豬源蓋塔病毒Cap蛋白與E2蛋白多克隆抗體的制備及其免疫特性分析
2021-05-28 04:14:12蘇靖茵燕詩雨張萌粟碩
畜牧與獸醫 2021年6期
關鍵詞:血清
蘇靖茵,燕詩雨,張萌,粟碩
(南京農業大學免疫研究所,江蘇 南京 210095)
蓋塔病毒(Getah virus, GETV)是典型的蟲媒病毒,最早于1955年從馬來西亞的白雪庫蚊中分離并命名,隨后陸續又從豬群、馬群中分離出該病毒。近年來,世界各國出現母豬感染零星病例,而在2017年中國湖南豬場出現母豬和仔豬蓋塔病疫情[1]。臨床上,GETV對豬和馬的侵害性較大,他們的癥狀主要表現為發熱、浮腫等[2]。GETV在國內外均有流行,且我國陸續報道除馬、豬以外的多種動物(牛和狐貍等)感染病例[3]。
GETV是單股正鏈RNA病毒,屬于披膜病毒科甲病毒屬的成員。GETV全基因組長11~12 kb,包含2個開放閱讀框,分別編碼非結構蛋白和結構蛋白。GETV的5個結構蛋白分別是C、E3、E2、6K和E1。在甲病毒中,Cap蛋白是一種多功能蛋白,包含N端RNA結合域和C端結構域[4]。Cap蛋白能與病毒基因組特性結合,這會影響病毒核衣殼形成和RNA合成[5]。此外,Cap蛋白不僅能促進核衣殼組裝過程中Cap蛋白二聚作用,還能促進在出芽過程中與糖蛋白細胞質結構域的相互作用[6]。E2是糖蛋白,分子量約為50 kDa;pE2是E2的前體,能裂解成E2和E3[5]。在甲病毒粒子感染過程中,E2作為介導病毒進入宿主細胞的主要蛋白。
GETV的Cap蛋白與E2蛋白可能在病毒感染和病毒復制中起重要重要作用。但現今對于GETV的病毒蛋白與及致病機制等研究仍較少[7],而GETV的抗體能促進其相關研究。因此,本文通過了原核表達Cap和E2蛋白并制備了GETV的多抗,以用于GETV的基礎研究。
1 材料與方法
1.1 材料
