地理因素對箭竹復合體遺傳多樣性與遺傳分化的影響
2021-06-22 01:20:32邢曉成張雨曲
西北植物學報 2021年5期
關鍵詞:物種
黃 蕾,邢曉成,張雨曲,任 毅
(陜西師范大學 生命科學學院,藥用植物資源與天然藥物化學教育部重點實驗室, 西安 710119)
生物多樣性包括遺傳多樣性、物種多樣性和生態系統多樣性三個層次,是指全部陸地或水域中生物所擁有的基因、細胞、物種乃至多個生態系統的總稱。其中,遺傳多樣性是生物多樣性研究的核心和基礎,廣義是指地球上所有的生物所攜帶的全部遺傳信息的總和;狹義的遺傳多樣性則是指種內不同居群間以及居群內的遺傳變異,即種內基因的變化[1-3]。一般認為,影響物種水平上的遺傳多樣性因素可能包括該物種的進化歷史、分布范圍、繁育系統等,通常認為分布地域廣、基因流較強、種子較多的物種其遺傳多樣性較高,一個物種包含的等位基因越豐富,說明它對環境適應性就越強,進化潛力也越大[4]。
影響植物居群遺傳多樣性的內部因素包括物種的繁育系統(生殖方式), 遺傳漂變, 自然選擇, 基因突變[5]和基因流[6], 同時還包括由于環境變化和人為干擾引起的種群隔離和生境片斷化[7]等外部因素。內部因素可直接作用于基因組, 引起等位基因數目與頻率的變化, 外部因素不會直接改變基因 (等位基因) 的數目與頻率, 一般通過間接方式使植物居群的遺傳多樣性水平和遺傳結構發生變化[8-9]。近年來,隨著分子標記技術的蓬勃發展,探討不同環境因子對植物遺傳多樣性的影響成為領域熱點。環境因子一般分為生物因子和非生物因子[10],非生物因子中溫度、降水、緯度、海拔等地理因素成為研究者關注的重點[11]。值得關注的是不同植物類群中,同一地理因素與遺傳多樣性可能呈現出截然相反的關聯性。徐振朋等[12]利用15對ISSR分子標記對石竹科裸果木屬12個天然居群的遺傳多樣性進行分析,發現海拔和緯度與遺傳多樣性呈顯著的正相關。蘇前[13]基于100個RAPD分子標記對夾竹桃科羅布麻屬6個天然居群的研究發現遺傳多樣性與緯度呈顯著正相關,而與海拔呈顯著的負相關。因此,為了更好地理解和闡釋“地理因素如何影響植物居群的遺傳多樣性”這一重要問題,需來自更多植物類群的研究實例。
箭竹屬隸屬于禾本科竹亞科 (Poaceae,Bambusoideae) 北美箭竹族 (tribe arundinariinae) 或木本竹類溫帶分支, 是竹亞科中的大屬之一, 包括80~100種。該屬廣泛分布于中國西南至喜馬拉雅東部的山地, 是大熊貓等珍稀動物的主要食物, 在當地生態系統中發揮重要作用[14]。箭竹屬的分類地位和屬的范圍一直存在爭議, 易同培[15-16]曾經依據營養器官性狀將箭竹屬分為2組6系。然而,與繁育器官相比, 營養器官容易受環境影響而發生變化, 同時營養器官普遍存在趨同演化或平行演化現象。