
E.coli Nissle 1917(EcN)curli菌毛和鞭毛對其生物被膜形成的影響
2021-06-15 04:17:37龔冰雪周賢軒
安徽農業科學 2021年10期
龔冰雪 周賢軒

摘要 E.coli Nissle 1917 是被廣泛認可的益生菌,可以通過形成生物被膜定殖在腸道中抑制沙門氏菌等病原菌的生長。通過λ-Red同源重組系統構建curli菌毛合成基因(csgA)和鞭毛調控基因(hnsA)突變菌株,探究curli菌毛和鞭毛對EcN生物被膜形成的影響。結果表明,curli菌毛對EcN運動能力以及生物膜形成沒有影響;EcNΔhnsA菌株的運動能力下降,生物被膜含量升高,說明鞭毛通過促進EcN運動,抑制細菌的附著,從而抑制生物膜的形成。
關鍵詞 E.coli Nissle 1917(EcN);curli菌毛;鞭毛;生物被膜;形成;影響
中圖分類號 Q939.9? 文獻標識碼 A? 文章編號 0517-6611(2021)10-0008-04
doi:10.3969/j.issn.0517-6611.2021.10.003
開放科學(資源服務)標識碼(OSID):
Effects of Curli and Histone-like Nucleoid Structuring Protein on Biofilm Formation of E.coli Nissle 1917 (EcN)
GONG Bing-xue,ZHOU Xian-xuan
(School of Food Science and Engineering, Hefei University of Technology, Hefei,Anhui 230009)
Abstract E.coli Nissle 1917 is a widely recognized probiotics, which can inhibit the growth of Salmonella and other pathogenic bacteria in the intestinal tract by forming biofilm colonization. The mutants of curli fimbriae synthesis gene (csgA) and flagellum regulatory gene (hnsA) were constructed by λ-Red homologous recombination system, and the effects of curli fimbriae and flagellum on the biofilm formation of EcN were investigated. The results showed that curli fimbriae had no effect on the ability of motility and biofilm formation of EcN. While the ability of motility of EcNΔhnsA strain decreased and biofilm formation increased, indicating that flagellum inhibited the formation of biofilm by promoting the motility ability and inhibiting the attachment of EcN.
Key words E.coli Nissle 1917(EcN);Curli fimbriae;Flagellum;Biofilm;Formation;Effect