Vero、BHK細胞和pET-32a(+)載體均為本實驗室保存。DH5α與BL21(DE3)感受態均購自康為公司。GETV由本實驗室分離純化后保存。6~8周齡雌性BALB/c小鼠,購自上海西普爾-必凱實驗動物公司。
限制性內切酶購自NEB公司;DNA Marker購自TaKaRa公司;胎牛血清、DMEM培養基、標準蛋白Marker均購自賽默飛公司;高保真DNA聚合酶、同源重組連接試劑盒,化學發光試劑(ECL)顯色液均購自諾唯贊公司;辣根過氧化物酶(HRP)標記的山羊抗鼠抗體、HRP標記的山羊抗兔抗體、異硫氰酸熒光素(FITC)標記的山羊抗鼠的抗體均購自KPL公司;His兔源抗體購自ABclonal公司。
1.2 方法
1.2.1 引物設計
以GenBank中登錄的GETV毒株NC_006558.1為參考毒株,設計E2基因糖蛋白區域和Cap基因編碼區特異性引物。引物由南京金斯瑞公司合成。
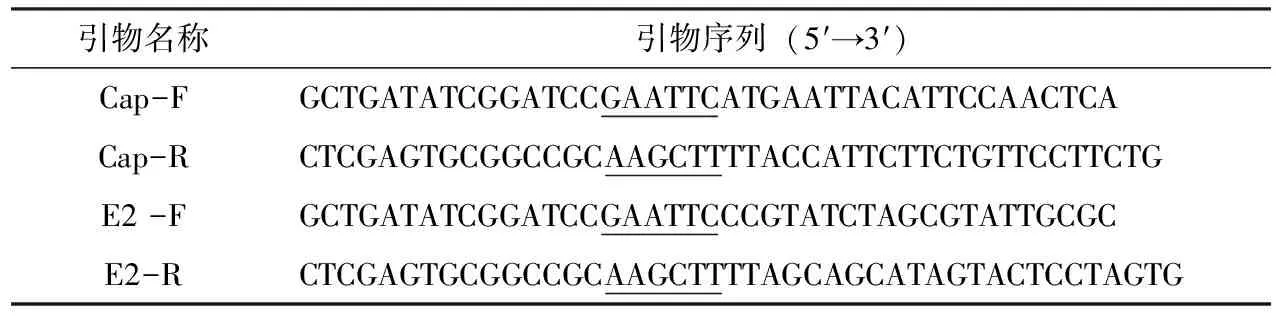
表1 GETV-Cap基因和GETV-E2基因擴增引物
1.2.2 目的基因的擴增
Vero細胞長至80%~90%時,接種GETV,細胞出現病變后,加入TRIzol裂解細胞。按照TRIzol試劑說明書,提取細胞培養物中病毒的RNA。使用RT-PCR方法擴增Cap基因及E2基因,反應體系為50 μL,反應條件為:94 ℃,4 min;98 ℃ 10 s,58 ℃ 30 s,72 ℃ 1 min,38 個循環;72 ℃ 10 min。PCR產物凝膠電泳后,回收目的片段。
針對水產品運輸環節監管,2017年,佛山市建立鮮活水產品跨區域產銷對接監管協作機制,與長沙市食安辦、食藥監、農業等部門聯合簽署《加強區域間鮮活水產品產銷對接監管合作框架協議》。建立水產品質量安全聯盟,以何氏水產為代表的10多家大型水產品經營企業共同建立了統一的“收販運”質量標準體系,有效解決水產品運輸環節非法添加問題。
