藥用植物根際微生物對其品質形成的影響及其作用機制的研究進展
2021-07-15 11:34:24張振宇段金廒
中草藥 2021年13期
關鍵詞:植物
祝 蕾,嚴 輝,劉 培,張振宇,張 森,郭 盛,江 曙,段金廒
藥用植物根際微生物對其品質形成的影響及其作用機制的研究進展
祝 蕾,嚴 輝*,劉 培,張振宇,張 森,郭 盛,江 曙,段金廒*
南京中醫藥大學 中藥資源產業化與方劑創新藥物國家地方聯合工程研究中心,江蘇省中藥資源產業化過程協同創新中心,國家中醫藥管理局中藥資源循環利用重點研究室,江蘇 南京 210023
根際是植物-土壤-微生物信息和物質交換的重要場所,是影響植物生長和抗逆性的重要因素。植物根系和根際微生物之間的相互作用對于植物的生長和品質形成至關重要。植物根系通過分泌有機酸、糖類、次生代謝物等物質特異性的促進有益微生物的生長,也能通過自身免疫系統和分泌抗生素抑制病原菌的增殖。根際微生物可以通過多種方式參與藥用植物的生長發育、代謝過程以及活性成分的積累,對于藥用植物的養分吸收和利用、土傳病害防治、非生物脅迫應激等方面具有至關重要的作用。主要對根際微生物對藥用植物品質形成的影響及其作用機制的研究進展進行綜述,以期為深入了解根際微生物與藥用植物間的互作關系提供參考。
根際;土壤;微生物;藥用植物;品質形成;作用機制
中醫藥在我國已經有幾千年的發展歷史,歷史上曾為抗擊疾病做出過重要貢獻,當前中醫藥又在新型冠狀病毒肺炎防控中發揮了獨特作用,在國際上引起了廣泛關注。中藥資源是我國重要的戰略性資源,普遍應用于藥物、保健品、食品、其他衛生產品以及出口貿易[1]。中藥材的品質受到藥用植物品種、土壤、氣候因子、農藝措施、采收加工貯藏方式等因素的影響[2]。根際是指受根系活動緊密影響的,物理、生物和化學特性上與周圍大塊土壤不同的靠近植物根系的土壤微域[3]。根際微生物被譽為植物的第2套基因組,對植物的養分循環、生長發育、產量提升、病蟲害防御等起著關鍵作用[4]。近年來,有關根際微生物影響藥用植物生長與健康的研究多集中于藥用植物產量提升、活性成分積累、連作障礙緩解、生防微生物篩選等方面。本文主要對根際微生物對藥用植物品質形成的影響及其作用機制的研究進展進行綜述,以期為深入了解根際微生物與藥用植物間的互作關系提供參考。
1 藥用植物根際微生物多樣性
根際是地球上物質和能量循環、信息交換最活躍的界面之一,也是植物-微生物-土壤三者相互作用密切的一個復雜的生態系統。細菌中變形菌(尤其是假單胞菌科(Pseudomonadaceae)和伯克氏菌科(Burkholderiaceae)是藥用植物根際中最為豐富的優勢菌群[5]。部分真菌如鐮刀菌屬sp.和絲核菌屬sp.是藥用植物重要的病原微生物,而白粉寄生菌屬sp.、盾殼霉屬sp.及木霉屬sp.等則是根際促生真菌,是生防微生物篩選的重要菌種來源[5]。放線菌(尤其是鏈霉菌sp.)能產生多種具有生物活性的代謝物,對于一些土傳病蟲害有較好的拮抗作用[6]。如Mojicevic等[7]從3種民族藥用植物虞美人L.、母菊L.和異株蕁麻L.的根際土壤中分離了103株產孢菌,鑒定屬于鏈霉菌屬,其中有43個鏈霉菌菌株具有產生抗真菌化合物的能力。
2 根際微生物對藥用植物品質形成的影響
根際微生物可以通過直接或間接的方式影響植物的生長和抗逆性。對植物生長有益的土壤微生物通常被稱為植物根際促生菌,主要包括根瘤菌屬、假單胞菌屬、芽孢桿菌屬、菌根真菌、木霉菌屬等。
2.1 根際微生物增強藥用植物的抗病蟲害能力
我國中藥材種類繁多、藥用部位多樣、產區跨度大、生物學特征差異明顯,從而導致病蟲害種類多樣性高且危害嚴重[8]。多年生中藥材(如人參、西洋參、三七等)地下病蟲害尤其普遍,常見的主要有根腐病、黑斑病、根結線蟲病、白粉病、圓斑病等[9]。Zohair等[10]從3種藥用植物根際分離出的2株真菌MG772677和蓋姆斯木霉KX685665,對4種病原菌(包括腐皮鐮孢菌、立枯絲核菌、齊整小核菌和大麗輪枝菌)的平均抑制率分別為77.90%、77.98%。Nakaew等[11]研究發現從姜黃根際土壤中分離到的鏈霉菌sp. TM32對植物病原菌具有較強的抗菌活性。Kim等[12]研究發現從人參根際分離出的解淀粉芽孢桿菌AK-0能夠抑制由人參銹腐病菌引起的根腐病。Li等[13]研究發現灰黃青霉菌CF3能夠顯著抑制附子中土傳根病原菌尖孢鐮刀菌和齊整小核菌的生長。
2.2 根際微生物增強藥用植物的抗逆能力