生物被膜是嵌入在生物或非生物表面上的自生三維結構基質中的微生物細胞集合[1],其生長發育一般由初始黏附、固定、成熟和分散4個階段組成[2]。細菌生物被膜的形成在病原微生物持續性感染中起著非常重要的作用,是許多臨床疾病慢性反復感染的根源[3]。
大腸桿菌Nissle 1917(EcN)是被普遍認為的腸道益生菌,可以通過形成生物膜在腸道中定殖,與病原菌競爭腸道附著空間,以此來抑制腸道致病菌繁殖[4-5]。因此EcN通常被用于預防與治療傳染性腹瀉和炎性腸疾病等。
curli菌毛是由革蘭氏陰性菌產生、位于細胞表面的蛋白質聚集體。在大腸桿菌和腸道沙門氏菌的生物被膜中,curli是細胞外基質的主要蛋白質組分。在生物膜內,curli纖維形成一個相互交織的網狀結構,支持細胞外基質并包裹住單個細胞。研究表明,curli菌毛通過促進細菌黏附來促進生物被膜的形成[6-7]。類組蛋白類核結構蛋白(H-NS)是大腸桿菌中高度保守的蛋白[8]。研究表明,H-NS通過結合在鞭毛主操縱子flhDC啟動子上影響其表達,hns突變會導致編碼鞭毛蛋白的結構基因fliC和編碼鞭毛sigma因子基因fliA的表達水平降低,導致細菌運動能力喪失[9-10],從而影響其生物被膜的形成[11]。
然而,curli菌毛和鞭毛對EcN生物膜形成的影響尚不清楚。筆者通過λ-Red同源重組系統構建curli 菌毛合成基因(csgA)和鞭毛調控基因(hnsA)突變菌株,探究curli菌毛和鞭毛對EcN生物被膜形成的影響。
1 材料與方法
1.1 試驗材料
大腸桿菌菌株Nissle 1917、BW25142,質粒pKD3、pKD46、pCP20,均由合肥工業大學微生物與酶工程實驗室保存。Primer Star DNA聚合酶、DNA分子量標準、乙酸、結晶紫,均購于生工生物工程(上海)股份有限公司;引物由通用生物系統(安徽)有限公司合成;大腸桿菌培養所用的培養基為LB和M9C(6.8 g/L Na2HPO4、3.0 g/L KH2PO4、0.5 g/L NaCl、1.0 g/L NH4Cl、0.4%葡萄糖、0.4%酪蛋白氨基酸、2 mmol/L MgSO4、0.1 mmol/L CaCl2);培養所需的抗生素為氨芐青霉素(Ap,100 μg/mL)和氯霉素(Cm,15 μg/mL)。
1.2 試驗方法
1.2.1 EcN中的基因敲除。
采用λ-Red同源重組系統對EcN進行基因敲除[12]。首先以質粒pKD3為模板,用兩端帶有目的基因上下游的39 bp同源臂的引物對PCR擴增FRT-Cm-FRT片段,將得到的PCR產物進行純化,電擊轉化入EcN/pKD46電擊感受態細胞中,涂于Cm抗性平板,37 ℃培養。挑選隔夜生長的單克隆菌落作為模板,加入相應檢驗引物進行菌落PCR檢測。
1.2.2 抗性的去除。
菌落PCR鑒定結果為陽性的菌株可利用輔助質粒pCP20將抗性片段去除,pCP20電轉至陽性菌落,涂布于Ap抗性平板,30 ℃培養過夜,挑取單克隆于LB培養基(Ap,100 μg/mL),30 ℃培養。蘸取菌液劃線于LB平板,42 ℃培養使溫敏性質粒pCP20丟失。將單菌落分別依次劃線于Ap抗性平板、Cm抗性平板、LB平板,37 ℃培養。對Ap平板和Cm平板上都不生長、LB平板上生長的菌落進行PCR檢驗。
1.2.3 生長曲線的測定。
用M9C培養基將隔夜培養的菌液稀釋至OD600為0.03,于37 ℃振蕩培養,每隔1 h取樣測OD600,直至細菌生長至穩定期。
1.2.4 生物被膜的形成。
用M9C培養基將隔夜培養的菌液稀釋至OD600為0.05,加入聚苯乙烯96微孔板,每孔200 μL,于37 ℃恒溫培養箱中靜置培養24 h,吸出菌液,置于50 ℃培養箱中烘干20 min。每孔加入250 μL 0.1%結晶紫染色液染色15 min。吸去染色液,去離子水洗滌3次以去除未結合的結晶紫,置于50 ℃培養箱中烘干20 min。每孔加入200 μL 30%乙酸溶液,洗凈孔壁及孔底,吸出脫色液,測OD570。
1.2.5 泳動試驗。
