利用CRISPR/Cas9 系統構建低DCPC 雜質含量的CPC 工業高產菌種
2021-07-26 08:14:08徐燕,馮濤,儲炬
華東理工大學學報(自然科學版) 2021年4期
徐 燕, 馮 濤, 儲 炬
(1. 華東理工大學生物反應器工程國家重點實驗室,上海 200237;2. 國藥集團威奇達藥業有限公司,山西大同 037300)
頭孢菌素C(CPC)被Abraham和Newton 兩位學者從頂頭孢霉(Acremonium chrysogenum)培養液中分離鑒定[1-2],隨后,以CPC 為原料的頭孢菌素類抗生素逐漸成為世界抗生素市場的重要一員。該類抗生素屬于β-內酰胺類抗生素,具有廣譜抗菌性,且相較于青霉素,其療效更好,安全性更高,副作用更少。此外,由于CPC 中二氫噻嗪環與β-內酰胺環相連的特殊母核結構,使其具有更好的青霉素酶耐受性[3]。
CPC 去除支鏈后得到的7-氨基頭孢烷酸(7-ACA)是各類頭孢菌素衍生藥物的起點化合物,常通過發酵手段獲得。在發酵過程中雖常含有青霉素N(PEN)、脫乙酰氧頭孢菌素C(DAOC)等中間產物,但脫乙酰頭孢菌素C(DCPC)為主要副產物。DCPC 結構與性質和CPC極為相似,兩者均為氨基酸,它常以內銨鹽形式存在,發酵生產中DCPC 的質量分數約為CPC 質量分數的15%~20%[4]。在現有的生產工藝中,DCPC 雖能夠與主產物CPC 進行分離,但其在余液中的大量殘留會造成環境污染。抗生素的長期累積,會使周圍土壤微生物與動植物生長進化均受到影響,相關聯的上下游生物也會受到干擾,從而影響該地區的生態平衡。雖然目前有許多學者對CPC 廢液中副產物的再利用進行了大量的研究,但是由于可操作性與經濟效益等問題,真正應用于實際生產的還少之又少。
DCPC 的累積有兩方面原因:一方面,頂頭孢霉自身DCPC 乙酰轉移酶(DCPC-AT)啟動子強度不足,轉錄量低,使DCPC 無法有效轉化為CPC,成為生產過程中的限制性步驟之一[5];另一方面,頂頭孢霉Ⅷ染色體上含有CPC 乙酰水解酶基因(cahB),菌體經72 h 培養后可編碼表達一個1.4 kb 的單鏈轉錄子,翻譯出的乙酰水解酶(CPC-AH)可將已有的CPC 水解為DCPC,CPC-AH 酶活性雖然從120 h 開始下降,但144 h 仍然能觀察到活性[6]。