1.2.3 原核表達質粒的構建
空載體pET-32a(+)經EcoRⅠ與HindⅢ雙酶切后回收載體。使用同源重組酶將回收目的片段與回收后的載體連接。連接產物轉DH5α,并涂布于含氨芐抗性的平板,37 ℃培養12~16 h。挑取單菌落,37 ℃擴培后,提質粒雙酶切(EcoRⅠ與HindⅢ)鑒定。根據酶切鑒定的結果,將陽性克隆送公司測序,鑒定正確后獲得質粒pET-Cap和pET-E2。
1.2.4 Cap與E2原核表達
將空載體pET-32a(+)、pET-Cap和pET-E2質粒分別轉入BL21(DE3)感受態中,挑取單菌落獲得pET-32a(+)、pET-Cap和pET-E2表達菌種。將pET-Cap 和pET-E2 菌種接種至含氨芐抗性的LB培養基中,37 ℃培養至菌液OD600值為0.6~0.8。摸索誘導條件分別為:誘導溫度為37、25 和16 ℃;異丙基-β-D-硫代半乳糖苷(IPTG)濃度為0.5、1.0 和1.5 mmol/L;誘導時間為3、5和7 h。選取較優誘導條件,誘導后取菌體超聲約30 min至菌體清亮。12 000 r/min離心,分離上清與沉淀。將沉淀溶于包涵體溶解液中,4 ℃過夜,具體操作參考文獻[8]。SDS-PAGE分析pET-Cap和pET-E2原核表達蛋白主要分布于包涵體中。把原核表達蛋白經鎳柱親和層析純化,通過SDS-PAGE分析純化效果。純化后的蛋白使用脲素梯度稀釋法進行透析復性,4 ℃,12 h/次,前后經過復性液1~4,具體操作參考文獻[9-10]。將復性蛋白和誘導空載體分別制備成蛋白樣品,SDS-PAGE分析后,半干轉至硝酸纖維素膜(NC膜);5% 脫脂奶封閉后,孵育His抗體和HRP標記的山羊抗兔抗體,ECL發光顯色。
1.2.5 小鼠多抗血清制備
