塔瑪亞歷山大藻產麻痹性貝類毒素能力的研究
2021-07-08 11:56:08張海燕隋延鳴湯云瑜劉淑晗蔡友瓊沈曉盛
海洋漁業 2021年3期
汪 宇,張海燕,隋延鳴,湯云瑜,孔 聰,王 媛,劉淑晗,蔡友瓊,沈曉盛
(1.中國水產科學研究院東海水產研究所,上海 200090;2.上海海洋大學食品學院,上海 201306)
麻痹性貝類毒素(paralytic shellfish poisoning,PSP)是一類海洋生物毒素的統稱,常被發現于雙殼貝類中,主要作用于鈉離子的傳輸通道,阻礙神經沖動的傳輸,從而產生麻痹作用。人類食用攜帶PSP的貝類后極易引起食物中毒,甚至死亡,死亡率可達1.5%[1]。目前,已發現的PSP組分至少有24種[2],主要來源于有毒的海洋甲藻,包括亞歷山大藻屬(Alexandrium)、鏈狀裸甲藻(Gymnodiniumcatenatum)和巴哈馬麥甲藻(Pyrodiniumbahamense)等。而最近的一些研究發現,某些海洋藍細菌也能夠產生PSP[3]。已有專家探究了溫度、光照、鹽度、養分、pH等對亞歷山大藻屬的微小亞歷山大藻(A.minutum)和塔瑪亞歷山大藻(A.tamarense)生長及毒素組成的影響[4-11]。研究這些地理上廣泛分布的有毒海洋甲藻產PSP的能力,有助于對藻類毒性引起的海洋污染風險進行評估。
就我國來看,PSP污染情況不容樂觀,僅2017—2019年,我國福建漳州和河北秦皇島就發生多起PSP中毒事件,不僅威脅到人類的生命安全,也造成了海水養殖業的巨大經濟損失。對此,我國政府高度重視,組織開展PSP監測項目,完善PSP風險監控體系。相關科研人員也積極投入PSP研究中,比如,研發更高效的PSP檢測手段、探究PSP在貝類中的代謝規律、分析產PSP藻類在不同環境中的生長及產毒能力等[12-13]。有研究表明,全球絕大多數PSP事件皆由亞歷山大藻屬藻類引起,該屬藻類目前已通過形態學確定的有30多種,其中一半以上被發現能夠產生PSP,且毒素成分多樣[1,14]。ZOU等[15]對67株分離自我國沿海的塔瑪亞歷山大藻PSP組分研究發現,不同藻株PSP成分存在較大差異,但主要成分基本相似。然而,對于該物種產PSP能力的研究仍然較有限。
塔瑪亞歷山大藻是一種典型的產PSP的甲藻,全球分布廣泛,是造成我國貝類受PSP污染的主要藻類之一。相關專家對塔瑪亞歷山大藻產毒情況進行了 研究[5,8,11,16-17],但對該 藻產PSP能力的研究仍不夠透徹,迄今尚少見外界環境因素對塔瑪亞歷山大藻中PSP組成影響的研究報道[18],其毒性強弱對人類生命安全的風險也未見評估。因此,本文選取塔瑪亞歷山大藻作為研究對象,將塔瑪亞歷山大藻置于不同鹽度、生長空間和營養素濃度下培養,分析其總毒素及各毒素組分含量,探究其產PSP能力,以期為PSP的基礎研究以及標準品制備等提供理論支持。