基于已有研究發現,箭竹屬內有15個近緣種——箭竹(FargesiaspathaceaFranch.)、矮箭竹(FargesiademissaYi)、毛龍頭竹(FargesiadecurvataJ. L. Lu)、拐棍竹(FargesiarobustaYi)、青川箭竹(FargesiarufaYi)、糙花箭竹(FargesiascabridaYi)、缺苞箭竹(FargesiadenudataYi)、華西箭竹[Fargesianitida(Mitford) Keng f. ex Yi)、秦嶺箭竹(FargesiaqinlingensisYi et J.X. Shao)、團竹(FargesiaobliquaYi)、伏牛山箭竹(F.funiushanensisYi)、神農箭竹[Fargesiamurielae(Gamble) Yi]、窩竹(Faregsiabrevissima)、細枝箭竹(Fargesiastenoclada)和甜箭竹(Fargesiaostrina)[17],以上物種之間的形態學差異不顯著,部分性狀存在連續變異,且在已有的系統發育樹上位于同一分支[18-19],而箭竹是其中最早命名的物種[20],因此本研究將其稱為箭竹復合體(Fargesiaspathaceacomplex)。本研究的前期工作篩選了箭竹復合體的39個天然居群共750個個體,利用14個SSR(微衛星 Simple Sequence Repeat)分子標記,詳細探討了該復合體內的遺傳變異式樣,并認為其中3個種是成立的,而有4個種顯然不成立。同時結合3種居群遺傳結構分析方法,將箭竹物種復合體的39個居群分為2個支系(A支系、B支系),其中A支系內部可再劃分出華西箭竹亞支系和團竹亞支系,B支系內部可再劃分出拐棍竹亞支系、窩竹亞支系、箭竹亞支系和糙花箭竹亞支系,即全部樣本分為6個亞支系。另一方面,還發現該復合體內的整體遺傳多樣性水平并不低,改變了“竹子以營養繁殖為主,遺傳多樣性水平較低”的傳統觀念[21]。但是需要指出的是,該研究集中在梳理箭竹復合體內部的遺傳結構和居群變異式樣,并未詳細探討環境因素,尤其是海拔和緯度對整個復合體內不同支系、亞支系的遺傳多樣性水平的影響。基于箭竹復合體現有的750個個體大量的遺傳變異信息,以及該類群對大熊貓保護領域的重要意義,本研究認為該復合體是深入探討“地理因素如何影響植物居群的遺傳多樣性”這一問題的理想材料。