復性的Cap和E2蛋白分別作為免疫原,經皮下背部多點方式,100 μg/只,免疫6~8周齡雌性BALB/c小鼠。首免免疫原和等體積完全弗氏佐劑乳化,而二免和三免用不完全弗氏佐劑替代完全弗氏佐劑。每次免疫間隔為2周,三免后1周采血,分離免疫小鼠血清作為陽性血清,不免疫小鼠血清作為陰性血清。ELISA測定血清效價,分別測定梯度稀釋后的陽性血清效價,操作參照文獻[11],酶標儀讀取各孔的OD值。
1.2.6 Westeron blot鑒定
BHK細胞以感染復數(MOI) 0.01接種GETV,細胞病變后48 h,制備接種與不接種GETV的蛋白樣品,細胞裂解后收集樣品,加入4×上樣緩沖液后煮樣。經SDS-PAGE分析后轉至NC膜;封閉NC膜后,孵育Cap和E2 陽性血清一抗和HRP標記的山羊抗鼠二抗;洗膜后,ECL發光顯色。
1.2.7 免疫熒光試驗(IFA)鑒定
BHK細胞長至80%以上時,以MOI為0.01接種GETV,48 h后使用冰甲醇固定細胞。封閉后,分別與 E2、Cap血清和PBS(對照組)作用;接著以FITC標記的山羊抗鼠的抗體為二抗孵育,并使用4′,6-二脒基-2-苯基吲哚(DAPI)染核,最后倒置顯微鏡下觀察拍照。
2 結果與分析
2.1 PCR目的條帶擴增與質粒構建
以GETV的cDNA為模板,通過Cap特異性引物,RT-PCR方法擴增出約800 bp的片段(圖1A),與GETV的Cap基因理論值(804 bp)相符;通過E2特異性引物,用RT-PCR方法擴增出約1 200 bp的片段(圖1B),與GETV的E2基因理論值(1 209 bp)大小一致。
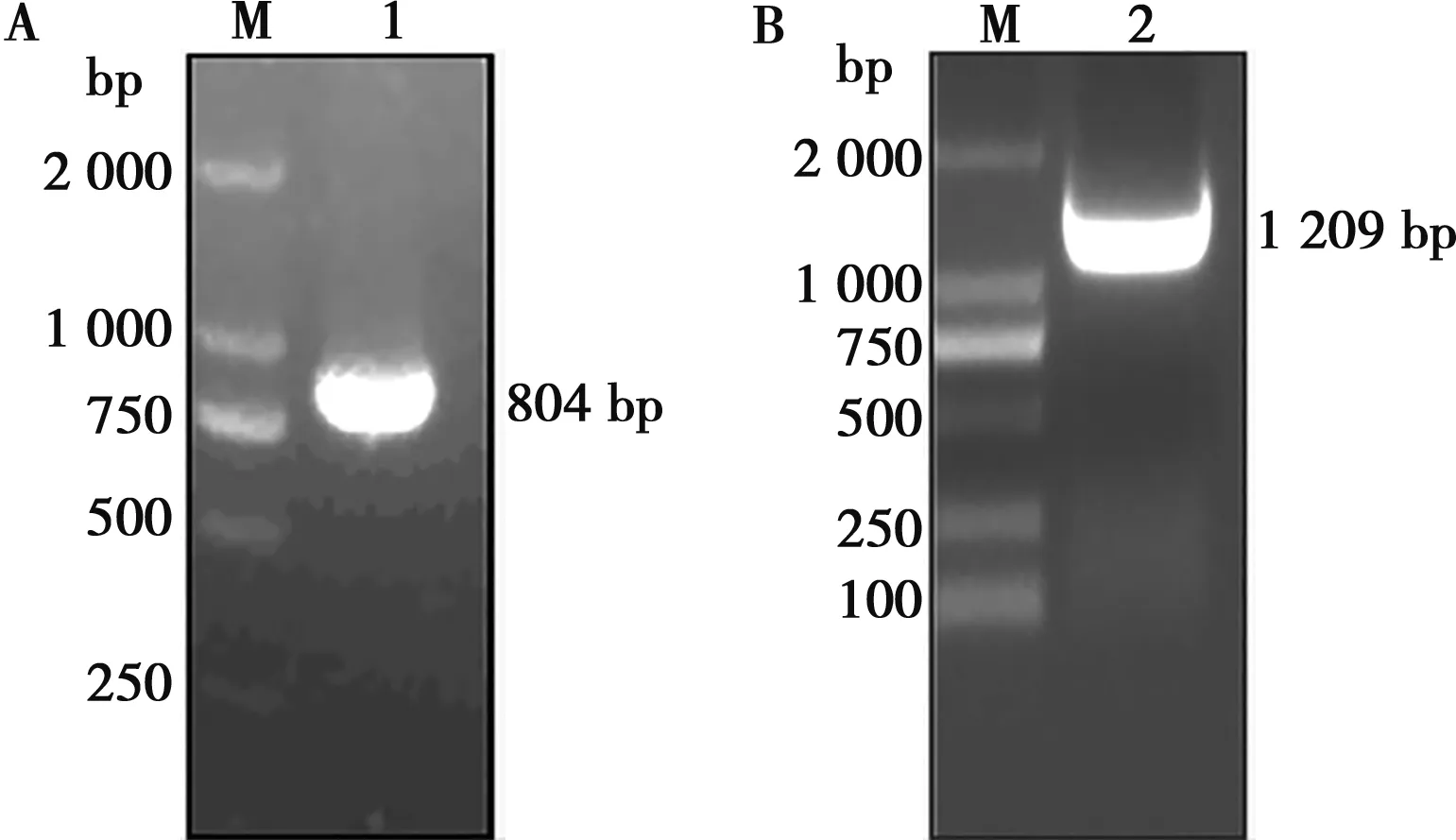
M. DNA Marker DL2000;1. Cap基因;2. E2基因
2.2 重組質粒的構建與鑒定