1 材料與方法
1.1 實驗材料
1.1.1 藻株來源
塔瑪亞歷山大藻GY-H31于2018年9月購自光語生物科技有限公司,藻株分離自我國東海長江入海口。
1.1.2 基礎培養基及海水
藻類培養所需培養基參考GUILLARD和RYTHER[19]的改良f/2培養基,自行配置。
海水由天然海鹽配置,海鹽購自以色列Red Sea公司,按照25.0 g海鹽∶1 L超純水配置成鹽度為25的海水。
1.1.3 化學試劑
冰乙酸(分析純),購自國藥集團化學試劑有限公司;甲酸(色譜純)、甲酸銨(色譜純)、乙腈(色譜純),購自美國J.T.Baker公司。
PSP標準品,包括石房蛤毒素(saxitoxin,STX)、新石房蛤毒素(neosaxitoxin,NEO)、膝溝藻毒素1-4(gonyautoxin 1-4,GTX1-4)、N-磺酰氨甲酰基類毒素1-2(N-sulfocarbamoylgonyautoxin 1-2,C1-2)、N-磺 酰 氨 甲 酰 基 類 毒 素5(gonyautoxin 5,GTX5)、脫氨甲酰基石房蛤毒素(decarbamoylsaxitoxin,dcSTX)、脫氨甲酰基新石房蛤毒素(decarbamoylneosaxitoxin,dcNEO)、脫氨甲酰基膝溝藻毒素 2-3(decarbamoylgonyautoxins 2-3,dcGTX2-3)均購自加拿大國家海洋生物研究所(National Research Council Canada)。
1.1.4 主要儀器
Ultimate 3000超高壓液相色譜-Q-Exactive靜電場軌道阱高分辨質譜聯用系統,美國Thermo Fisher公司;16RXⅡ高速冷凍離心機,日本HITACHI CF公司;Milli-Q超純水機,美國Millipore公司;VCX500超聲波破碎儀,美國Sonics公司;MGC-300B光照培養箱,上海一恒科學儀器有限公司;XSP-44X9多用途生物顯微鏡,上海光學儀器一廠;超潔凈工作臺,北京東聯哈爾儀器制造有限公司;LT-CPS80D高壓蒸汽滅菌鍋,澳大利亞LERD-Tech公司。
1.2 實驗方法
1.2.1 藻株的活化與確認
藻株4℃轉移至實驗室后,取出1 mL置于顯微鏡下進行鑒定[20],其余藻液立即接種于鹽度為25的f/2培養基中,置于25℃、光照4 000 lx、12∶12光暗循環的光照培養箱中培養。約十天取出1 mL于顯微鏡下觀察,視野中藻細胞均處于游動狀態,且存在細胞壁相連的細胞,表明藻種活力良好,可用于實驗。