用M9C培養基將隔夜培養的菌液稀釋至OD600為0.1,取2 μL滴于0.3%瓊脂糖半固體培養基上,37 ℃恒溫培養24 h,保持空氣濕潤。拍照記錄并測量泳動圈的直徑。
2 結果與分析
2.1 EcN中csgA及hnsA的敲除
以pKD3為模板擴增FRT-Cm-FRT片段,電泳結果如圖1所示,泳道1、2是帶有csgA基因同源臂的FRT-Cm-FRT片段,泳道3、4是帶有hnsA基因同源臂的FRT-Cm-FRT片段。條帶大小約為1 kb,符合預期大小。將純化后的DNA片段電轉至EcN/pKD46中,在pKD46表達的λ噬菌體重組酶的作用下其同源臂序列可與基因組上的同源序列發生同源重組。將轉化的菌液涂在相對應的抗性平板上,隔夜培養后挑取單克隆進行菌落PCR檢測。
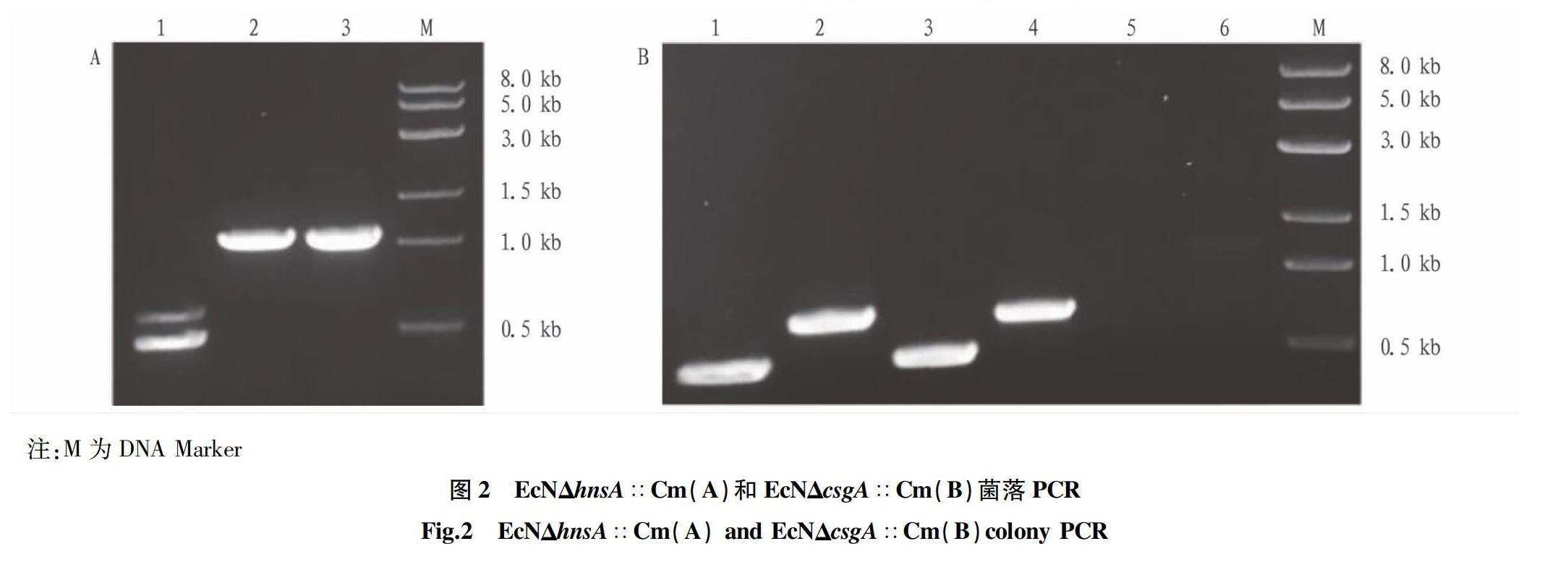
hnsA的敲除結果如圖2A,選取2個單克隆和陰性對照
EcN/pKD46,同一個模板分別加入2對檢驗引物,泳道1、2為①號菌,泳道3、4為②號菌,泳道5、6為陰性對照EcN/pKD46,陽性克隆的預期條帶大小為0.49和0.71 kb,①號菌與②號菌均為陽性克隆EcNΔhnsA∷Cm。
csgA的敲除結果如圖2B,泳道1、2為隨機挑選的2個單克隆,加入4條檢驗引物進行菌落PCR鑒定,泳道3為陰性對照EcN/pKD46,陽性克隆目的片段大小應為0.47和0.61 kb,由圖可知泳道1電泳結果符合預期,該單克隆為陽性克隆EcNΔcsgA∷Cm。
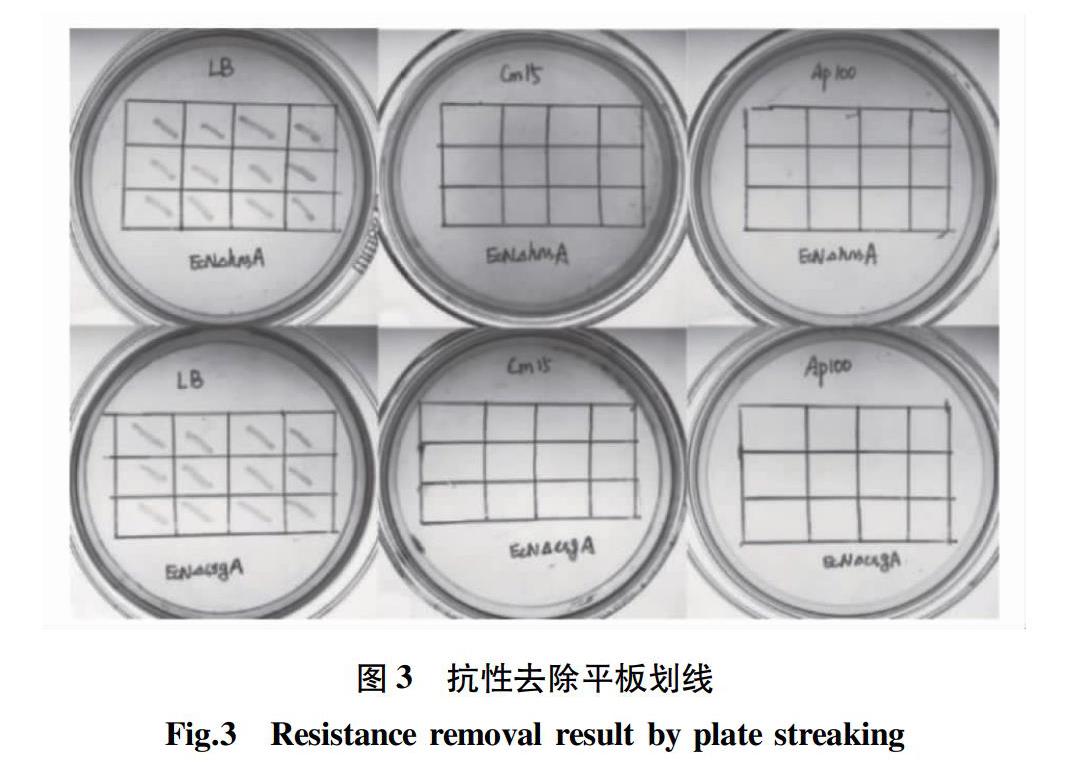
2.2 抗性的去除
將EcNΔcsgA∷Cm/pCP20和EcNΔhnsA∷Cm/pCP20劃線于LB板42 ℃培養過夜。挑取單克隆,依次劃線LB板、Cm(15 μg/mL)板和Ap(100 μg/mL)板,在37 ℃培養,結果如圖3所示,所有的單克隆均在LB平板上能夠生長,抗性平板上不生長,說明菌株的氯霉素抗性片段去除成功,且質粒pCP20也已丟失。
為保證細菌的抗性去除成功,通過菌落PCR的方法再次檢驗,各隨機挑取2個單克隆作為模板,加入相應引物進行菌落PCR檢測。成功去除抗性的菌株條帶大小分別為0.75 kb(csgA)、0.87 kb(hnsA)。如圖4,條帶與預期相符,說明菌株的抗性片段被成功去除。
2.3 EcN野生型及突變株生長曲線
基因敲除成功后,對突變株EcN△csgA、EcN△hnsA與野生型EcN的生長情況進行測定,進而來評估該基因是否影響細菌的生長。該研究選用M9C培養基來測定生長曲線,如圖5A所示,csgA的敲除導致細菌在前4 h內生長較野生型慢,4 h后的生長情況則基本一致,但hnsA的敲除導致細菌生長明顯緩慢,且24 h后細
菌濃度較低。根據生長曲線,計算各菌株的倍增時間,結果如圖5B,野生型、EcN△csgA、EcN△hnsA的倍增時間分別為36、57、115 min。以上結果說明csgA對細菌的生長略有影響,
hnsA基因的缺失嚴重影響了EcN的生長,H-NS蛋白對EcN的生長起著較重要的作用。
2.4 EcN及其突變株的運動能力檢測
細菌在半固體培養基上生長形成的泳動圈可用于反映細菌的運動能力,EcN及其突變株的泳動圈如圖6,csgA突變株與EcN的泳動圈大小基本一致,直徑約為2 cm;而hnsA突變株的泳動圈明顯小于EcN和EcNΔcsgA,直徑只有0.5 cm左右。表明csgA的缺失并未影響細菌的運動能力,而hnsA的缺失會導致細菌的運動能力受到極大限制。
2.5 EcN及其突變株生物被膜定量分析
結晶紫染色法被廣泛應用于細菌生物被膜的半定量測定。EcN及其突變株在96孔板中用M9C培養基培養24 h,生物被膜的形成情況如圖7所示。從圖7可以看出,EcN野生型菌株與EcNΔcsgA菌株形成的生物被膜的量基本一致,而EcNΔhnsA菌株的生物被膜的形成量明顯高于野生型,說明curli對EcN生物被膜形成沒有明顯影響,而H-NS蛋白對EcN生物被膜的形成有抑制作用。