同源重組酶將目的片段與雙酶切(EcoRⅠ和HindⅢ)的pET-32a(+)連接,構建成重組克隆質粒。分別使用Cap和E2的特異性引物,以Cap陽性質粒和E2陽性質粒為模板,分別擴增出804 bp與1 209 bp的單一條帶。質粒酶切鑒定結果顯示,空載體酶切后可見大小為5 900 bp條帶;Cap陽性質粒有5 900和804 bp 2條帶;E2陽性質粒有2條分別在5 900和1 209 bp附近的條帶,均符合預期(圖2)。將陽性質粒送公司測序,測序結果與原始序列比對一致,分別命名為pET-Cap和pET-E2。
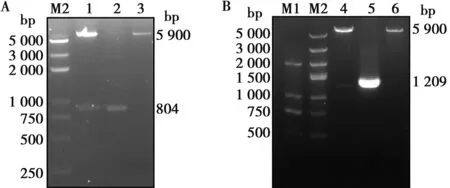
M1. DNA Marker DL2000;M2. DNA Marker DL5000;1.雙酶切pET-Cap質粒;2.Cap基因擴增;3.雙酶切空載體;4.雙酶切pET-E2質粒;5.E2基因擴增;6.雙酶切空載體
2.3 重組蛋白的表達與純化
通過比較不同條件下誘導pET-Cap情況,最終選擇誘導條件為:IPTG濃度為1 mmol/L,37 ℃,誘導3 h。誘導后包涵體中40~55 kDa附近有明顯條帶,而上清僅有少量蛋白,表明Cap蛋白主要在包涵體中大量表達(圖3A)。包涵體中的蛋白經鎳柱親和層析純化后獲得重組Cap蛋白(圖3B)。純化復性后Cap蛋白樣品與His抗體結合,在40~55 kDa附近有條帶,純化復性蛋白為重組Cap蛋白(圖3C)。
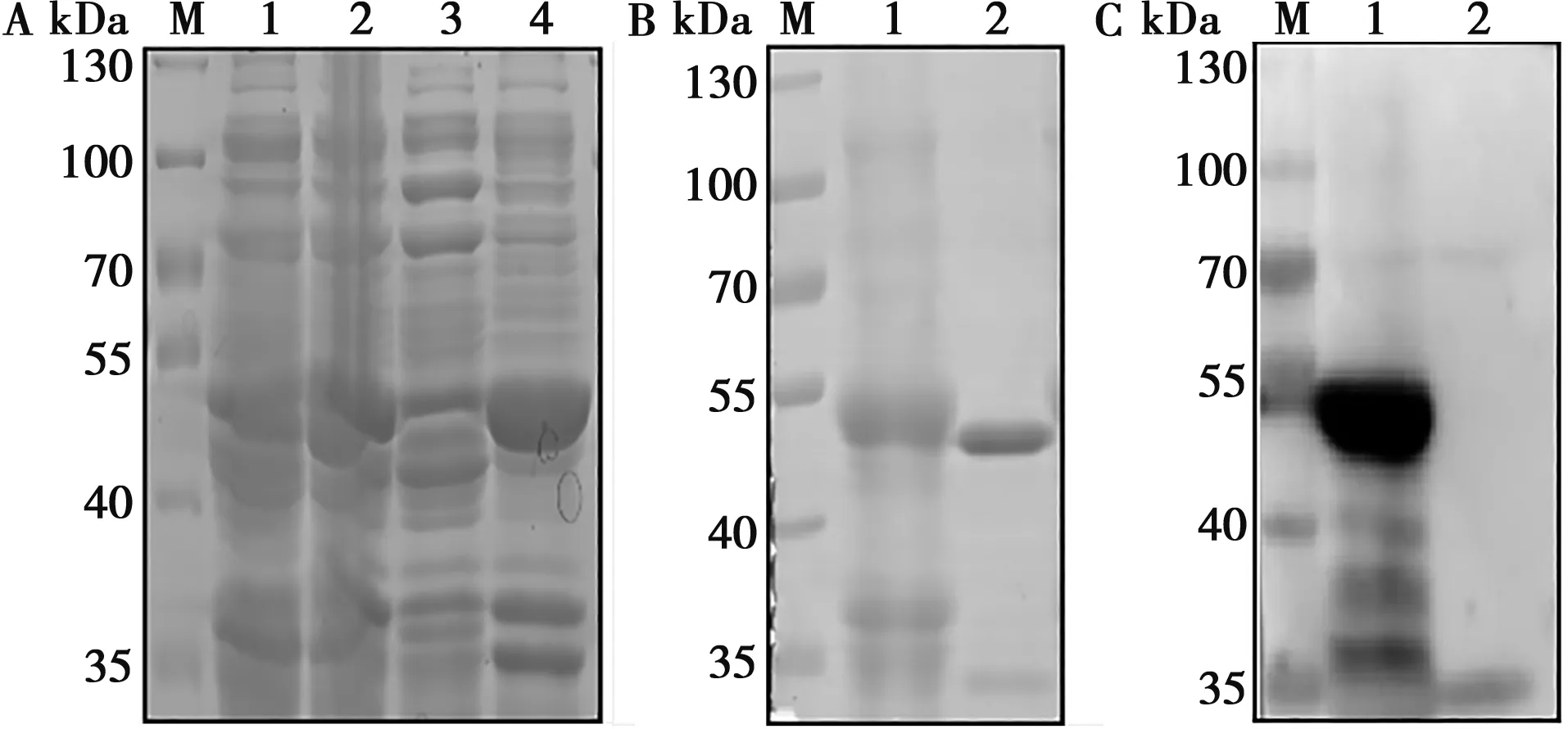
A.pET-Cap表達:M.標準蛋白Marker;1. pET-Cap未誘導;2.pET-Cap誘導;3.pET-Cap超聲后上清;4.pET-Cap超聲后沉淀;B.Cap蛋白純化:M.標準蛋白Marker;1.純化pET-Cap沉淀;2.復性Cap蛋白;C.Cap蛋白與His抗體反應:M.標準蛋白Marker;1.復性Cap蛋白;2.pET-32a(+)誘導