1.2.2 藻株的保存
塔瑪亞歷山大藻培養二十天左右后,傳代接種,并按照活化條件培養,以保留藻種。
1.2.3 空間效應對塔瑪亞歷山大藻產毒影響
設置0.1、0.5、1.0 L共3個生存空間(即f/2培養基體積)梯度,每個梯度3組重復。將指數生長期的塔瑪亞歷山大藻分別接種于含有以上體積培養基的250 mL、1 L、2 L錐形瓶中,初始密度400個·mL-1、溫度25℃,鹽度25、光照4 000 lx、12∶12光暗循環。每天定時振蕩錐形瓶。在培養期第18天(以接種日期為第0天計算)收集相同數量的藻細胞進行PSP測定。
1.2.4 鹽度對塔瑪亞歷山大藻產毒影響
依據塔瑪亞歷山大藻能夠存活的鹽度范圍,設置15、25、35共3個鹽度梯度,每個梯度3組重復。將指數生長期的塔瑪亞歷山大藻接種于含有1 L f/2培養基的2 L錐形瓶中,初始密度為400個·mL-1、溫度25℃、光照4 000 lx、12∶12光暗循環。每天定時振蕩錐形瓶。在培養期第18天收集相同數量的藻細胞進行PSP測定。
1.2.5 營養素濃度對塔瑪亞歷山大藻產毒影響
設置1/4倍、1倍、4倍3個營養素濃度梯度,每個梯度3組重復。將指數生長期的塔瑪亞歷山大藻分別接種于含有1 L的1/4倍、1倍和4倍f/2培養基濃度的2 L錐形瓶中,初始密度為400個·mL-1、溫度25℃、鹽度25、光照4 000 lx,12∶12光暗循環。每天定時振蕩錐形瓶。在培養期第18天收集相同數量的藻細胞進行PSP測定。
1.2.6 PSP測定
1.2.6.1 毒素提取
離心收集相同數量的藻細胞于50 mL聚丙烯離心管中,加入20 mL 0.05 mol·L-1的乙酸溶液,在冰浴條件下于細胞破碎儀中超聲處理5 min(200 W,超20 s停40 s),隨后超聲波輔助提取15 min。4℃下4 000 r·min-1離心10 min,取上清液1 mL于超濾離心管(10 000 u)中,4℃下10 000 r·min-1離心10 min,取超濾液于進樣小瓶中,4℃下保存待分析。
1.2.6.2 儀器條件
儀器條件參考GB 5009.213-2016中液相色譜-質譜條件[21]。樣品分析在Ultimate 3000超高壓液相色譜-Q-Exactive靜電場軌道阱高分辨質譜聯用系統上進行。色譜柱為TSK-gel Amide-80(3μm,2 mm×15 cm)。
色譜條件:柱溫30℃,樣品溫度為4℃,進樣量10μL,流動相A為甲酸銨緩沖溶液(含5 mmol·L-1甲酸銨和0.1%甲酸),B為酸化乙腈(含5 mmol·L-1甲酸銨和0.1%甲酸),梯度洗脫條件:0~3 min,70%B;3~4 min,70%~60%B;4~16 min,60%B;16~19 min,60%~5%B;19~23 min,5%B;23~27 min,5%~70%B;27~35 min,70%B。