低溫、高溫、干旱、鹽堿、水澇、重金屬等不利因子會擾亂植物的生理生化狀態,影響植物的生長發育,嚴重的會導致植株死亡。研究表明,根際微生物通過調控植物體內與脅迫相關基因的表達,增強植物在脅迫條件下的養分吸收,提高植物的應急耐受性[14]。Egamberdieva等[15]研究發現共接種中慢生根瘤菌sp. NWXJ19或NWXJ31和極端東方化假單胞菌TSAU20顯著緩解了甘草植株的鹽脅迫,使甘草的產量和根瘤數增加。Yousefi等[16]研究發現接種產脂固氮螺菌或圓褐固氮菌能顯著提高車桑子種子的耐鹽度,使種子的發芽率以及根、莖的長度和生物量增加。菌根真菌提高了干旱條件下小茴香的滲透調節能力,使小茴香揮發油產量、葉片和果實的養分含量增加[17]。菌根真菌還能顯著提高鉛脅迫下龍葵的根際微生物多樣性和對氨基酸、糖類、脂肪酸等的代謝活性[18]。此外,研究表明重金屬如Co、Cd和Zn的存在會促進根際微生物分泌鐵載體。如Zn2+脅迫下,人參根際真菌哈茨木霉、圓紅冬孢酵母、尖孢鐮刀菌會增加鐵載體的分泌,以應對環境的威脅[19]。
2.3 根際微生物促進藥用植物的生長和藥用部位發育
根際有益微生物可以促進植物對氮、磷、鉀以及鐵等礦質元素的吸收直接促進植物的生長。具有固氮作用的根際微生物主要包括根瘤菌屬、緩生根瘤菌屬、固氮螺菌屬、菌根真菌等,芽孢桿菌、假單胞桿菌、沙雷氏菌、青霉菌、鏈霉菌、菌根真菌等被具有解磷作用[20]。張海珠等[21]研究發現,菌根真菌提高了滇重樓根際土壤中氮、磷、鉀等營養元素的含量,使滇重樓藥材對Mg、Na、Zn、Ni的富集能力增加,有利于其生長和品質形成。韓麗珍等[22]從茶樹根際分離了2株溶磷細菌貝萊斯芽孢桿菌和堅強芽孢桿菌,通過產生有機酸來溶解磷元素,顯著促進花生的生長。部分根際微生物還能產生具有促生作用的植物激素。王琦琦等[23]、Chandra等[24]和費詩萱等[25]分別從木堿蓬、甜葉菊、紅棗根際中分離得到具有產吲哚-3-乙酸活性的根際細菌,具有增強植株的根系生長,提高株高以及根和莖生物量等作用。此外,根際有益微生物還可以通過誘導宿主的系統抗性達到拮抗病原菌和促進植株生長的目的。Berendsen等[26]研究發現擬南芥根際的微桿菌sp.、寡養單胞菌sp.和黃單胞菌sp.共同參與了誘導擬南芥對霜霉病的系統抗性。
2.4 根際微生物促進藥用植物有效成分的合成
植物的次生代謝在信息交換、環境適應和協同進化中起著重要的作用,并且能吸引傳粉者和傳種者,防御天敵和病原體,還能參與植物之間的協同和競爭作用,有助于植物對物理化學環境改變的適應等[27]。植物的次生代謝產物不僅是植物長期適應進化的結果,還常常作為一種藥物植的有效成分而存在。
根際微生物通過調控藥用植物中與次生代謝產物合成相關基因的表達,合成轉化植物活性成分前體的關鍵酶等機制促進藥用植物有效成分的積累[28-29]。Zeng等[30]研究發現板藍根際顯著富集的伯克霍爾德菌sp.參與板藍功效物質靛藍的合成。Zhai等[31]研究發現棘孢木霉ACCC30536通過上調青蒿素生物合成關鍵酶基因等的表達,提高了黃花蒿葉片中青蒿素濃度。接種混合的根際促生菌增加了栝樓中藥用成分多糖和天花粉蛋白的積累[32]。短小芽孢桿菌通過增加關鍵酶的表達來提高甘草酸的含量[33]。圓褐固氮菌和巴西固氮螺菌共接種能使唇萼薄荷中脫落酸、可溶性糖、蛋白質、酚類、黃酮類以及含氧單萜的含量提高[34]。假單胞菌和泛菌sp.可能通過產生植物激素和多糖參與丹參次生代謝產物的合成,對丹參中酚類物質迷迭香酸、丹酚酸B的積累有顯著促進[35]。
2.5 根際微生物緩解藥用植物連作障礙
我國中藥材栽培歷史悠久,本草中記載有關藥用植物栽培的方法可追溯至2600多年以前[36]。目前已經實現了300多種藥用植物的人工種植,大約有70%的根及根莖類藥材受到連作障礙的困擾[37]。連作障礙的形成是土壤養分缺乏和理化性質改變、根系分泌物和根殘留物引起的化感自毒作用、土壤傳染性病蟲害加劇等多種因子相互作用的結果[38]。研究表明尖孢鐮刀菌能刺激植株內酚酸物質的積累,導致植株程序性死亡[39]。酚酸類物質能差異性地調節根際土壤微生物群落,促進致病菌的增殖同時使有益菌衰減[40]。酚酸與土壤致病菌的正相互作用,可能是連作障礙形成的主要機制。
目前解決連作障礙的措施主要包括選育抗連作障礙的藥用植物品種、建立合理的耕作制度、施用有機肥和微生物菌肥[41]。在農業生產中,間套作和輪作制度是一種傳統有效的減輕或避免連作障礙的種植模式[42]。施用添加木霉的有機肥使黃瓜連作土壤更接近初始土壤,影響了其根際微生物的組成,提高了真菌的多樣性,使植株生長狀態提升[43]。根際微生物能夠改善土壤微環境,增加根際微生物的生物量和土壤酶活性,降低土壤中真菌/細菌的比例,緩解連作障礙。如施用有益微生物復合菌劑能夠緩解太子參[44]、三七[45]、廣藿香[46]、滇重樓[47]等中藥材的連作障礙。Zhao等[48]研究發現三七-玉米體系中土壤微生物酶活性顯著高于三七單作體系,細菌和真菌數量增多,但真菌-細菌的比例減小,在屬水平上鐮刀菌和不動桿菌sp的相對豐度顯著減少,而芽孢桿菌相對豐度顯著增加。Tian等[49]從人參根際分離到1株解淀粉芽孢桿菌TB6,接種后通過降低根際真菌的豐富度和多樣性,提高多酚氧化酶、磷酸酶、脲酶和過氧化氫酶的活性,促進人參根系的生長。