近年來,原為細菌和古細菌的獲得性自我免疫預防機制CRISPR/Cas9 系統[7]在基因編輯領域大放異彩,在釀酒酵母(Saccharomyces cerevisiae)中,已實現多基因[8-9]、多位點[10]、長片段(大于30 kb)[11]的成功編輯,在米曲霉(Aspergillus oryzae)[12]、構巢曲霉(Aspergillus nidulans)[13]、里 氏 木 霉(Trichoderma reesei)[14]等真菌中有多種應用,極大提高了真菌中基因編輯效率。頂頭孢霉屬于半知菌門真菌,與其他絲狀真菌相比,其生長周期長且無有性生殖階段。本實驗室首先在頂頭孢霉野生型CGMCC3.3795中構建了CRISPR/Cas9 系統,并對聚酮合酶相關基因(sorA、sorB)分別進行了敲除,編輯效率遠高于傳統方法[15]。
本研究利用CRISPR/Cas9 系統,對頂頭孢霉發酵過程中雜質DCPC 累積問題進行了定向改造,構建基因cahB缺陷菌株與基因cefG過表達菌株,以期降低DPCP 雜質含量,提高CPC 產量與質量。
1 材料與方法
1.1 材料
1.1.1 菌株與質粒 頂頭孢霉(Acremonium chrysogenum)1-D1 工業菌株、大腸桿菌(Escherichia coli)DH5α均由本實驗室保藏。真菌表達質粒pAN7-1 由本實驗室保存,含RGR(Ribozyme-gRNA-Ribozyme)結構質粒pUC57 由華大基因合成,含CRISPRCas9 基因編輯系統質粒pAN7-sorA 由本實驗室構建與保存。
1.1.2 培養基與培養方法 麥芽汁培養基(用于工業頂頭孢霉菌株斜面培養)、種子培養基(用于頂頭孢霉菌株種子培養)、搖瓶發酵培養基(用于頂頭孢霉菌株發酵),配方與培養方法參見文獻[16],并略有調整(種子培養基pH 為7.2)。原生質體培養基與轉化方法參見文獻[14]。
1.1.3 儀器與檢測方法 利用高效液相色譜儀(日本島津有限公司LC-20T)對CPC 和DCPC 進行檢測。CPC 檢測方法參考文獻[16]。DCPC 檢測:流動相為體積分數50%的甲醇與磷酸鹽的混合溶液(體積比為3∶97),流速為1 mL/min,等度洗脫,檢測波長為254 nm,柱溫為30 ℃,進樣量為20 μL,檢測時間35 min。
1.1.4 酶與試劑 質粒提取試劑盒、膠回收試劑盒:Axygen 公司;真菌基因組抽提試劑盒、真菌RNA 提取試劑盒:生工生物工程(上海)股份有限公司;Evo M-MLV Reverse Transcriptase 反轉錄試劑盒與SYBR? Green Premix qPCR 試劑盒:湖南艾科瑞生物工程有限公司;限制性內切酶:NEB 公司;一步克隆試劑盒、Phanta酶等PCR 酶:Vazyme 公司;CPC 標準品與DCPC 標準品(含量未知)由國藥集團山西威奇達藥業有限公司饋贈。