另一方面,遺傳多樣性可為人們估算植物居群的遺傳分化提供重要的基礎數據。遺傳多樣性的計算是基于遺傳變異信息,這些遺傳變異在居群之間的不均衡分布表現為遺傳分化,而遺傳分化同樣受到地理因素的重要影響[8-9]。因此,本研究在深入分析前期研究工作的箭竹復合體SSR標記數據時,同時關注地理距離與箭竹復合體內的遺傳分化水平是否表現出顯著的相關性。根據前期野外觀察,箭竹復合體內同一支系內的不同類群存在海拔差異,這些類群可能發生了對高海拔區域的適應性分化,目前并不清楚地理距離是否影響了復合體內的適應性分化。綜上所述,本研究探討箭竹復合體的遺傳多樣性及其與地理因素的關系,旨在揭示箭竹復合體遺傳多樣性對地理變化的響應,為后續深入研究竹類植物對高山環境的適應性分化奠定基礎,并為箭竹復合體內遺傳多樣性保護和大熊貓主食竹相關研究提供科學依據和理論支持。
1 材料和方法
1.1 微衛星(SSR)引物篩選
本研究的測序樣品選擇箭竹物種復合體的4個竹種:缺苞箭竹(甘肅省文縣)、毛龍頭竹(陜西省太白縣)、細枝箭竹(四川省彭州縣)和伏牛山箭竹(河南省欒川縣),用其新鮮葉片,干冰保存送至上海天昊遺傳分析中心,利用 Illumina HiSeq 2500平臺進行配對末端(paire-end)測序。根據返回的高通量測序信息,在設定文庫片段大小時,在主峰基礎上±50 bp設定其最大與最小片段范圍,其中毛龍頭竹(310~410 bp)、缺苞箭竹(280~380 bp)、細枝箭竹(280~380 bp)和伏牛山箭竹(280~380 bp),以雙末端方式導入CLC Genomics Workbench 7.5(QIAGEN, Germantown, MD, Germany) 軟件中,先去除質量較低的序列,再用從頭組裝的策略對上一步得到的結果進行組裝,所得結果以*.fasta文件格式輸出。本研究所篩選14對SSR引物的詳細信息,參照黃蕾等文章中相應表格[18]。
1.2 箭竹復合體樣本采集
參考《中國植物志》[14]和《中國竹類圖志》[22],整理箭竹物種復合體中15個竹種的詳細地點,并歷時3年對其進行樣品采集。采集地點包括四川、陜西、貴州、湖北、河南、甘肅、寧夏等省份。樣品采集按照每個居群20~30份DNA材料(由于箭竹物種復合體竹種地下莖屬于合軸叢生,地上同一叢不同稈株來自同一個體,因此在樣品采集時,選擇個體間間隔15 m以上),總共采集39個居群,包括箭竹(2個居群)、矮箭竹(3個居群)、毛龍頭竹(7個居群)、拐棍竹(4個居群)、青川箭竹(2個居群)、糙花箭竹(2個居群)、缺苞箭竹(3個居群)、華西箭竹(3個居群)、秦嶺箭竹(4個居群)、團竹(1個居群)、伏牛山箭竹(1個居群)、神農箭竹(4個居群)、窩竹(1個居群)、細枝箭竹(1個居群)和甜箭竹(1個居群)。每個居群挑選15~20個個體,共計750個個體進行SSR片段的擴增和測序,詳細信息請見黃蕾等文章中相應表格[21]。
1.3 總DNA提取與PCR擴增