質譜條件:采用電噴霧離子源(ESI)平行反應監測(PRM)模式,噴霧電壓:3 200 V(+),2 800 V(-);鞘氣:40氣體流速單位(arb);輔助氣:10氣體流速單位(arb);吹掃氣:1氣體流速單位(arb);氣體加熱溫度:350℃;離子傳輸管溫度:325℃。
1.3 數據分析
實驗數據采用SPSS 22.0和Origin Pro 8軟件進行處理。采用單因子方差分析,并進行事后多重比較(Duncan檢驗),P<0.05表示差異顯著。
試樣中PSP總毒素含量(STXeq)參考如下公式計算:
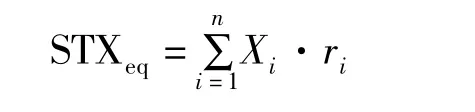
式中,STXeq為試樣中PSP總毒素含量(μg);Xi為各種PSP的含量(μg),ri為PSP的毒性因子(表1)。

表1 PSP毒性因子Tab.1 Toxicity factor of PSP
2 結果與分析
2.1 塔瑪亞歷山大藻中PSP組分分析
表2是塔瑪亞歷山大藻經培養后收集獲得的1.0×106個藻細胞中PSP組成及含量。從表2中可以看出,塔瑪亞歷山大藻至少含有13種PSP組分,可以推測塔瑪亞歷山大藻是貝類中PSP的來源之一。ANDERSON等[14]的報道也證實了這一點。從各組分的含量來看,不同組分的含量差異較大,每1.0×106個塔瑪亞歷山大藻細胞能夠檢出17.171μg毒素,GTX1、GTX4、GTX5、C1、C2為主要成分,占毒素總含量的97.2%(質量分數),其中僅GTX1和GTX4為高毒性成分,表明塔瑪亞歷山大藻的毒性強弱主要體現在GTX1和GTX4含量的高低。

表2 塔瑪亞歷山大藻中PSP毒素成分及含量Tab.2 Toxin profile and content of PSP in A.tamarense
2.2 不同PSP組分定量的最低藻細胞數量分析
塔瑪亞歷山大藻中PSP組分定量與藻細胞數量關系如圖1所示。不同毒素組分的定量與藻細胞數量高低存在明顯差異,當藻細胞數量低于4.0×103個時,13種組分均不能定量檢出;當藻細胞數量在4.0×103個至5.9×106個之間時,不能完全定量出13種組分;只有當藻細胞數高于5.9×106個時,13種組分才均能夠檢出。從PSP各組分定量所需藻細胞數量表明,GTX1和GTX4檢出定量所需藻細胞量最少,高于4.0×103個藻細胞就可以達到定量分析要求;dcSTX和dcNEO定量所需藻細胞數量最多,藻細胞數量必須分別為3.0×106個和5.9×106個以上;STX、NEO、GTX2和GTX3需要至少1.0×105個藻細胞才可被定量;其余組分定量所需藻細胞數基本處于1.0×104~1.0×105個。