3 藥用植物調控根際微生物組成的分子機制
植物在塑造根際微生物群落方面起著至關重要的作用。植物根系分泌物、根系代謝、根系角質層成分和植物免疫系統是植物調控根際微生物的組成的主要途徑[50],見圖1。
3.1 藥用植物的根系分泌物調控根際微生物
植物在其生長過程中不斷的向根際分泌包括糖類、氨基酸、有機酸、脂肪酸、酚酸、甾醇、蛋白質以及生長因子在內的各種物質[53]。根系分泌物能夠調控根際微生物與根際趨化、信號響應、次生代謝、生物膜形成等基因的表達,促進微生物在根際的定殖和生長。研究表明,根際細菌優先利用植物分泌的芳香有機酸如煙堿、莽草酸、水楊酸、肉桂酸和吲哚-3-乙酸[54]。如水楊酸可以調節特定的細菌類群在根際的定殖[55],擬南芥中三萜生物合成基因簇的代謝產物能調節擬南芥根際中特定細菌類群的生長[56]。根系分泌物中的信號分子是調節根際微生物在根際定殖的關鍵因子。根瘤菌和叢枝菌根真菌是植物與根際微生物共生的2種主要形式。類黃酮是根瘤菌與豆科植物形成共生固氮的信號分子,通過激活根瘤菌基因的表達,誘導根瘤形成[57]。菌根真菌可與80%以上的高等植物根系形成共生菌根,植物根系釋放的獨腳金內酯作為信號分子,誘導菌根真菌菌絲分枝并侵染植物形成菌根[58]。除了這2種共生形式,蘭科植物天麻在人工栽培過程中需要小菇屬萌發菌石斛小菇Fan et Guo、紫萁小菇Lange、開唇蘭小菇Fan et Guo、蘭小菇Fan et Guo等才能完成種子的萌發[59]。Carvalhais等[60]研究發現植物合成獨腳金內酯的能力對根際真菌群落有明顯的影響,野生型擬南芥與獨腳金內酯合成受損的突變體相比根際真菌群落的β-多樣性存在顯著差異,黑附球菌、青霉菌sp.、和等對獨腳金內酯有趨化作用。
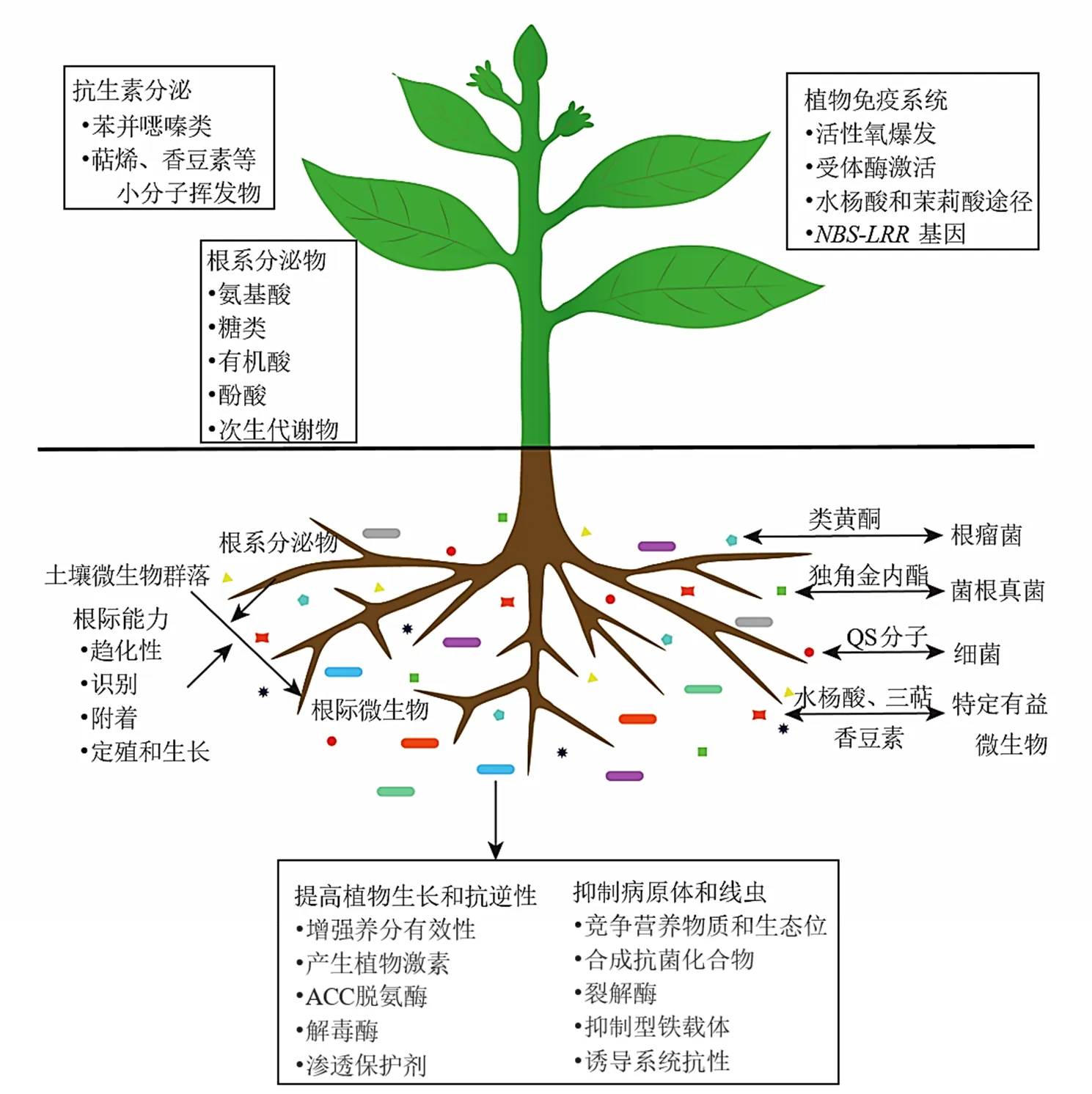
該圖在Trivedi等[51]和Huang等[52]繪制基礎上修改而來。不同顏色的矩形代表不同的根際微生物類群。三角形、正方形、五邊形、圓形和星形代表植物根系釋放的不同種類的化合物
3.2 藥用植物分泌抗生素
3.3 藥用植物的自身免疫
植物自身的免疫系統也可以調節根際微生物的定殖。植物的免疫系統主要由病原相關分子模式觸發的免疫反應(PAMP-triggered immunity,PTI)和效應因子觸發的免疫反應(effector-triggered immunity,ETI)組成。PTI能夠識別病原體的鞭毛蛋白、脂多糖、幾丁質和延伸因子Tu等,通過活性氧爆發、絲裂原活化蛋白激酶的激活以及水楊酸和茉莉酸信號途徑的誘導產生防御應答[51]。ETI則可以識別微生物分泌的效應蛋白,誘導過敏反應,使被侵染部位的植物細胞死亡,阻止病原體的進一步擴散。當植物受到病原體侵害時,會啟動自身免疫防御反應,間接對根際微生物的組成產生影響[53]。植物免疫激素會因不同病原菌而產生響應,進而相應的調控植物防御信號的產生。水楊酸途徑主要介導由病原微生物引起的非特異性的系統獲得性抗性[67],而茉莉酸和乙烯則介導由特定的根定殖根際細菌引起的誘導性系統抗性[68]。水楊酸處理可以增強水稻和二穗短柄草(L.) P. Beauv對土傳真菌紋枯病菌的抗性,而水楊酸水平降低的轉基因水稻植株則更容易被紋枯病菌感染[69]。乙烯響應因子(ethylene response factors,)基因缺失的突變體與野生型番茄相比,根系分泌物組成發生了顯著變化,基因的敲除導致根系分泌物對南方根結線蟲和馬鈴薯白線蟲的趨化性增強[70]。
4 根際微生物增強藥用植物生長和適應性的分子機制
根際微生物可以通過直接或間接的方式影響植物的生長和抗逆性[71]。一方面,根際微生物可以增強植物對養分的吸收利用,產生促進植物生長的化合物如植物激素;另一方面還可以通過直接競爭效應和產生抑菌物質幫助植物抵御各種生物和非生物脅迫(圖1)。
4.1 根際微生物增強藥用植物的養分有效性