利用改良的CTAB[23-24]法提取竹類植物的總DNA, 步驟為: 1)取1.5 g的干燥葉片手工研磨至粉末狀,再將粉末狀顆粒轉入2 mL離心管中,加入800 μL DNA緩沖液Ⅰ,0 ℃保存30 min; 2)離心管放入4 ℃恒溫離心機,4 000 r/min恒溫離心10 min,棄上清; 3)將離心管取出,加入800 μL已預熱的DNA緩沖液Ⅱ(65 ℃),并在65 ℃水浴鍋中水浴1 h,間隔15 min輕微搖晃使顆粒與液體充分接觸; 4)取出離心管放置于4 ℃恒溫離心機中12 000 r/min離心10 min; 吸取上清液至2 mL離心管中,加800 μL的氯仿/異戊醇(體積比24∶1)與80 μL DNA緩沖液Ⅱ,顛倒混合10 min; 5) 4 ℃恒溫離心機中常溫10 000 r/min離心10 min,取上清液至2 mL離心管中,加入700 μL的氯仿/異戊醇(24∶1)與70 μL DNA緩沖液Ⅱ,顛倒混合10 min; 6)常溫10 000 r/min離心10 min,取上清液,加入等體積無水乙醇在-20 ℃冰箱放置1 h;10 000 r/min離心12 min,加入400 μL ddH2O,完全溶解; 7)加入500 μL冰乙醇在-20 ℃放置1 h,12 000 r/min離心10 min,棄上清液,用70%乙醇洗滌沉淀,干燥過夜; 8)加入200 μL ddH2O溶解DNA,-20 ℃保存備用。
PCR擴增首輪反應——10 μL的反應體系,包括酶 2×Es Taq MasterMix 5 μL,ddH2O 4 μL,正反向引物各 0.25 μL,DNA模板0.5 μL。反應程序采用梯度 PCR(TD-PCR)程序:94 ℃ 4 min; 94 ℃ 30 s,62~0.5 ℃, 20 s,72 ℃ 30 s,10個循環; 94 ℃ 30 s, 57 ℃ 20 s,23~24 循環, 72 ℃ 30 s; 72 ℃ 10 min; 25 ℃。第二輪反應——10 μL反應體系,包括酶 2×Es Taq MasterMix 10 μL,ddH2O 8 μL,熒光引物正反向各0.5 μL,DNA模板(首輪反應產物稀釋100倍)1 μL。反應程序: 94 ℃ 4 min; 94 ℃ 30 s, 62~0.5 ℃30 s, 72 ℃ 30 s,10個循環; 94 ℃ 30 s, 57 ℃ 30 s, 72 ℃ 30 s, 23~24 循環; 72 ℃ 10 min, 25 ℃。兩輪反應中,不同的引物采用不同的熒光進行標記。擴增產物用1.5%瓊脂糖凝膠電泳檢測后送至江蘇金唯智生物科技有限公司進行毛細管電泳檢測,使用ABI3730XL測序儀完成熒光檢測,得到擴增產物的峰型圖譜。
1.4 數據處理與分析