四丁基氫氧化銨(Tetrabutylammonium Hydroxide):色譜級,上海阿拉丁生化科技股份有限公司。流動相磷酸鹽(用于CPC 與DCPC 檢測):850 mL超純水中加入0.8 g NaH2PO4,攪拌混勻后,用四丁基氫氧化銨調節pH 至7.3,0.45 μm 濾膜抽濾除雜,超聲30 min 去除氣泡。
1.1.5 引物合成與測序 測序、引物合成均由生工生物工程(上海)股份有限公司完成。表1 為引物列表,其中大寫字母為模板質粒pUC57 中RGR 特異性結構的骨架。

表1 引物列表Table 1 List of primers

續表1
1.2 sgRNA 設計與載體構建
經NCBI 數據庫查詢,得到頂頭孢霉野生型( ATCC11550) CPC 乙 酰 水 解 酶( CPC-AH) 基 因cahB(ACRE_083500),通過與工業菌1-D1 全基因測序結果進行比對,找到工業菌中相對應的基因序列(GME1727_g)。通過sgRNA 設計網站(https://gt-scan.csiro.au/submit/)設計靶目標序列5’-gcggcaagaccggccacatgAGG-3 ’ 。 設 計 引 物 sgcahB-1、 sgcahB-2、sgcahB-3、sgcahB-4(見表1)。以pUC57 為模板,進行PCR 擴增,回收得到長度分別為84 bp 與204 bp的兩片段,將片段融合PCR 后獲得可識別cahB基因的特異性sgRNA 片段cahB-sgRNA。
將質粒pAN7-sorA 雙酶切(HindIII、SpeI)膠回收含Cas9 蛋白表達盒的大片段質粒骨架,與cahBsgRNA 片段一步克隆連接,構建pAN7-cahB質粒。將質粒轉大腸DH5α感受態,挑取單菌落,液體培養后提質粒送測序。載體pAN7-cahB 構建電泳圖見圖1 所示。
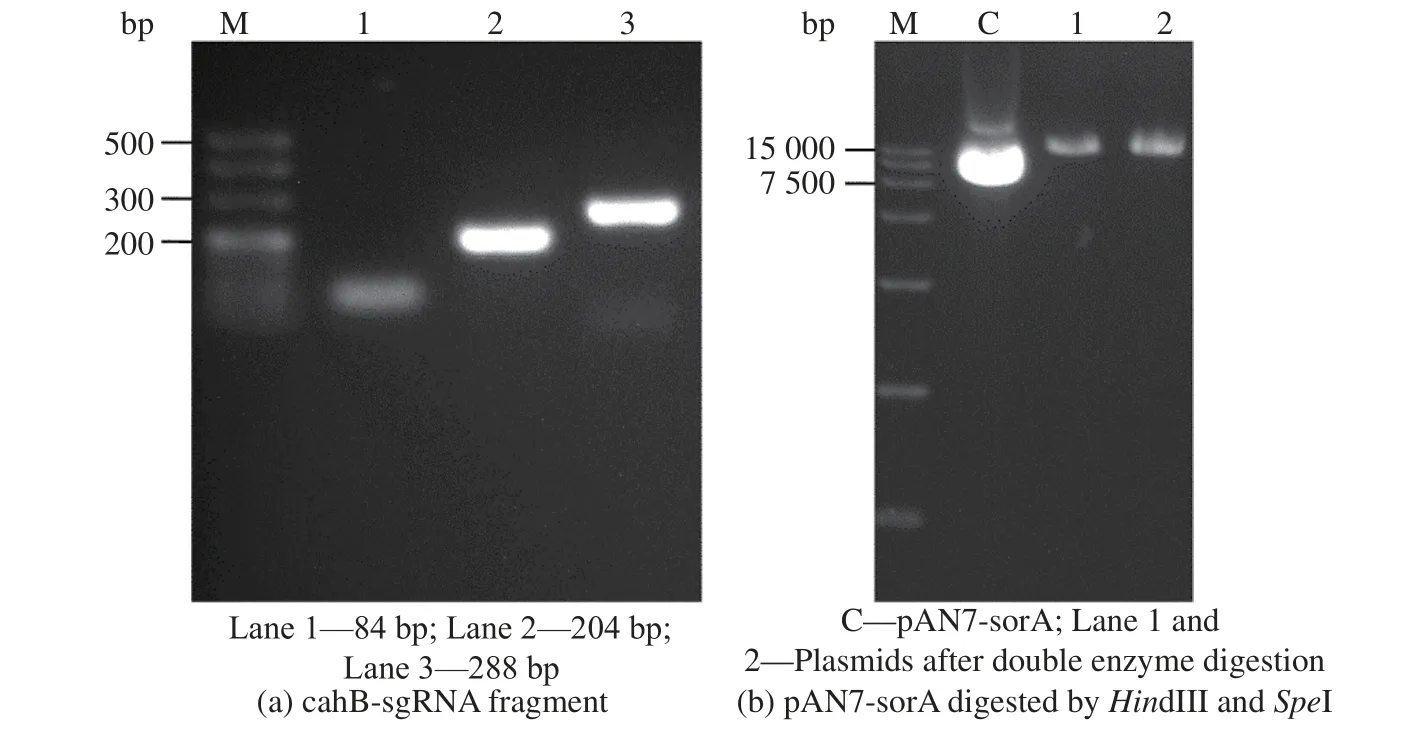
圖1 pAN7-cahB 構建電泳圖Fig. 1 Electrophoretogram of pAN7-cahB construction
1.3 cefG 基因表達盒與Donor DNA 構建
依據NCBI 數據庫中cefG序列(GI: 672 794 352),比對本實驗室已有測序結果,獲得工業菌株1-D1中cefG序列,設計引物,以1-D1 基因組為模板擴增,獲得cefG基因片段,見圖2。由圖可見,以pAN7-1為模板,設計反向引物(R-pAN7-1-F,R-pAN7-1-R),PCR 得到含啟動子與終止子的反向骨架PgpdApAN7-Ttrpc,采用一步克隆的方法將骨架與cefG基因片段連接,構建cefG表達質粒pAN7-1-cefG,進行大腸桿菌感受態轉化,挑單菌落PCR 初步驗證后,提質粒測序。