根際微生物影響藥用植物營養狀況的機制主要包括向植物提供養分、增強養分的生物利用度以及增強植物對養分的獲取能力[72]。根瘤菌可以與豆科植物形成共生固氮體,在非豆科植物桔梗[73]、甘草[74]、纈草[75]等中也發現了根瘤菌的存在。菌根真菌可與大多數陸生高等植物形成共生菌根,通過根部的菌絲網絡增大與土壤的接觸面積,增強藥用植物對氮、磷的吸收[58]。有些根際微生物如根瘤菌、假單胞菌、木霉菌分泌的鐵載體可以有效地與鐵結合釋放出鐵離子被藥用植物吸收[76]。在牡丹根際發現的黑曲霉具有溶解含鈣、鐵、鋅、鋁等的難溶磷酸鹽的能力[77]。Hussein等[19]從人參根際分離得到具有高效產鐵載體能力的青霉菌sp. JJHO。此外,根際微生物還可以通過產生植物激素(如生長素、細胞分裂素、赤霉素、脫落酸、乙烯、油菜素甾醇、獨腳金內酯等)促進根的增殖,使根的總表面積增加,有利于藥用植物對土壤中水和養分的吸收,對藥用植物的生長和活性成分的積累有促進作用。大多數根際促生菌中都發現有生長素的產生,如桔梗[73]根際的根瘤菌,甜葉菊[24]、人參[78]和紅棗[25]的根際促生菌均能產生生長素。此外赤霉菌具有產生赤霉素的能力。研究表明,伯克氏菌可參與板藍根中功效物質靛藍的合成[30],木霉菌通過上調黃花蒿中青蒿素生物合成關鍵酶的表達促進了青蒿素的合成[31],菌根真菌通過上調類黃酮生物合成基因的表達增加了金線蓮中類黃酮的含量[79]。
4.2 根際微生物增強藥用植物的抗性
在干旱、高鹽度、水澇、重金屬污染、低溫、高溫等非生物脅迫下,根際微生物產生1-氨基環丙烷-1-羧酸(1-amino-cyclopropanecarboxylic acid,ACC)脫氨酶調節乙烯合成前體ACC的濃度,降低乙烯的產量,緩解乙烯介導的植物生長抑制對各種脅迫的響應。如芽孢桿菌通過調節ACC的積累、ACC-氧化酶和ACC-合酶的活性,改善了鹽脅迫下玉米植株的生長[80]。酶或非酶抗氧化劑活性的增強是根際微生物應對各種脅迫的主要機制之一。印度梨形孢和圓褐固氮菌共接種降低了黃花蒿植株丙二醛和H2O2的濃度,增強了抗氧化酶(超氧化物歧化酶、過氧化氫酶、抗壞血酸過氧化物酶和谷胱甘肽還原酶)和非酶抗氧化劑(總黃酮、酚類和類胡蘿卜素)的活性,并使青蒿素、脯氨酸的含量顯著提高[81]。共接種混合的根際促生菌提高了姜黃對1,1-二苯基-2-三硝基苯肼自由基和2,2′-聯氮-雙-(3-乙基苯并噻唑啉-6-磺酸)二銨鹽自由基的清除能力以及酚類和總黃酮的含量[82]。接種根際促生菌還對曼陀羅葉和根中的氨基酸代謝有顯著影響[83]。Amanifar等[84]發現菌根真菌能增加甘草中磷鉀的濃度,促進脯氨酸的積累,調節植株中Na+/K+的穩態以增強甘草對鹽的耐受。此外,根際微生物還能夠上調植物相關抗性基因的表達,如枯草芽孢桿菌的應用增強了擬南芥中抗干旱和耐鹽相關基因的表達[85]。根際微生物還能通過對重金屬進行吸附、沉淀或者產生金屬螯合物,減少植物對重金屬的吸收利用緩解重金屬脅迫[86]。芽孢桿菌和根瘤菌共接種能降低紫花苜蓿植株中銅的積累,增加土壤脲酶、蔗糖酶和β-葡萄糖苷酶活性[87]。
4.3 根際微生物拮抗藥用植物病原微生物
根際微生物對病原菌的拮抗主要通過抗生作用、對生態位和營養元素的競爭、干擾群效應、重寄生作用和誘導植物抗病性等機制[4]。有些根際微生物如青霉菌、鏈霉菌、放線菌通過產生抑菌物質抑制病原體的生長。人參根際的解淀粉芽孢桿菌對人參灰霉病菌Pers.[88]、人參銹腐病菌[12]具有顯著的抑制活性,其中人參根際的解淀粉芽孢桿菌AK-0以及姜黃[11]根際的鏈霉菌sp. TM32通過合成抗菌素抑制多種植物病原菌的生長。還有一些根際微生物如假單胞菌、芽孢桿菌能產生揮發性有機化合物,研究表明醛類化合物是發揮抑菌作用的關鍵物質[89]。從根際土壤中分離的枯草芽孢桿菌通過產生苯甲醛、壬醛、苯并噻唑和苯乙酮等揮發性有機化合物抑制馬鈴薯環腐病菌ssp.的生長[90]。鐵元素是根際微生物與病原菌爭奪激烈的稀缺資源,某些微生物產生的抑制型鐵載體不能被病原菌識別,在根際鐵元素競爭中處于優勢,從而抑制病原菌生長[91]。Sheng等[92]從半夏根際土壤中分離的產鐵載體短短芽孢桿菌GZDF3對白色念珠菌有顯著拮抗活性。40%~50%細菌通過群體感應調節微生物-微生物相互作用。根際微生物通過產生裂解酶(如內酯酶、酰胺酶、還原酶、氧化酶等)破壞細菌群體感應系統,對病原體生物膜的形成、毒力因子的合成和分泌等行為產生干擾[93]。芽孢桿菌和貪噬菌通過降解丁香假單胞菌pv.產生的群體感應信號分子-十四烷酰基高絲氨酸內酯和-己酰基高絲氨酸內酯緩解番茄植株的過敏反應[94]。此外,微生物的分泌系統、鞭毛和菌毛等細胞結構,以及效應蛋白等蛋白質通過激發植物的誘導性系統抗性,間接促進植物防御[51]。如芽孢桿菌屬的植物促生根瘤菌通過激活水楊酸、茉莉酸或乙烯信號通路,觸發擬南芥、煙草、番茄等植物的誘導性系統抗性[95]。
5 結語與展望