SSR原始數據進行匯總整理后,輸入CONVERT 1.3[25]軟件進行格式轉換,用于后續軟件分析。使用POPGENE 1.32軟件[26]進行遺傳多樣性——平均等位基因數(Na)、有效等位基因數(Ne)、香農指數(I)、擴增多態性位點比率(PPL)、期望雜合度(He)、觀測雜合度(Ho)的計算。本研究所用的線性回歸分析采用R軟件包進行計算并繪圖[27]。在遺傳分化指標上,利用GenAlEx 6.502軟件進行Mantel檢驗[28],通過1 000次迭代運行計算地理距離與遺傳距離的相關性。本研究采用的地理距離是基于居群經緯度所得的歐式距離。
2 結果與分析
2.1 海拔和緯度對箭竹復合體遺傳多樣性的影響
基于750個個體和14對SSR引物的遺傳結構分析,39個居群可分為A, B兩個支系,A支系可進一步分為2個亞支系,B支系可分為4個亞支系[21]。首先對A, B兩個支系的遺傳多樣性參數——平均等位基因數(Na)、有效等位基因數(Ne)、香農指數(I)、擴增多態性位點比率(PPL)、期望雜合度(He)、觀測雜合度(Ho)進行計算,A支系遺傳多樣性參數的結果為Na=2.93,Ne=1.85,I=0.64,Ho= 0.47,He=0.37,PPL=0.81,B支系為Na=3.05,Ne=1.91,I=0.66,Ho=0.45,He=0.37,PPL=0.86。由圖1可見,B支系的遺傳多樣性略高于A支系。
對6個亞支系的遺傳多樣性參數進行計算(圖2),華西箭竹亞支系內13個居群Na=6.21,Ne=2.05,I=0.84,Ho=0.50,He=0.44,PPL=0.819;團竹亞支系內4個居群Na=4.14,Ne=1.69,I=0.61,Ho=0.37,He=0.33,PPL=0.732;拐棍竹亞支系內4個居群Na=4.57,Ne=1.84,I=0.73,Ho=0.40,He=0.38,PPL=0.857;窩竹亞支系內9個居群Na=5.43,Ne=2.09,I=0.80,Ho=0.43,He=0.41,PPL=0.821;箭竹亞支系內4個居群Na=4.50,Ne=2.08,I=0.76,Ho=0.47,He=0.39,PPL=0.875;糙花箭竹亞支系內5個居群Na=5.50,Ne=2.38,I=1.00,Ho=0.51,He=0.50,PPL=0.929。6個亞支系遺傳多樣性由高到低的順序為:糙花箭竹亞支系>華西箭竹亞支系>窩竹亞支系>箭竹亞支系>拐棍竹亞支系>團竹亞支系。在遺傳多樣性指標上,擴增多態性位點比率PPL在不同亞支系之間變化最小;平均等位基因數Na的變化最大。
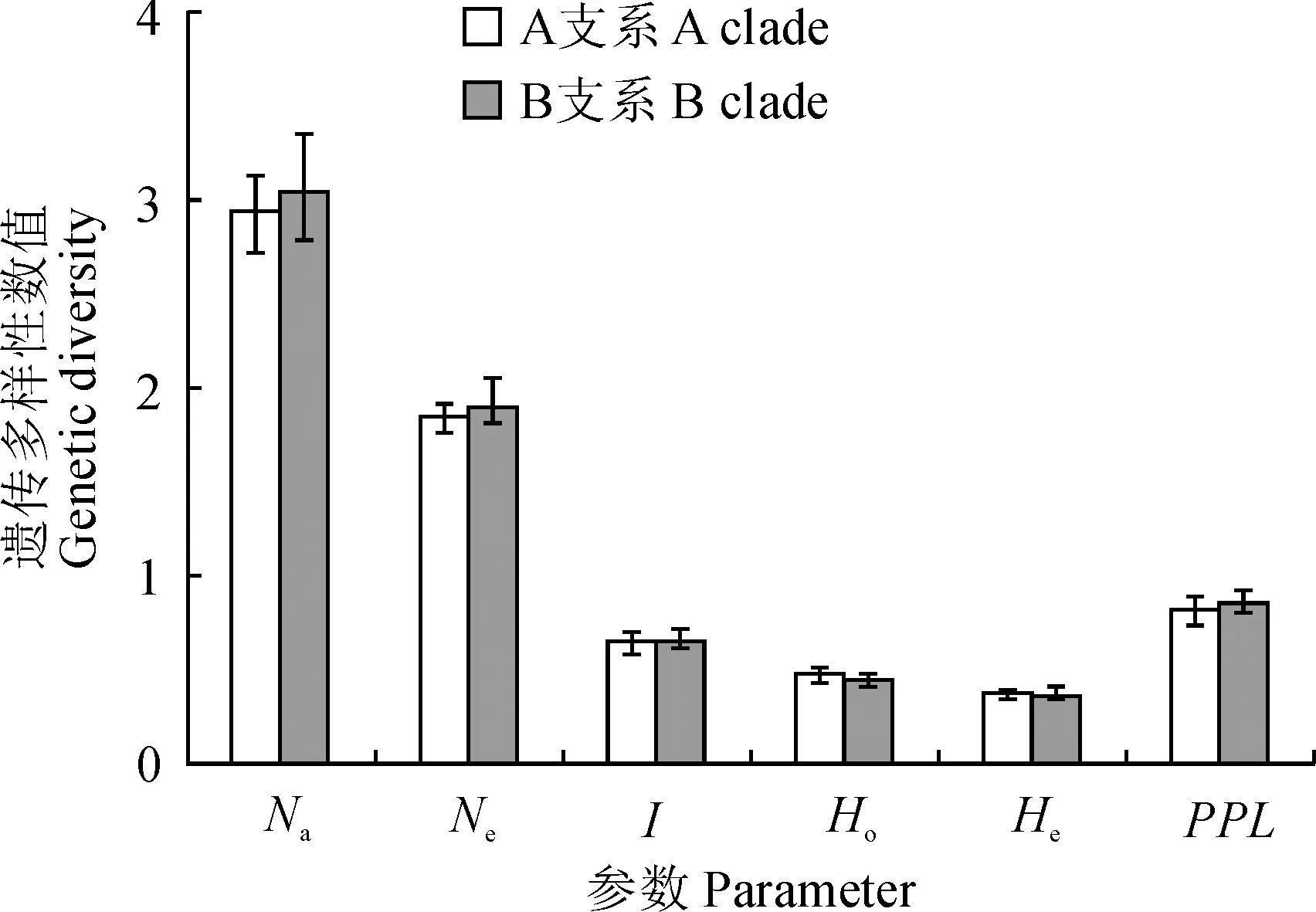
圖1 箭竹復合體A、B兩支系內遺傳多樣性參數對比Fig.1 The genetic diversity of A and B clades in Fargesia spathacea complex