通過分析比較不同溫度、時間與不同濃度的IPTG誘導pET-E2的效果,IPTG濃度為1 mmol/L時,在25 ℃,誘導5 h下E2蛋白大量表達,55~70 kDa附近有蛋白帶,且蛋白主要存在于包涵體中(圖4A)。經鎳柱親和層析純化和復性透析包涵體中蛋白后,獲得重組E2蛋白(圖4B)。純化復性后的E2蛋白與His抗體反應,在55 kDa處有明顯條帶而空載體沒有,表明純化復性的蛋白為重組E2蛋白(圖4C)。
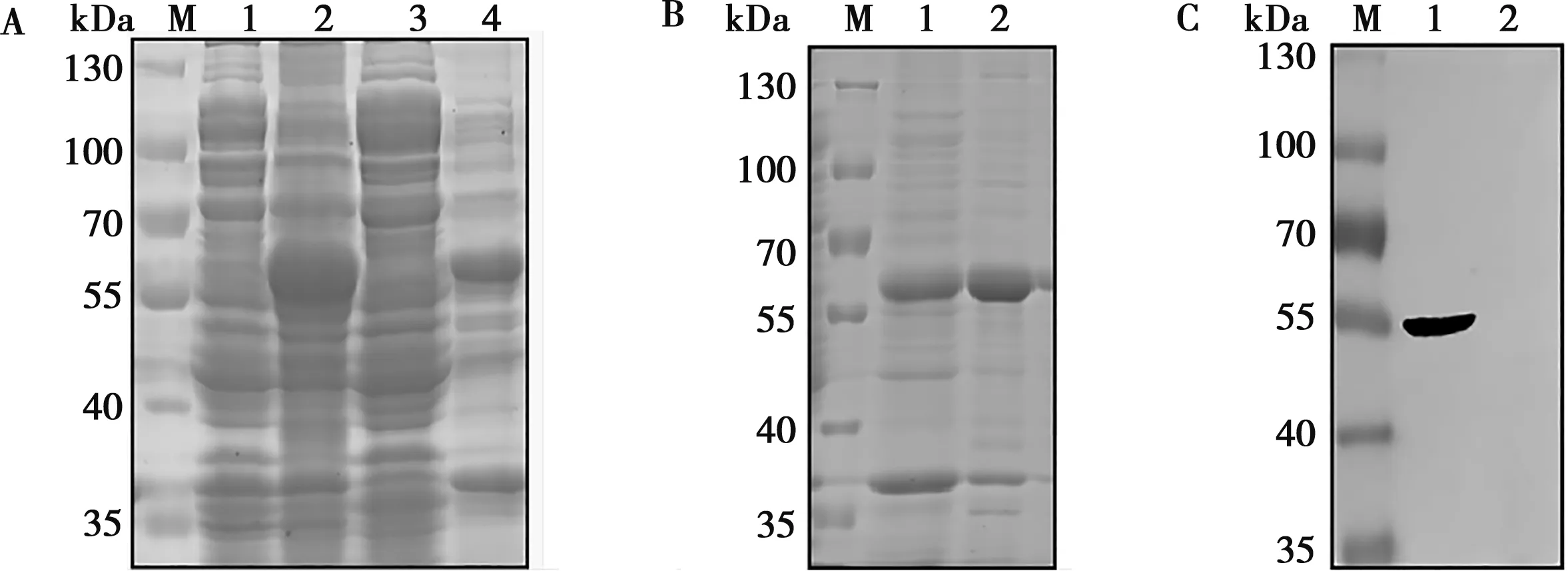
A.pET-E2表達:1. pET-E2未誘導;2.pET-E2誘導;3.pET-E2超聲后上清;4.pET-E2超聲后沉淀;M.標準蛋白Marker;B. E2蛋白的純化:1. 純化pET-E2沉淀;2.復性E2蛋白;C. E2蛋白與His抗體反應:1.復性E2蛋白;2.pET-32a(+)誘導
2.4 Cap蛋白與E2蛋白多克隆抗體的制備
Cap與E2原核表達蛋白分別免疫小鼠制備多克隆抗體,經3次免疫后ELISA檢測小鼠血清抗體效價,結果2種抗體效價均達到1∶105以上。
2.5 Western blot 驗證多抗效果
GETV的Cap蛋白理論大小約為30 kDa,BHK(圖5A)和Vero(圖5C)細胞接種病毒后不同時間點的樣品在25~35 kDa附近可見有明顯條帶,而不接種病毒樣品未見條帶,說明Cap多抗能與GETV發生特異性結合。
GETV的E2蛋白理論大小為46 kDa,其前體pE2為62 kDa。BHK(圖6B)和Vero(圖6C)細胞接種病毒后不同時間點的樣品在40~55 kDa與55~70 kDa附近均可見明顯條帶,而不接種病毒樣品未見條帶,說明E2多抗與GETV的反應性良好。