植物與根際微生物之間存在著復雜的相互作用關系。根際具有極其豐富的微生物多樣性,這些微生物可以直接或間接的影響藥用植物的生長發育和品質形成。根際微生物能夠影響植物的光合作用、呼吸代謝等生理過程,進而對藥用植物的生長發育及根系功能產生影響,還可以通過改變藥用植物根系分泌物的組成,改善土壤理化性質,緩解藥用植物連作障礙;一些根際微生物被報道能夠參與藥用成分的合成。某些根際微生物自身可以產生抗菌代謝物,如2,4-二乙酰間苯三酚、吩嗪、脂肽以及揮發性有機化合物等。因此,有效的利用根際微生物的促生機制以及對植物病蟲害的生物防治作用,對提高植物的產量和品質具有重要的意義。公認的道地性形成的理論主要包括環境生態論、品種延續與產地變遷論、品質生態學理論、逆境效應理論、本草基因組學理論。此外,何冬梅等[29]提出了中藥微生態與中藥道地性的理論,從中藥微生態的角度闡釋了中藥道地性的形成原因。目前藥用植物根際微生物研究的重點領域是藥用植物連作障礙,已經在三七、地黃、人參、西洋參等中藥材中取得了重要突破。其次是對于中藥材道地性的研究,多集中于根際微生物對道地藥材形成的物質基礎即次生代謝產物的影響,但對于作用機制的研究還比較少。
隨著各種新興技術如現代分子生物技術、人工智能技術、信息科學技術以及各種觀測技術的發展和完善,未來對根際微生物與藥用植物互作機制的研究會不斷的深入。今后藥用植物根際微生物的研究重點應該包括以下幾個方面:(1)加強對根際促生菌促生機制的研究,加快生防菌劑的開發。多年生藥用植物地下病蟲害普遍且危害嚴重,導致中藥材品質的下降和對環境的污染,微生物菌劑具有高效、安全、無公害等優點,能夠改善土壤環境,對于提升藥用植物的產量和質量具有關鍵作用;(2)加強對藥用植物連作障礙機制的研究。中藥材需要一定的種植年限才能達到藥用標準,長期在同一土地上連代種植導致連作障礙普遍存在。關于連作障礙形成的具體機制還不清楚。闡明藥用植物連作障礙的內在機制,為藥用植物的種植提供理論指導,對于中藥材品質提升和中藥資源可持續發展具有重要的意義;(3)加強對根際微生態介導的中藥材道地性形成機制的研究。特定的生態地理環境造就了道地藥材優良的品質和臨床療效,研究根際微生態與道地藥材品質間的關聯,將微觀的藥用植物代謝產物變化,根際微生物組成和宏觀上土壤理化性質,植物生長特性統一結合、系統研究,有助于全面地闡釋藥用植物的生長調控機制,從而闡明道地藥材的形成機制。
利益沖突 所有作者均聲明不存在利益沖突
[1] 馬曉晶, 郭娟, 唐金富, 等. 論中藥資源可持續發展的現狀與未來 [J]. 中國中藥雜志, 2015, 40(10): 1887-1892.
[2] 楊毅, 田侃, 倪新興, 等. 中藥材品質影響因素實證研究 [J]. 中藥材, 2016, 39(6): 1251-1256.
[3] Garcia J, Kao-Kniffin J. Microbial group dynamics in plant rhizospheres and their implications on nutrient cycling [J]., 2018, 9: 1516.
[4] Mendes R, Garbeva P, Raaijmakers J M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms [J]., 2013, 37(5): 634-663.
[5] Schlaeppi K, Bulgarelli D. The plant microbiome at work [J]., 2015, 28(3): 212-217.
[6] 肖艷紅, 李菁, 劉祝祥, 等. 藥用植物根際微生物研究進展 [J]. 中草藥, 2013, 44(4): 497-504.
[7] Mojicevic M, D'Agostino P M, Nikodinovic-Runic J,. Antifungal potential of bacterial rhizosphere isolates associated with three ethno-medicinal plants (poppy, chamomile, and nettle) [J]., 2019, 22(3): 343-353.
[8] 沈亮, 徐江, 陳士林, 等. 無公害中藥材病蟲害防治技術探討 [J]. 中國現代中藥, 2018, 20(9): 1039-1048.
[9] 王瑞, 董林林, 徐江, 等. 農田栽參模式中人參根腐病原菌鑒定與防治 [J]. 中國中藥雜志, 2016, 41(10): 1787-1791.
[10] Zohair M M, El-Beih A A, Sadik M W,. Promising biocontrol agents isolated from medicinal plants rhizosphere against root-rot fungi [J]., 2018, 15: 11-18.