圖2 箭竹復合體6個亞支系組內遺傳多樣性參數對比圖Fig.2 The genetic diversity of six sub-clades in F. spathacea complex
為檢測箭竹物種復合體A支系和B支系的遺傳多樣性與海拔、緯度是否具有顯著相關性,將兩組各自的遺傳多樣性指標He與海拔和緯度分別進行線性回歸分析,結果如圖3所示。A支系的遺傳多樣性與緯度、海拔均具有顯著相關(圖3, A支系:R2=0. 5635,r=0.751,P<0.01;R2=0.0151,r=0.123,P<0.01),B支系也可得出同樣結論(圖3, B支系:R2=0.0700,r=0.264,P<0.01;R2=0.1302,r=0.360,P<0.01)。這表明隨著緯度和海拔的增加A支系和B支系的遺傳多樣性也隨之增大。
為檢測箭竹物種復合體6個亞支系的遺傳多樣性與海拔、緯度是否具有顯著相關性,將每個亞支系的遺傳多樣性指標He與海拔和緯度分別進行線性回歸分析。拐棍竹亞支系He與緯度的相關性R2=0.3772,r=0.615,P<0.01;He與海拔的相關性R2=0.1948,r=-0.442,P<0.01。窩竹亞支系He與緯度的相關性R2=0.0002,r=0.014,P<0.01;He與海拔的相關性R2=0.6353,r=0.797,P<0.01。箭竹亞支系He與緯度的相關性R2=0.7542,r=-0.868,P<0.01;He與海拔的相關性R2=0.3406,r=0.584,P<0.01。糙花箭竹亞支系He與緯度的相關性R2=0.2314,r=0.481,P<0.01;He與海拔的相關性R2=9E-06,r=0.003,P<0.01。華西箭竹亞支系He與緯度的相關性R2=0.0935,r=0.306,P<0.01;He與海拔的相關性R2=0.1945,r=-0.441,P<0.01。團竹亞支系He與緯度的相關性R2=0.6121,r=0.782,P<0.01;He與海拔的相關性R2=0.0442,r=0.210,P=0.02。由此可見,盡管6個亞支系的遺傳多樣性和緯度、海拔呈現出顯著相關性,但表現出不同的趨勢,部分支系表現為負相關。
2.2 地理距離對箭竹復合體遺傳分化式樣的影響
利用GenAlEx 6.41在箭竹物種復合體水平和A、B支系水平分別進行Mantel檢驗(圖4)。在箭竹復合體水平上,全部39個居群的遺傳距離和地理距離未表現出顯著的線性關系(R2=0.0032,r=0.057,P=0.11)A支系的檢測結果表明遺傳距離和地理距離存在顯著相關性(R2=0.0613,r=0.25,P=0.01),B支系的檢測結果表明遺傳距離和地理距離不存在顯著相關性(R2=0.0036,r=0.06,P=0.21)。
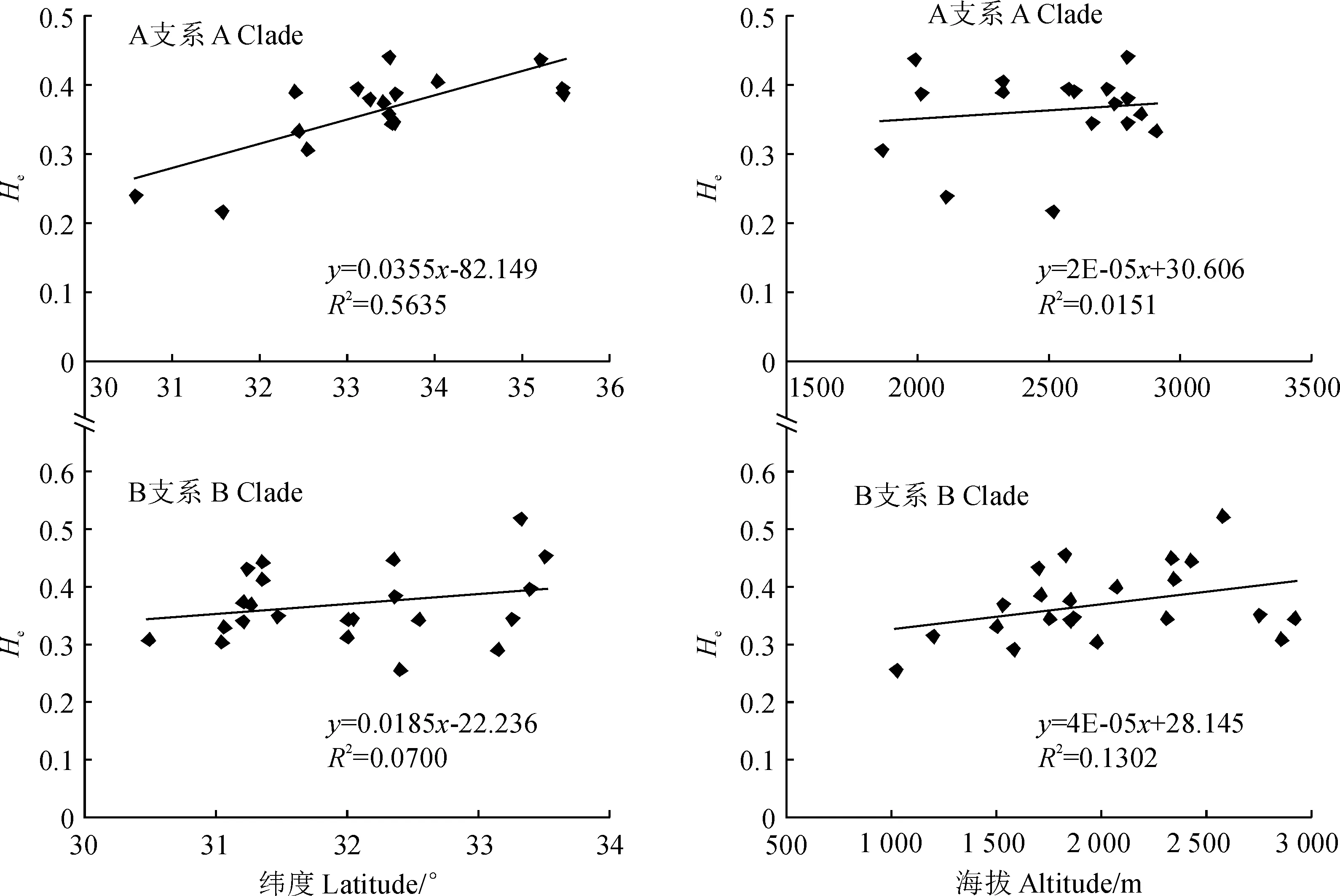
