA protein
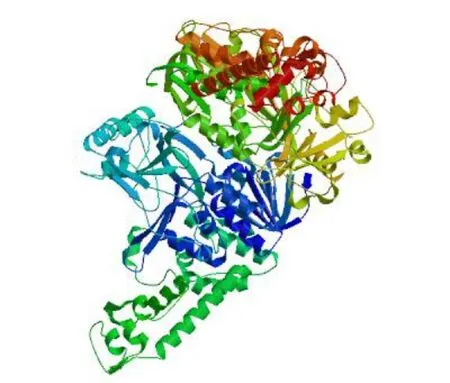
圖6 RsGidA蛋白的三級結構預測Fig.6 Prediction of tertiary structure of RsGidA protein
3.4.3 RsGidA蛋白與不同植物病原細菌同源氨基酸的系統進化分析 一般認為氨基酸序列相似性大于30%的蛋白質可能由同一種蛋白分化而來,為同源蛋白,可能具有相似的結構和功能[8]。RsGidA蛋白與不同植物病原菌中的GidA氨基酸序列進行比對及系統進化分析,blastp結果顯示,RsGidA與丁香假單胞菌Pseudomonas syringae、菠蘿泛菌Pantoea ananatis、生癌腸桿菌Enterobacter cancerogenus、西瓜食酸菌Acidovorax citrulli及野油菜黃單胞菌Xanthomonas campestris的同源氨基酸序列相似性分別為71.36%、66.77%、66.03%、64.45%及64.68%。采用MEGA 5.1軟件的鄰接法構建系統發育樹,表明RsGidA蛋白與其他植物病原細菌同源蛋白的關系與blastp比對結果一致(圖7),其中,RsGidA蛋白與丁香假單胞菌的GidA蛋白親緣關系最近。
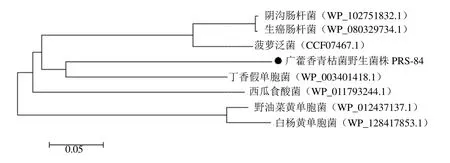
圖7 RsGidA蛋白與不同植物病原細菌同源氨基酸的系統進化分析Fig.7 Phylogenetic tree analysis of RsGidA protein and homologous proteins from different plant pathogen strains
4 討論
GidA蛋白又名MnmG[9](tRNA uridine 5-carboxymethylaminomethyl modification enzyme MnmG),在原核及真核細胞中廣泛存在且高度保守[10-11],其介導的tRNA修飾對于蛋白質的準確翻譯具有重要作用[12],該蛋白的缺失或者突變會引起基因表達功能紊亂,導致細胞正常生命活動異常。von Meyenburg等[13]對大腸埃希菌進行了Tn10轉座子插入突變,獲得了在含有葡萄糖培養基中細胞形態改變的突變株,研究表明該突變株的轉座子插入了1個功能基因,該基因在葡萄糖的作用下可以使細胞的分裂受到抑制(glucose-inhibited division),因此將該基因命名為gidA。Shippy等[14-15]對沙門氏菌的gidA基因進行缺失突變后,細胞形態呈絲狀改變,還發現突變株的染色體分裂受阻,與細胞分裂相關的基因表達異常。
最近的研究表明,gidA基因還與病原菌的致病性相關。Kinscherf等[16]通過Tn5轉座子插入突變獲得了丁香假單胞菌Pseudomonas syringae的毒性突變菌株,該突變株的脂肽毒素分泌能力及運動性消失,毒性減弱,研究發現突變株的轉座子插入位點位于gidA基因的開放閱讀框內,表明gidA基因對丁香假單胞菌的毒性具有調控作用。Gupta等[17]構建了銅綠假單胞菌Pseudomonas aeruginosa的轉座子插入突變體庫,并篩選得到多個減毒株,其中gidA基因的失活對LasA蛋白酶和綠膿菌素等毒力因子的合成具有顯著影響。此外,張偉等[18]以生防菌熒光假單胞菌Pseudomonas fluorescens為研究對象,通過Tn5轉座子插入突變的方法獲得gidA基因突變的菌株,該突變株生物膜形成能力及對小麥根部的定殖能力均較野生菌株顯著降低。
在細菌分類鑒定中,假單胞菌屬Pseudomonas曾是一個大屬,勞爾氏菌屬細菌Ralstonia曾隸屬于假單胞菌屬,而青枯勞爾氏菌Ralstonia solanacearum(簡稱青枯菌)最早也被命名為青枯假單胞菌Pseudomonas solanacearum,表明青枯菌與假單胞菌有較大的關聯。本研究成功從廣藿香青枯菌菌株R.solanacearumPRS-84中克隆了RsgidA基因,系統進化分析表明,該基因與丁香假單胞菌P.syringae的gidA基因具有較近的親緣關系。
有研究表明,細菌的運動性和生物膜形成能力與病原菌的致病力相關。病原菌的運動性在其侵染寄主植物的過程中具有重要作用[19-20],運動性功能缺陷可導致病原菌致病性的減弱[21]。植物病原細菌的生物膜的形成有利于病原菌在寄主植物中定殖,一些植物病原細菌的生物膜可在寄主植物的木質部中形成,導致導管堵塞,從而使植株出現萎蔫失水等癥狀[22-23]。本實驗室前期對低致病力突變株PRS-84-4-7的運動性及生物膜形成能力進行了研究,結果顯示,與野生菌株PRS-84相比,突變株PRS-84-4-7的運動性降低,生物膜形成減少(未發表數據),推測RsgidA基因可能通過調控青枯菌的運動性和生物膜形成,從而在青枯菌侵染廣藿香及其在體內定殖過程中發揮作用。
利益沖突所有作者均聲明不存在利益沖突