[11] Nakaew N, Lumyong S, Sloan W T,. Bioactivities and genome insights of a thermotolerant antibiotics-producing Streptomyces sp. TM32 reveal its potentials for novel drug discovery [J]., 2019, 8(11): e842.
[12] Kim Y S, Balaraju K, Jeon Y H. Biological characteristics ofAK-0 and suppression of ginseng root rot caused by[J]., 2017, 122(1): 166-179.
[13] Li Y, Guo Q, Wei X,. Biocontrol effects ofagainst monkshood (Debx.) root diseases caused byandspp [J]., 2019, 127(5): 1532-1545.
[14] 王希付, 張雪, 趙榮華, 等. 叢枝菌根真菌在藥用植物中的作用及機制研究進展 [J]. 中國實驗方劑學雜志, 2020, 26(11): 217-226.
[15] Egamberdieva D, Li L, Lindstr?m K,. A synergistic interaction between salt-tolerantandstrains improves growth and symbiotic performance of liquorice (Fish.) under salt stress [J]., 2016, 100(6): 2829-2841.
[16] Yousefi S, Kartoolinejad D, Bahmani M,. Effect ofandon germination and early growth of hopbush shrub (L.) under salinity stress [J]., 2017, 36(2): 107-120.
[17] Zardak S G, Dehnavi M M, Salehi A,. Effects of using arbuscular mycorrhizal fungi to alleviate drought stress on the physiological traits and essential oil yield of fennel [J]., 2018, 6: 31-38.
[18] 韓娟, 張向前, 趙金莉. 鉛脅迫下接種AM真菌對龍葵根際土壤微生物群落代謝特征的影響 [J]. 中國生態農業學報: 中英文, 2019, 27(4): 545-553.
[19] Hussein K A, Joo J H. Zinc ions affect siderophore production by fungi isolated from therhizosphere [J]., 2019, 29(1): 105-113.
[20] 童琳, 唐旭利, 張靜, 等. 菌根形成對不同成熟度的森林優勢樹種磷吸收的影響 [J]. 生態科學, 2015, 34(4): 93-98.
[21] 張海珠, 李楊, 張彥如, 等. 菌根真菌處理下滇重樓對營養元素的吸收和積累 [J]. 環境化學, 2019, 38(3): 615-625.
[22] 韓麗珍, 周靜, 王歡. 兩株對花生促生的芽孢桿菌的鑒定及溶磷特性研究 [J]. 基因組學與應用生物學, 2019, 38(9): 4066-4076.
[23] 王琦琦, 馮麗, 李楊, 等. 新疆木堿蓬()根際耐鹽促生細菌的篩選及鑒定 [J]. 微生物學通報, 2019, 46(10): 2569-2578.
[24] Chandra S, Askari K, Kumari M. Optimization of indole acetic acid production by isolated bacteria fromrhizosphere and its effects on plant growth [J]., 2018, 16(2): 581-586.
[25] 費詩萱, 張敏, 王迎, 等. 具有ACC脫氨酶活性的紅棗根際促生菌株的分離篩選及其促生效果研究 [J]. 西北林學院學報, 2019, 34(6): 140-146.
[26] Berendsen R L, Vismans G, Yu K,. Disease-induced assemblage of a plant-beneficial bacterial consortium [J]., 2018, 12(6): 1496-1507.
[27] 陳曉亞, 王凌健, 毛穎波, 等. 植物萜類生物合成與抗蟲反應 [J]. 生命科學, 2015, 27(7): 813-818.
[28] Zhai X, Jia M, Chen L,. The regulatory mechanism of fungal elicitor-induced secondary metabolite biosynthesis in medical plants [J]., 2017, 43(2): 238-261.
[29] 何冬梅, 王海, 陳金龍, 等. 中藥微生態與中藥道地性 [J]. 中國中藥雜志, 2020, 45(2): 290-302.
[30] Zeng M J, Zhong Y J, Cai S J,. Deciphering the bacterial composition in the rhizosphere of(NeeS) Bremek [J]., 2018, 8(1): 15831.
[31] Zhai T T, Wang Y F, Liu C L,.ACCC30536 inoculation improves soil nutrition and leaf artemisinin production in[J]., 2019, 41(4): 46.
[32] Jiang C H, Xie P, Li K,. Evaluation of root-knot nematode disease control and plant growth promotion potential of biofertilizer Ning shield onin the field [J]., 2018, 49(2): 232-239.
[33] Xie Z C, Chu Y K, Zhang W J,.alleviates drought stress and increases metabolite accumulation inFisch [J]., 2019, 158: 99-106.
[34] Asghari B, Khademian R, Sedaghati B. Plant growth promoting rhizobacteria (PGPR) confer drought resistance and stimulate biosynthesis of secondary metabolites in pennyroyal (L.) under water shortage condition [J]., 2020, 263: 109132.
[35] You H, Yang S J, Zhang L,. Promotion of phenolic compounds production inhairy roots by six strains of rhizosphere bacteria [J]., 2018, 18(3): 160-168.
[36] 郭巧生, 王長林. 我國藥用植物栽培歷史概況與展望 [J]. 中國中藥雜志, 2015, 40(17): 3391-3394.
[37] 張重義, 林文雄. 藥用植物的化感自毒作用與連作障礙 [J]. 中國生態農業學報, 2009, 17(1): 189-196.
[38] Alemán F. Common bean response to tillage intensity and weed control strategies [J]., 2001, 93(3): 556-563.
[39] Qin X J, Wu H M, Chen J,. Transcriptome analysis ofin response to the infection of pathogenic[J]., 2017, 17(1): 155.
[40] Wu H M, Xu J J, Wang J Y,. Insights into the mechanism of proliferation on the special microbes mediated by phenolic acids in therhizosphere under continuous monoculture regimes [J]., 2017, 8: 659.
[41] 周芳, 曹國璠, 李金玲, 等. 藥用植物連作障礙機制及其緩解措施研究進展 [J]. 山地農業生物學報, 2019, 38(3): 67-72.
[42] 王素娜. 輪作緩解甜瓜連作障礙的機理研究 [D]. 沈陽: 沈陽農業大學, 2017.
[43] Cai F, Pang G, Li R X,. Bioorganic fertilizer maintains a more stable soil microbiome than chemical fertilizer for monocropping [J]., 2017, 53(8): 861-872.