圖3 箭竹復合體A、B兩支系遺傳多樣性水平與緯度、海拔的回歸分析圖Fig.3 The regression analysis of genetic diversity and latitude, altitude within A and B clade in F. spathacea complex

Ⅰ. 箭竹復合體; Ⅱ. A支系; Ⅲ. B支系圖4 不同劃分組下遺傳距離和地理距離 之間的Mantel 檢驗Ⅰ. Fargesia spathacea complex; Ⅱ. A clade; Ⅲ. B cladeFig.4 The Mantel test of genetic distance and geographic distance under different groups
在6個亞支系水平上, Mantel 檢驗的結果如圖5所示。 拐棍竹亞支系(a)檢測結果不顯著,相關系數R2=0.3944,r=0.63,P=0.15;糙花箭竹亞支系(b)檢測結果不顯著,相關系數R2=0.1153,r=0.34,P=0.15;窩竹亞支系(c)檢測結果不顯著,相關系數R2=0.0558,r=0.24,P=0.10;箭竹亞支系(d)檢測結果不顯著,相關系數R2=0.0223,r=0.15,P=0.58;華西箭竹亞支系(e)檢測結果不顯著,相關系數R2=0.011,r=0.11,P=0.40;團竹亞支系(f)檢測結果顯著,相關系數R2=0.3571,r=0.60,P=0.02。由此可見,B支系內4個亞支系均未表現出遺傳距離和地理距離之間的顯著相關性,這一結果與前述結果一致;A支系內2個亞支系僅團竹亞支系的遺傳距離和地理距離存在較弱的顯著相關性。
3 討 論
影響植物居群遺傳多樣性的因素包括物種的生殖方式、遺傳漂變、自然選擇、突變率、基因流,以及種群隔離和生境片斷化。另一方面,樣本大小和分子標記數量也能對遺傳多樣性的估算產生一定影響[29-30]。在本研究中,華西箭竹亞支系的居群數目最多,但其遺傳多樣性并非最高。由此可見,在亞支系水平,樣本大小對估算箭竹復合體遺傳多樣性的影響較小。這表明,箭竹復合體的不同類群在適應不同生境時,遺傳多樣性水平可能會受到其他生態因子或進化因素的影響。其次,在地理因素與遺傳多樣性的相關性上,不同物種的研究結果并不一致[9-10]。在本研究中,地理因素與遺傳多樣性的相關性在不同進化尺度上并不相同。在A、B支系水平上檢測出顯著相關性,可能是掩蓋亞支系之間的遺傳結構的結果,并不能反映真實情形。因此建議,開展類似研究工作時應充分考慮遺傳結構的影響,先厘清樣本內部的遺傳分化,再開展多因素與遺傳多樣性的相關性檢測。