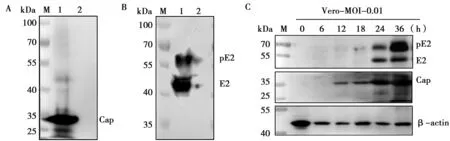
M.蛋白標準Marker;1.BHK細胞接種GETV;2.BHK細胞不接種病毒
2.6 IFA驗證多抗效果
GETV接種于BHK細胞,以Cap多抗和E2多抗為一抗進行IFA,均可檢測到綠色熒光,而對照組沒有綠色熒光,說明Cap多抗和E2多抗能特異性識別GETV(圖6)。
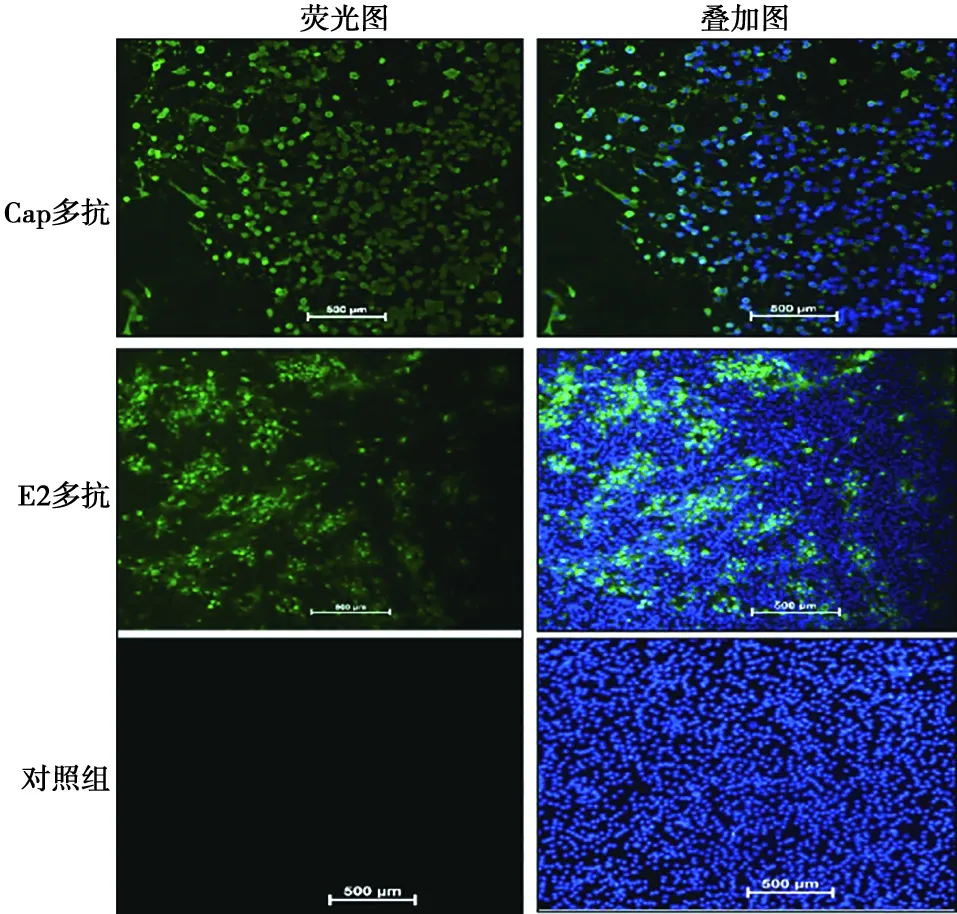
圖6 Cap和E2多抗與GETV反應性的IFA鑒定(標尺=500 μm)
3 討論
GETV在我國廣西、四川、云南、海南、河北、湖南和甘肅等多地均有報道,表明該病毒在我國普遍流行[12]。在自然界中,GETV是一種蟲媒病毒,帶毒蚊蟲通過叮咬豬、馬、鼠等易感動物,使病毒在這些動物體內增殖[13]。Li等[14]對家畜血清樣本進行調查后,發現在雞、鴨、奶牛、豬和肉牛中均能檢測GETV的中和抗體,其中豬和肉牛抗體陽性率較高(46%~72%)。2010年在河北省涉縣出現正常人群和發燒患者感染GETV情況,流行病學篩查后發現正常人群的總感染率高達16.67%,遠高于海南瓊中地區的陽性率(約2.0%)[15]。這說明GETV可能具有重要的公共衛生學意義。
近年來對甲病毒的功能報道較多,而GETV的報道相對較少。研究發現甲病毒囊膜蛋白E2存在糖基化位點、宿主細胞受體識別位點和中和抗體的識別位點[5]。甲病毒科辛德比斯病毒(Sindbis virus)E2蛋白單抗能夠影響病毒在體內和體外的復制[16]。E2單抗處理后感染病毒,盡管細胞內核衣殼蛋白和囊膜蛋白仍持續合成,但病毒粒子釋放減少[17]。甲病毒囊膜糖蛋白E2蛋白C端接觸的是由Cap蛋白和基因組RNA組成的病毒核衣殼[6]。一旦核衣殼蛋白從初生的多肽鏈中釋放出來,N端信號序列將發揮作用,導致糖蛋白pE2插入內質網[5]。