該結果與表2中毒素含量百分比不完全相同,質量分數高的毒素其定量所需藻細胞數不一定高,說明各組分定量所需藻細胞數還與毒素本身響應值有關。為保證13種毒素成分均能夠更方便、快捷地被檢出,本實驗將藻細胞數均取值在6.0×106個以上。
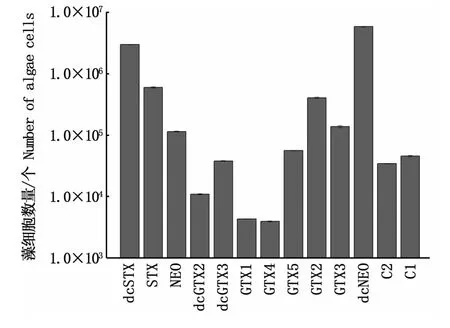
圖1 不同PSP組分定量限與藻細胞數之間的關系Fig.1 Relationship between LOQ of each PSP toxin and the number of A.tamarense cells
2.3 空間效應對塔瑪亞歷山大藻產毒能力的影響
圖2是3種不同空間效應培養條件下收集的相同數量的藻細胞的總毒素含量。可以看出,空間效應對塔瑪亞歷山大藻產毒能力的影響存在一定的差異,0.1 L與1.0 L培養條件下的總毒素含量表現出顯著性差異(Duncan檢驗,P<0.05),說明在相同環境下,培養空間與PSP的合成存在一定相關性。圖3的結果表明,空間效應會影響總毒素含量,但對各組分的定量檢出沒有影響,13種PSP均能檢出。但毒素組分的含量在不同空間效應下存在一定差異,其中dcSTX、STX、GTX2含量均表現出顯著性差異(P<0.05),dcSTX、STX在0.1 L培養條件下含量最高,分別為0.27、0.33μg,GTX2在1.0 L培養條件下含量最高,為0.12μg。而GTX1、GTX3均未出現由生長空間引起的顯著性差異(P>0.05),這表明空間效應不會影響塔瑪亞歷山大藻合成GTX1、GTX3,其總毒素含量的差異主要體現在其余毒素成分含量的高低。
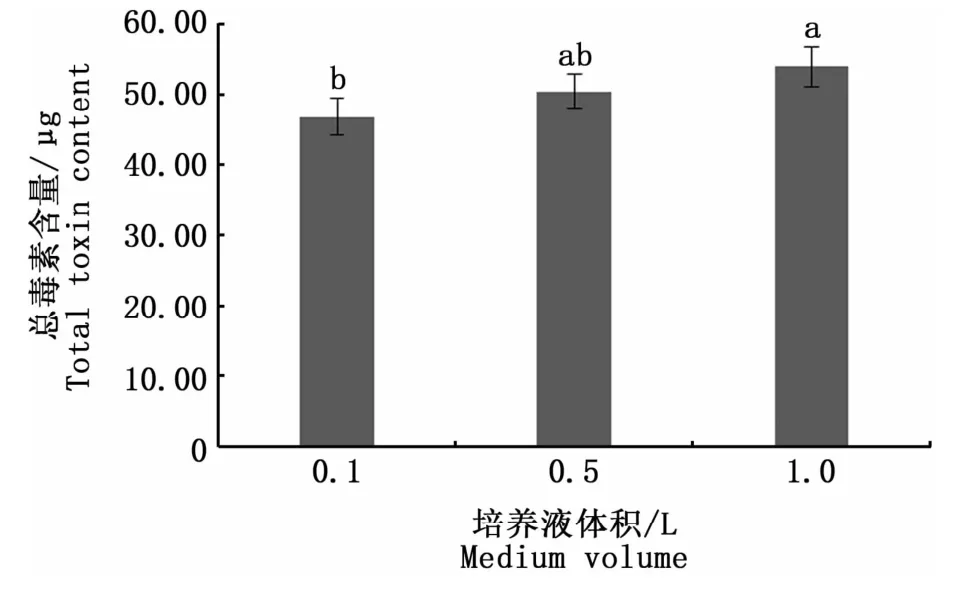
圖2 空間效應對塔瑪亞歷山大藻中總毒素含量的影響Fig.2 Impact of spatial effects on the total toxin content in A.tamarense
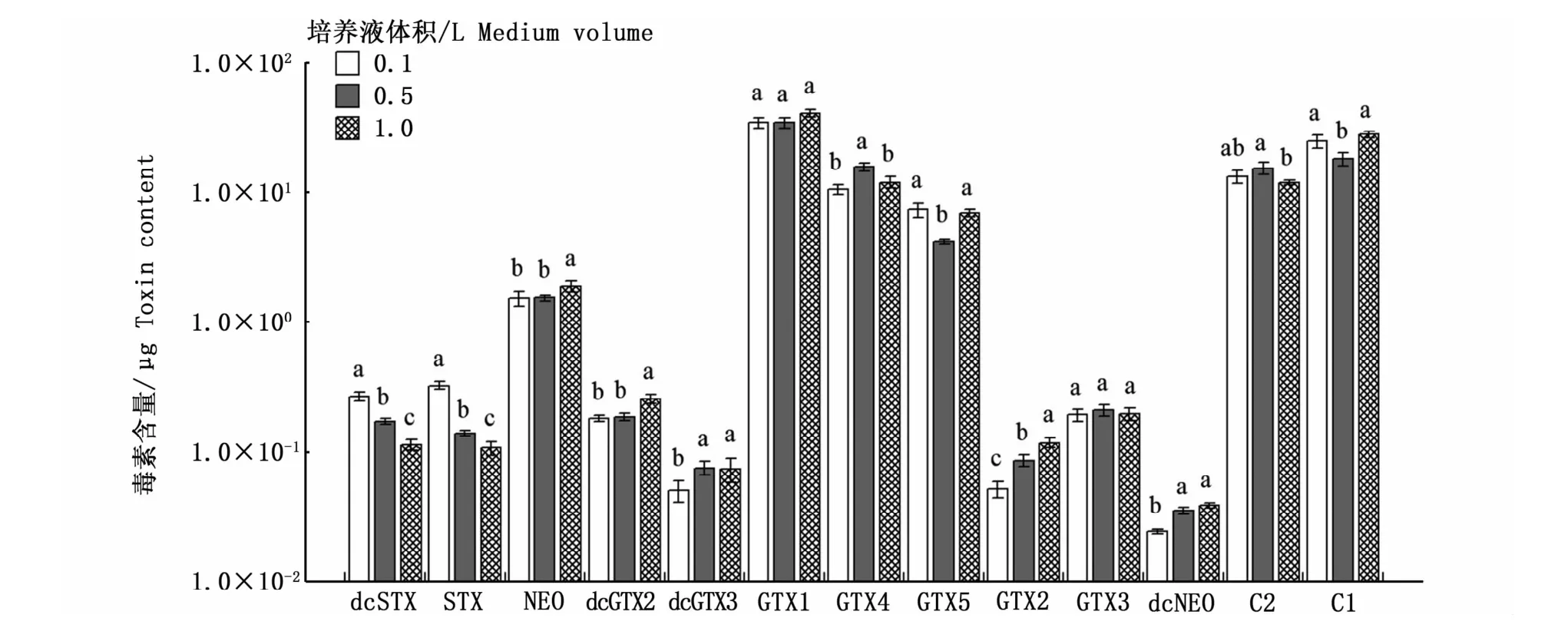
圖3 空間效應對塔瑪亞歷山大藻中PSP各組分含量的影響Fig.3 Impact of spatial effects on each PSP toxin content in A.tamarense
2.4 鹽度對塔瑪亞歷山大藻產毒能力的影響