在地理距離與遺傳距離的相關性上,A支系內包含華西箭竹亞支系和團竹亞支系,本研究結果表明地理隔離在一定程度上影響了其現有分布格局;而B支系內的遺傳結構更為復雜,其分布格局可能受其他因素比如自然選擇、基因流、遺傳漂變等影響。據此推測,地理距離并非是影響箭竹復合體內遺傳分化的主導因素。依據本研究前期的發現,B支系下的糙花箭竹和窩竹亞支系內的部分個體檢測到明顯的雜交跡象,這也為B支系Mantel檢驗結果不顯著提供了合理解釋[21]。后續需加強基因流和雜交事件的檢測,以明確不同亞支系之間的基因流是否參與了現有遺傳分化格局的建立。
本研究通過對箭竹復合體內39個居群的14對SSR數據進行深入分析,發現糙花箭竹亞支系具有最高的遺傳多樣性,而團竹亞支系的遺傳多樣性最低。盡管在A、B兩個支系水平,遺傳多樣性和緯度、海拔呈現了顯著正相關,但在更精細的亞支系水平,遺傳多樣性的變化趨勢呈現出更為復雜的局面,部分支系表現為負相關。由此推測,箭竹復合體的不同類群在適應不同生境時,遺傳多樣性可能會受到其他生態因子或進化因素的影響。另一方面,通過Mantel檢驗結果發現地理距離并非是影響箭竹復合體內遺傳分化的主導因素。B支系內的4個亞支系,均未表現出遺傳距離和地理距離之間的顯著相關性;A支系中僅團竹亞支系的遺傳距離和地理距離存在較弱的顯著相關性,這表明自然選擇、基因流、遺傳漂變等因素可能影響了該復合體內的遺傳分化。已有結果表明該復合體內部分居群之間可能存在基因流,后續需結合基因流檢測推斷雜交事件對其遺傳分化的影響。鑒于箭竹復合體內不同類群之間存在海拔分化,本研究的結果可為后續深入研究竹類植物的適應性分化奠定基礎,亦可為大熊貓主食竹的相關研究提供重要的參考信息。
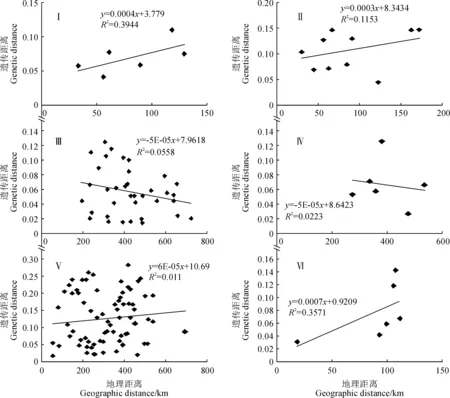
Ⅰ. 拐棍竹亞支系; Ⅱ.糙花箭竹亞支系; Ⅲ.窩竹亞支系; Ⅳ.箭竹亞支系; Ⅴ.華西箭竹亞支系; Ⅵ.團竹亞支系圖5 6個亞支系下遺傳距離和地理距離之間的Mantel 檢驗Ⅰ. F. robusta sub-clade; Ⅱ. F. scabrida sub-clade; Ⅲ. F. brevissima sub-clade; Ⅳ. F. spathacea sub-clade; Ⅴ. F. nitida sub-clade; Ⅵ. F. obliqua sub-cladeFig.5 The Mantel test of genetic distance and geographic distance within six sub-clades
作者貢獻:邢曉成和張雨曲采集了全部樣本;邢曉成完成了SSR數據的測序和數據分析;任毅教授設計了研究方案;邢曉成和黃蕾完成文章撰寫。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