[44] 吳林坤, 吳紅淼, 朱銓, 等. 不同改良措施對太子參根際土壤酚酸含量及特異菌群的影響 [J]. 應用生態學報, 2016, 27(11): 3623-3630.
[45] 趙涵予, 位小丫, 林煜, 等. 兩種菌劑處理對連作三七根際土壤的修復效果分析 [J]. 福建農業學報, 2018, 33(11): 1181-1189.
[46] 鄭揚波, 李明, 張梓豪, 等. EM菌對連作廣藿香扦插苗生長特性及土壤微生態的影響 [J]. 華南農業大學學報, 2019, 40(2): 60-64.
[47] 歐洪, 郭冬琴, 林俊杰, 等. AM真菌對滇重樓根際土壤微生物數量及酶活性的影響 [J]. 中藥材, 2016, 39(5): 948-955.
[48] Zhao J, Li Y L, Wang B Y,. Comparative soil microbial communities and activities in adjacent Sanqi ginseng monoculture and maize-Sanqi ginseng systems [J]., 2017, 120: 89-96.
[49] Tian L, Shi S H, Ji L,. Effect of the biocontrol bacteriumon the rhizosphere in ginseng plantings [J]., 2018, 21(3): 153-162.
[50] 葛藝, 徐紹輝, 徐艷. 根際微生物組構建的影響因素研究進展 [J]. 浙江農業學報, 2019, 31(12): 2120-2130.
[51] Trivedi P, Leach J E, Tringe S G,. Plant-microbiome interactions: From community assembly to plant health [J]., 2020, 18(11): 607-621.
[52] Huang X F, Chaparro J M, Reardon K F,. Rhizosphere interactions: Root exudates, microbes, and microbial communities [J]., 2014, 92(4): 267-275.
[53] 吳林坤, 林向民, 林文雄. 根系分泌物介導下植物-土壤-微生物互作關系研究進展與展望 [J]. 植物生態學報, 2014, 38(3): 298-310.
[54] Zhalnina K, Louie K B, Hao Z,. Dynamic root exudate chemistry and microbial substrate preferences drive patterns in rhizosphere microbial community assembly [J]., 2018, 3(4): 470-480.
[55] Lebeis S L, Paredes S H, Lundberg D S,. PLANT MICROBIOME. Salicylic acid modulates colonization of the root microbiome by specific bacterial taxa [J]., 2015, 349(6250): 860-864.
[56] Huang A C, Jiang T, Liu Y X,. A specialized metabolic network selectively modulatesroot microbiota [J]., 2019, 364(6440): eaau6389.
[57] Broughton W J, Perret X. Genealogy of legume-symbioses [J]., 1999, 2(4): 305-311.
[58] Davison J, Moora M, ?pik M,. FUNGAL SYMBIONTS. Global assessment of arbuscular mycorrhizal fungus diversity reveals very low endemism [J]., 2015, 349(6251): 970-973.
[59] 王彩云, 侯俊, 王永, 等. 天麻種子萌發菌研究進展 [J]. 北方園藝, 2017(12): 198-202.
[60] Carvalhais L C, Rincon-Florez V A, Brewer P B,. The ability of plants to produce strigolactones affects rhizosphere community composition of fungi but not bacteria [J]., 2019, 9: 18-26.
[61] Bednarek P, Osbourn A. Plant-microbe interactions: Chemical diversity in plant defense [J]., 2009, 324(5928): 746-748.
[62] Hu L F, Robert C A M, Cadot S,. Root exudate metabolites drive plant-soil feedbacks on growth and defense by shaping the rhizosphere microbiota [J]., 2018, 9(1): 2738.
[63] 張賀, 何依依, 吳家慶, 等. 玉米根系分泌物中關鍵抑菌物質對大豆疫霉的抑菌活性 [J]. 植物保護, 2019, 45(6): 124-130.
[64] 王菲, 孫文帥, 張秀婷, 等. 間作大蔥對桔梗根系分泌物的影響 [J]. 山東農業科學, 2019, 51(11): 68-73.
[65] Lv H, Cao H S, Nawaz M A,. Wheat intercropping enhances the resistance of watermelon towilt [J]., 2018, 9: 696.
[66] Li X G, de Boer W, Zhang Y N,. Suppression of soil-bornepathogens of peanut by intercropping with the medicinal herb[J]., 2018, 116: 120-130.
[67] Fu Z Q, Dong X N. Systemic acquired resistance: Turning local infection into global defense [J]., 2013, 64: 839-863.
[68] Pieterse C M J, van der Does D, Zamioudis C,. Hormonal modulation of plant immunity [J]., 2012, 28: 489-521.
[69] Kouzai Y, Kimura M, Watanabe M,. Salicylic acid-dependent immunity contributes to resistance against, a necrotrophic fungal agent of sheath blight, in rice and[J]., 2018, 217(2): 771-783.
[70] Dyer S, Weir R, Cox D,. Ethylene Response Factor () genes modulate plant root exudate composition and the attraction of plant parasitic nematodes [J]., 2019, 49(13/14): 999-1003.
[71] Mantelin S, Touraine B. Plant growth-promoting bacteria and nitrate availability: Impacts on root development and nitrate uptake [J]., 2004, 55(394): 27-34.
[72] Müller D B, Vogel C, Bai Y,. The plant microbiota: Systems-level insights and perspectives [J]., 2016, 50: 211-234.
[73] Huang C M, Chen W C, Lin S H,. Exploration of root-associated bacteria from the medicinal plant[J]., 2019, 34(4): 413-420.
[74] He C, Wang W, Hou J. Characterization of dark septate endophytic fungi and improve the performance of liquorice under organic residue treatment [J]., 2019, 10: 1364.
[75] Ghodsalavi B, Ahmadzadeh M, Soleimani M,. Isolation and characterization of rhizobacteria and their effects on root extracts of[J]., 2013, 7(3): 338-344.
[76] David S R, Ihiawakrim D, Regis R,. Iron removal from raw asbestos by siderophores-producing[J]., 2020, 385: 121563.
[77] 薛冬, 黃向東, 宋根娣, 等. 牡丹根際溶磷真菌的篩選及其促生效應 [J]. 生態環境學報, 2018, 27(9): 1639-1645.