圖4是3種不同鹽度培養條件下收集的相同數量藻細胞的總毒素含量,可以看出,鹽度25培養組塔瑪亞歷山大藻總毒素含量與鹽度35培養組無顯著性差異(P>0.05),但均顯著高于鹽度15培養組(P<0.05)。說明在相同環境下,鹽度對毒素的合成具有一定的影響。圖5是鹽度對塔瑪亞歷山大藻中PSP各組分含量影響,觀察到各組分均能夠定量檢出,且單一組分的含量在不同鹽度條件下均存在一定的差異。其中鹽度對STX、NEO、dcGTX2、GTX4、GTX2、GTX3、dcNEO、C1形成均表現出顯著性影響(P<0.05)。NEO、dcGTX2、GTX2、GTX3、dcNEO、C1含量均在鹽度35培養組達到峰值,分別為2.18、0.31、0.34、0.32、0.06、34.86μg;STX含量在鹽度15培養組最高,為0.20μg;GTX4含量在鹽度25培養組最高,為12.00 μg。但鹽度不影響塔瑪亞歷山大藻dcGTX3的合成(P>0.05)。GTX1含量在鹽度25培養組與鹽度35培養組無顯著性差異(P>0.05),這可能是鹽度25培養組和鹽度35培養組總毒素含量無顯著差異的一個原因。
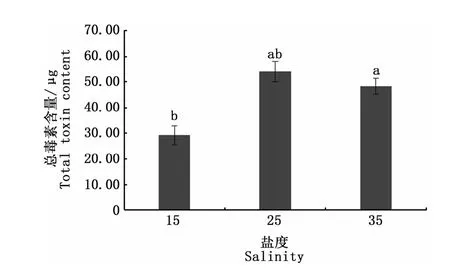
圖4 鹽度對塔瑪亞歷山大藻總毒素含量的影響Fig.4 Impact of salinity on the total toxin content in A.tamarense
2.5 營養素濃度對塔瑪亞歷山大藻產毒能力的影響
圖6是3種營養素濃度培養條件下收集的相同數量藻細胞的總毒素含量,可以看出,營養素濃度對塔瑪亞歷山大藻的總毒素含量有顯著影響(P<0.05),總毒素含量在1/4倍營養素濃度下達到最高,為89.15μg;而在4倍營養素濃度時降到最低,僅有42.62μg。表明在相同環境下,塔瑪亞歷山大藻總毒素含量與營養素濃度呈負相關。圖7顯示,13種毒素組分均在1/4營養素濃度時達到最高。在本研究中,營養素濃度對dcGTX2、dcGTX3、GTX1、GTX4、GTX5、GTX3、C1、C2的合成均有顯著影響(P<0.05)。
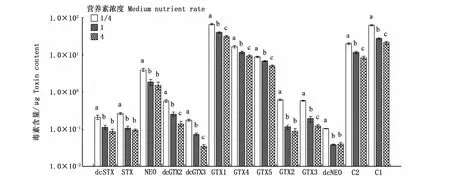
圖7 營養素濃度對塔瑪亞歷山大藻中PSP各組分含量的影響Fig.7 Impact of medium nutrient rate on each PSP toxin content in A.tamarense
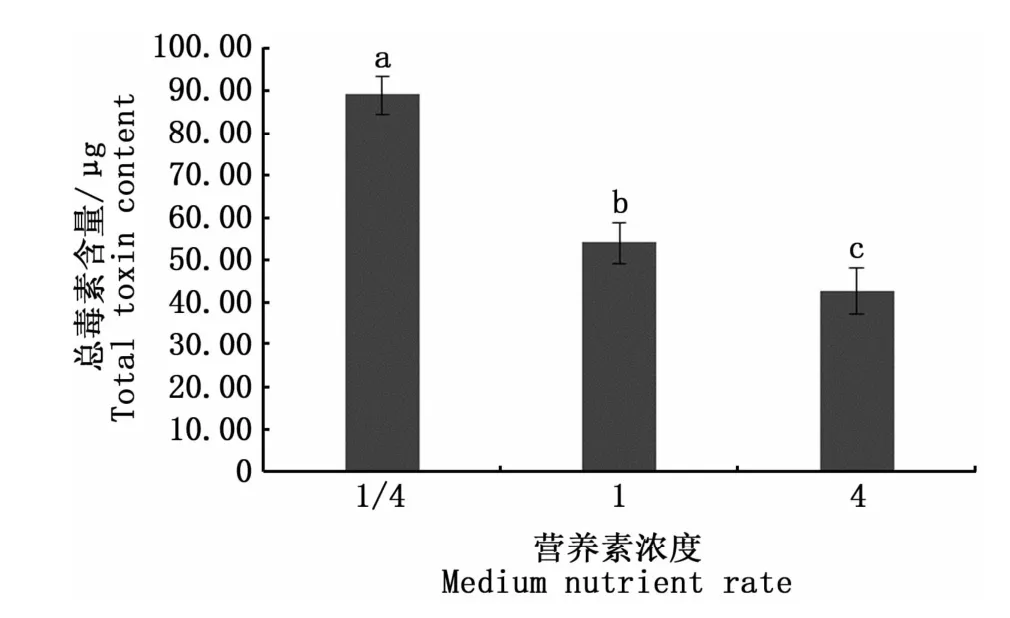
注:不同小寫字母表示有顯著差異(P<0.05);總毒素含量由6.0×106個藻細胞計算得出Note:Different lowercase letters denote significant difference(P<0.05);total toxin content is calculated from 6.0×106 algal cells