3 討論與結論
EcN是廣泛認可的益生菌,可通過形成生物膜在腸道中定殖,抑制腸道致病菌繁殖。因此,研究影響EcN生物被膜的形成將有助于研究宿主對病原體的抵抗力,是減緩生物被膜形成的治療學發展的重要理論基礎,為生物被膜預防和制定控制新策略提供思路。
該研究探究了curli菌毛和鞭毛對EcN生物被膜形成的影響。通過λ-Red同源重組系統構建curli菌毛合成基因(csgA)和鞭毛調控基因(hnsA)突變菌株,并對野生型、EcNΔhnsA、EcNΔcsgA的生長情況進行了監測,結果顯示,csgA的敲除導致細菌的生長速度較野生型略慢,hnsA的敲除則嚴重影響了EcN的生長,這可能是由于hnsA編碼的類組蛋白類核結構蛋白(H-NS)是全局調控因子,與DNA結合調控轉錄許多基因的表達,因而影響了細菌的生長能力。
細菌運動能力的降低會導致其黏附能力的增強,有利于生物被膜的初始黏附,由此可促進生物被膜的形成。泳動試驗結果顯示,csgA突變株與EcN運動能力基本相同,說明curli菌毛不影響細菌的運動能力。csgA突變株與EcN和生物被膜的形成量基本一致,說明csgA的缺失對細菌生物被膜的形成沒有影響,可能是由于EcN除curli外具有的另外2種菌毛(type1和F1C菌毛)彌補了curli菌毛缺失對生物被膜的影響。
研究表明,H-NS可調控細菌鞭毛基因的表達。hnsA突變株的運動能力明顯弱于野生型,說明H-NS對鞭毛是正調控作用,hnsA的缺失抑制了鞭毛的合成,使EcN的運動能力降低。且96孔板試驗顯示,hnsA的缺失導致細菌生物被膜形成明顯增多,說明hnsA突變株的運動能力降低,導致其黏附能力的增強,以此促進了生物被膜的形成。
參考文獻
[1] SUN F J,QU F,LING Y,et al.Biofilm-associated infections:Antibiotic resistance and novel therapeutic strategies[J].Future microbiology,2013,8(7):877-886.
[2] VOGELEER P,TREMBLAY Y D N,MAFU A A,et al.Life on the outside:Role of biofilms in environmental persistence of Shiga-toxin producing Escherichia coli[J].Frontiers in microbiology,2014,5:1-12.
[3] STEWART P S.Theoretical aspects of antibiotic diffusion into microbial biofilms[J].Antimicrobial agents and chemotherapy,1996,40(11):2517-2522.
[4] YAN H H,BAO F F,ZHAO L P,et al.Cyclic AMP(cAMP)receptor protein-cAMP complex regulates heparosan production in Escherichia coli strain nissle 1917[J].Applied and environmental microbiology,2015,81(22):7687-7696.
[5] DERIU E,LIU J Z,PEZESHKI M,et al.Probiotic bacteria reduce Salmonella typhimurium intestinal colonization by competing for iron[J].Cell host & microbe,2013,14(1):26-37.