[78] Ji W X, Leng X, Jin Z X,. Plant growth promoting bacteria increases biomass, effective constituent, and modifies rhizosphere bacterial communities of[J]., 2019, 69(2): 135-146.
[79] Zhang Y, Li Y Y, Guo S X. Effects of the mycorrhizal fungussp. AR2 on growth and flavonoid accumulation in[J]., 2020, 8: e8346.
[80] Misra S, Chauhan P S. ACC deaminase-producing rhizosphere competentspp. mitigate salt stress and promotegrowth by modulating ethylene metabolism [J]., 2020, 10(3): 119.
[81] Arora M, Saxena P, Abdin M Z,. Interaction between Piriformospora indica anddiminish the effect of salt stress inL. by enhancing enzymatic and non-enzymatic antioxidants [J]., 2020, 80(1): 61-73.
[82] Dutta S C, Neog B. Accumulation of secondary metabolites in response to antioxidant activity of turmeric rhizomes co-inoculated with native arbuscular mycorrhizal fungi and plant growth promoting rhizobacteria [J]., 2016, 204: 179-184.
[83] Rahmoune B, Zerrouk I Z, Bouzaa S,. Amino acids profiling inand study of their variations after inoculation with plant growth promoting Rhizobacteria [J]., 2019, 74(10): 1373-1383.
[84] Amanifar S, Khodabandeloo M, Mohseni Fard E,. Alleviation of salt stress and changes in glycyrrhizin accumulation by arbuscular mycorrhiza in liquorice () grown under salinity stress [J]., 2019, 160: 25-34.
[85] Woo O G, Kim H, Kim J S,.strain GOT9 confers enhanced tolerance to drought and salt stresses inand[J]., 2020, 148: 359-367.
[86] 瞿攀, 伏毅, 劉綿學, 等. 植物-微生物聯合修復鎳污染土壤研究進展 [J]. 土壤, 2019, 51(1): 11-18.
[87] Ju W L, Jin X L, Liu L,. Rhizobacteria inoculation benefits nutrient availability for phytostabilization in copper contaminated soil: Drivers from bacterial community structures in rhizosphere [J]., 2020, 150: 103450.
[88] Sun Z, Yang L M, Han M,. Biological control ginseng grey mold and plant colonization by antagonistic bacteria isolated from rhizospheric soil ofMeyer [J]., 2019, 138: 104048.
[89] 張鵬鵬. 幾株植物根際促生細菌釋放的揮發性物質對擬南芥及病原菌的影響 [D]. 泰安: 山東農業大學, 2013.
[90] Rajer F U, Wu H J, Xie Y L,. Volatile organic compounds produced by a soil-isolate,FA26 induce adverse ultra-structural changes to the cells ofssp., the causal agent of bacterial ring rot of potato [J]., 2017, 163(4): 523-530.
[91] Gu S H, Wei Z, Shao Z Y,. Competition for iron drives phytopathogen control by natural rhizosphere microbiomes [J]., 2020, 5(8): 1002-1010.
[92] Sheng M M, Jia H K, Zhang G Y,. Siderophore production by rhizosphere biological control bacteriaGZDF3 ofand its antifungal effects on[J]., 2020, 30(5): 689-699.
[93] 郭冰怡, 董燕紅. 細菌群體感應抑制劑研究進展 [J]. 農藥學學報, 2018, 20(4): 408-424.
[94] Jose P A, Krishnamoorthy R, Kwon S W,. Interference in quorum sensing and virulence of the phytopathogenpv.byandspecies [J]., 2019, 64(4): 423-433.
[95] Wu L M, Huang Z Y, Li X,. Stomatal closure and SA-, JA/ET-signaling pathways are essential forFZB42 to restrict leaf disease caused byin[J]., 2018, 9: 847.
Research progress on effects of rhizosphere microorganisms on quality formation of medicinal plants and their interaction mechanisms
ZHU Lei, YAN Hui, LIU Pei, ZHANG Zhen-yu, ZHANG Sen, GUO Sheng, JIANG Shu, DUAN Jin-ao
State Administration of Traditional Chinese Medicine Key Laboratory of Chinese Medicinal Resources Recycling Utilization, Jiangsu Collaborative Innovation Center of Chinese Medicinal Resources Industrialization, National and Local Collaborative Engineering Center of Chinese Medicinal Resources Industrialization and Formulae Innovative Medicine, Nanjing University of Chinese Medicine, Nanjing 210023, China
The rhizosphere plays important roles in the exchange of information and material among plant-soil-microbes, and is also the major factor effecting on plant growth and stress resistance. The interaction between plant roots and rhizosphere microorganisms is critical for plant growth and quality formation. Plant roots can specifically promote the growth of beneficial microorganisms by secreting organic acids, sugars, and secondary metabolites, etc., and inhibit the proliferation of plant pathogens through its autoimmune systems and the secretion of antibiotics. Rhizosphere microbes can participate in the growth, development, metabolic processes, and active components accumulation of medicinal plants in both direct and indirect ways, and have important contributions on the nutrient absorption, utilization, soil-borne disease control, and abiotic stress responses of medicinal plants. Research progress on effects of rhizosphere microorganisms on quality formation of medicinal plants and their interaction mechanisms are reviewed in this paper, in order to provide reference for further understanding the interaction between rhizosphere microorganisms and medicinal plants.
rhizosphere; soil; microorganisms; medicinal plants; quality formation; mechanism
R282.2
A
0253 - 2670(2021)13 - 4064 - 10
10.7501/j.issn.0253-2670.2021.13.030
2020-11-10
國家自然基金面上項目(81773848);國家中藥標準化項目(ZYBZH-C-JS-34);財政部和農業農村部: 現代農業產業技術體系建設專項(CARS-21);名貴中藥資源可持續利用能力建設項目(2060302)
祝 蕾(1997—),女,碩士研究生,研究方向為中藥資源與鑒定。Tel: 13222030367 E-mail: zhulei0402@163.com
嚴 輝(1980—),男,博士,副教授,研究方向為中藥資源與資源化學。E-mail: glory-yan@163.com
段金廒(1956—),男,教授,主要從事中藥資源化學與資源循環利用研究。Tel: (025)85811291 E-mail: dja@njucm.edu.cn
[責任編輯 崔艷麗]
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13