3 討論
3.1 塔瑪亞歷山大藻產毒能力
塔瑪亞歷山大藻是所有能夠產生PSP的甲藻中分布最廣泛、認知度最高的物種之一[14],而不同地區的藻株產生的PSP成分不盡相同[15,22]。在本研究中,由于受到標準品數量的限制,該塔瑪亞歷山大藻種共發現有13種PSP組分,這與徐金濤等[11]的研究結果基本一致。DEEDS等[3]、WIESE等[23]以及TAN和RANSANGAN[1]的研究發現,塔瑪亞歷山大藻中有C4和GTX6存在,這說明該藻種的產毒能力非常強,目前來看至少能夠產生15種毒素。本研究采用的塔瑪亞歷山大藻的毒素成分主要為GTX1、GTX4、GTX2、C1、C2和NEO,其中GTX1含量更是高達40%,并且當藻細胞數高于4.0×103個時即可用于定量分析,表明GTX1的含量直接決定了該藻株毒性的強弱。由此看來,塔瑪亞歷山大藻種可用于PSP的基礎研究,亦是分離提取GTX1的最佳理論藻株。
3.2 環境脅迫下塔瑪亞歷山大藻產毒能力差異
在海洋中,單細胞藻類用于生長與代謝的空間資源是有限的,在資源限制下,不僅存在種間競爭,也存在種內競爭,而有害藻類的生理條件可隨著時間和空間變化而變化[24-25]。在資源有限的環境中,塔瑪亞歷山大藻處于種間競爭劣勢[26],生長及產毒也必然受到抑制;而當環境中僅存在塔瑪亞歷山大藻時,因為沒有其他物種的競爭壓力,種內競爭趨勢逐漸顯現。在本研究中,塔瑪亞歷山大藻的總毒性與生存空間成正相關,這說明生存空間的增加顯然更有利于塔瑪亞歷山大藻中PSP的合成。較大的生存空間和較低的繁殖速率保證了更多的PSP合成所需前體物質(如精氨酸)的積累[27],同時相對較低的細胞密度意味著單位藻細胞能夠利用相對多的營養素,這種協同效應最終促進了塔瑪亞歷山大藻的產毒能力差異性的形成。
鹽度主要是通過調節細胞與外界的離子交換速率來影響塔瑪亞歷山大藻的產毒能力[28]。塔瑪亞歷山大藻具有廣鹽性的特征,本實驗結果顯示,該藻株在鹽度15~35均能夠長時間存活,這主要體現在對外界滲透壓的適應性[29]。環境鹽度越低,細胞內部與外界的濃度差越大,細胞需要消耗更多的能量以維持滲透平衡,這導致毒素合成速率較低;而外界環境過高的滲透壓則會使藻細胞更易丟失水分,不利于塔瑪亞歷山大藻的生長及毒素的合成[30]。在本研究中,塔瑪亞歷山大藻產毒能力在鹽度為25時達到峰值,葉志林等[8]發現塔瑪亞歷山大藻的產毒能力隨著鹽度增加而提高,在鹽度35時達到最高;而LIM和OGATA[31]的研究表明,塔瑪亞歷山大藻在鹽度25時產毒能力最低。