[6] SHARMA V K,BEARSON B L.Hha controls Escherichia coli O157:H7 biofilm formation by differential regulation of global transcriptional regulators FlhDC and CsgD[J].Applied and environmental microbiology,2013,79(7):2384-2396.
[7] SALDAA Z,XICOHTENCATL-CORTES J,AVELINO F,et al.Synergistic role of curli and cellulose in cell adherence and biofilm formation of attaching and effacing Escherichia coli and identification of Fis as a negative regulator of curli[J].Environmental microbiology,2009,11(4):992-1006.
[8] SINGH K,MILSTEIN J N,NAVARRE W W.Xenogeneic silencing and its impact on bacterial genomes[J].Annual review of microbiology,2016,70(1):199-213.
[9] SOUTOURINA O,KOLB A,KRIN E,et al.Multiple control of flagellum biosynthesis in Escherichia coli:Role of H-NS protein and the cyclic AMP-catabolite activator protein complex in transcription of the flhDC master operon[J].Journal of bacteriology,1999,181(24):7500-7508.
[10] BERTIN P,TERAO E,LEE E H,et al.The H-NS protein is involved in the biogenesis of flagella in Escherichia coli[J].Journal of bacteriology,1994,176(17):5537-5540.
[11] ZHANG L,WENG Y,WU Y,et al.H-NS is an activator of exopolysaccharide biosynthesis genes transcription in Vibrio parahaemolyticus[J].Microbial pathogenesis,2018,116:164-167.
[12] DATSENKO K A,WANNER B L.One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products[J].Proceedings of the national academy of sciences of the United States of America,2000,97(12):6640-6645.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
小讀者(2021年2期)2021-03-29 05:03:48
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