甲病毒的Cap蛋白N端結構域雖保守性較低,富含正電荷。Cap蛋白存在核定位信號和核出口信號位點,在核胞質運輸中發揮作用[18]。而Cap蛋白的C端蛋白酶結構域高度保守,Cap蛋白的114~264殘基表面存在疏水區域,并被譽為疏水口袋。該口袋能與E2蛋白C端的16個殘基相互作用[19],該結合在甲病毒組裝中發揮積極作用。
目前,國內外對于GETV的E2蛋白研究主要是基于原核表達E2蛋白并建立ELISA方法[20- 21],而對于其抗體制備報道較少。E2與Cap蛋白抗體對研究GETV蛋白的功能起著重要的作用。姜焱等[22]成功制備了GETV的Cap蛋白多抗,但沒有進一步驗證Cap多抗與GETV的反應性。GETV的Cap蛋白與E2蛋白多抗不僅能應用于分離鑒定GETV,還能應用于流行病學調查。為深入了解GETV,本文制備了其鼠源Cap多抗與E2多抗,Western blot和IFA均證實此2種多抗均能特異性識別GETV,為進一步探究GETV致病機制提供了物質基礎。
猜你喜歡
中老年保健(2021年3期)2021-08-22 06:50:04
天津醫科大學學報(2021年2期)2021-03-29 05:31:08
昆明醫科大學學報(2021年1期)2021-02-07 01:06:36
現代臨床醫學(2021年1期)2021-01-26 00:56:02
昆明醫科大學學報(2020年12期)2021-01-26 00:44:04
中華養生保健(2020年4期)2020-11-16 01:31:40
中西醫結合肝病雜志(2020年2期)2020-10-27 02:18:50
豬業科學(2018年8期)2018-09-28 01:27:38
中成藥(2017年8期)2017-11-22 03:18:47
川北醫學院學報(2015年5期)2015-12-05 08:22:29