引起這種差異的原因可能是藻株生長的水域不同,環境的差異加上長時間的適應,使得塔瑪亞歷山大藻對其所處海域的鹽度最為適應,其產毒能力也最高[15]。研究還發現G.catenatum和A.peruvianum毒素含量均在鹽度30時達到最高[15],本實驗結果與其相似。另外,環境鹽度的差異性也會改變塔瑪亞歷山大藻中GTX的組成,隨著鹽度的上升,毒性強的組分百分比含量增加,如GTX4含量的增加以及GTX5含量的減少[28]。
對于塔瑪亞歷山大藻,營養素提供了生長所必需的養分,氮元素決定了PSP的合成。有研究表明[22,28],在氮元素不足時,塔瑪亞歷山大藻的PSP含量和毒性均較低,但本研究中低營養素濃度組中,氮元素含量仍處于相對較高狀態,與上述現象不同。在本研究中,營養素濃度對塔瑪亞歷山大藻產毒能力存在顯著影響,營養素濃度越高,產毒能力越弱。推測當環境中營養素含量較低時,塔瑪亞歷山大藻的種內競爭增加,而適當的競爭有利于個體的生長和代謝。攝入的能量更多地用于增加自身在環境中的競爭力,涉及細胞分裂的營養需求減弱,細胞生化反應增加,更多的精氨酸可用于PSP的合成[32]。WANG和HSIEH[28]發現,增加培養液硝酸鹽濃度,塔瑪亞歷山大藻中GTX1含量比例下降,而GTX5含量比例上升,過低或過高的磷酸鹽濃度均會抑制塔瑪亞歷山大藻產毒;也有研究[28]發現,氮元素的限制有利于GTX1、GTX4的合成,而磷元素的限制有利于GTX2、GTX3的合成。這說明在一定范圍內,低營養環境中,塔瑪亞歷山大藻高毒性成分含量較高,而低毒性成分含量較低,這使得該藻株在營養限制下仍有較高的毒性。此外,筆者在預實驗中發現,當營養素濃度低于1/4倍時,塔瑪亞歷山大藻生長速度逐漸降低,這并不利于毒素的提取與分析。
3.3 海洋中高PSP毒性的有害藻華暴發風險性
海洋生態系統具有自我調節能力,可維持水體環境在一定的范圍內波動。而當氣候驟變或產生人為干預,外力因素超出了海洋自我調節的可承受范圍時,水體環境會發生較大改變,這將對海洋藻類產生巨大影響[33]。一般情況下,海洋環境并不適宜塔瑪亞歷山大藻大量繁殖,水體中僅可能存在微量的營養細胞,亦或是以孢囊的形式存在于底泥中[34]。我國是貝類養殖大國,養殖區遍布沿海各處,水體交換優良,利于產毒藻的遷移和養分的補充,且我國近海海水鹽度基本處于25~35,可見我國海洋環境皆具備滿足塔瑪亞歷山大藻高產毒能力的條件,這提示了高PSP毒性藻類增殖的風險,且該風險在水體富營養化時顯著增加。
本研究結果表明,具有高PSP毒性的有害藻華可能發生在鹽度高、空間充足、富營養化程度較高的海洋環境中。因此,為降低高PSP毒性的有害藻華暴發的風險,應加大符合以上條件海域的監管力度。