圖2 pAN7-1-cefG 構建電泳圖Fig. 2 Electrophoretogram of pAN7-1-cefG construction
Donor DNA 構建流程與原理見圖3。首先,依據1-D1 測序結果,選取cahB基因上下游長度為1 000 bp左右片段作為同源臂,PCR 擴增獲得同源臂B1、B2。以Split-Marker 重組技術為原理,設計引物(cahB-PT-F1,cahB-PT-R1)、(cahB-PT-F2,cahB-PTR2),將同源修復片段分上下游片段進行導入。將質粒上PgpdA-cefG-Ttrpc 表達盒分兩部分進行擴增得到PgpdA-cefG(2 286 bp)、cefG-TtrpC(2 370 bp),中間含200 bp重疊區。電泳條帶見圖4(a)。采取融合PCR 方式與上下游同源臂分別連接,獲得Donor DNA(B1-PgpdA-cefG、cefG-TtrpC-B2),見圖4(b)。
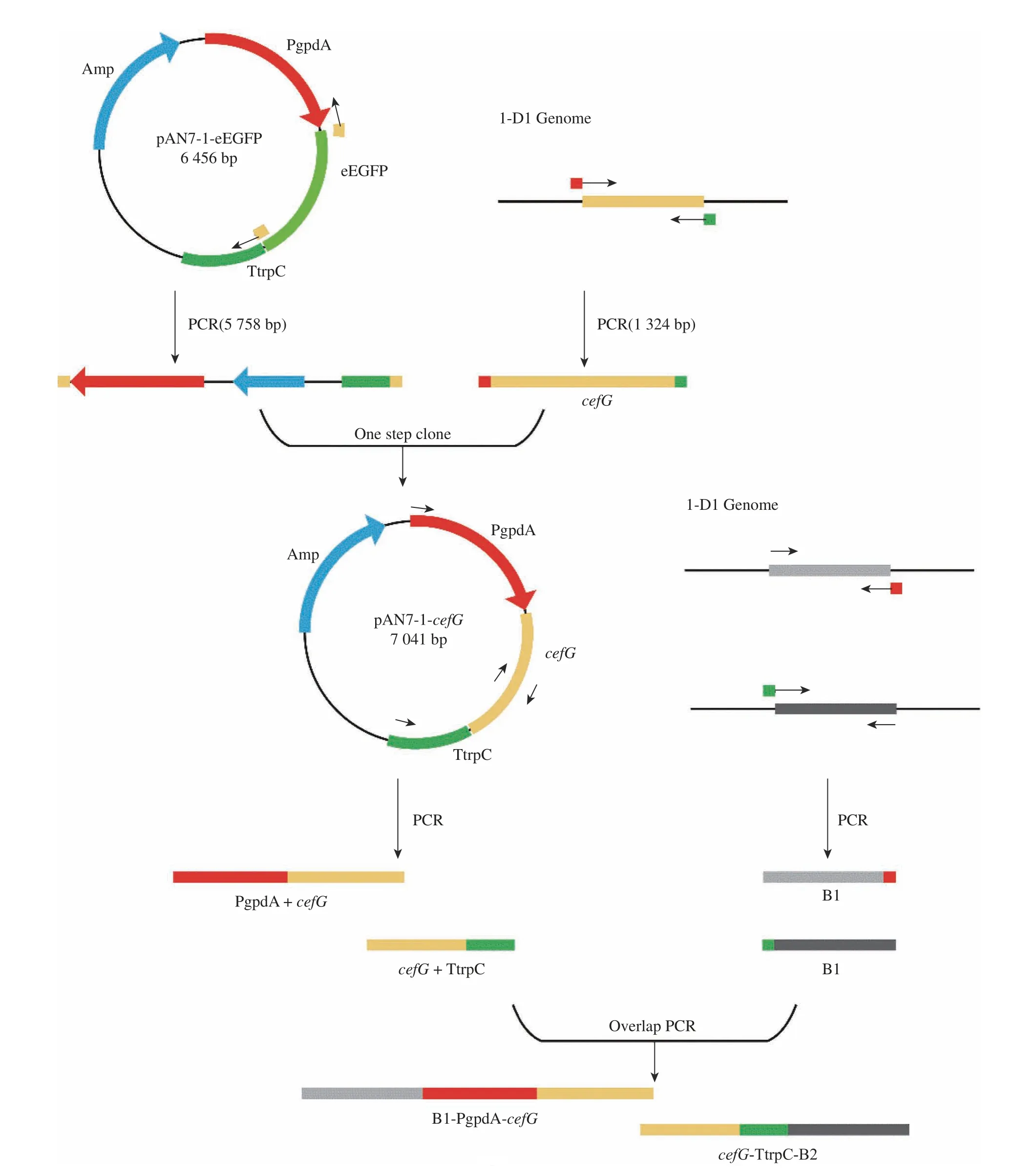
圖3 Donor DNA 構建流程圖Fig. 3 Donor DNA generation flow chart
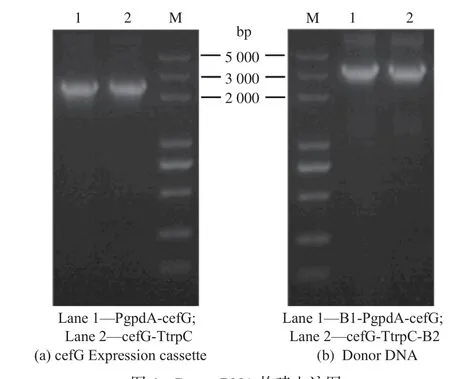
圖4 Donor DNA 構建電泳圖Fig. 4 Electrophoretogram of donor DNA construction
1.4 原生質體轉化
將構建完成的pAN7-cahB 質粒與同源修復片段(B1-PgpdA-cefG、cefG-TtrpC-B2)進行頂頭孢霉原生質體共同轉化,在PEG 介導下,將外源質粒與同源修復表達盒導入菌體內,進行特異性打靶與同源修復。轉化時,外源質粒的質量為8~10 μg,pAN7-cahB 質粒、同源修復片段(B1-PgpdA-cefG)和同源修復片段(cefGTtrpC-B2)質量比為4∶1∶1,總體積不超過20 μL(當無需同源修復片段協同作用時,外源質粒所需量不變但總體積不超過10 μL)。當外源核酸物質與原生質體輕柔混勻后,將該體系與上層培養基混合后倒至含有下層轉化培養基的平板中,28 ℃培養10~12 d,待轉化板長出半透明的白色菌落,使用已滅菌牙簽轉移至麥芽汁平板(潮霉素抗性:150 μg/mL),28 ℃培養7 d。轉化步驟見圖5。
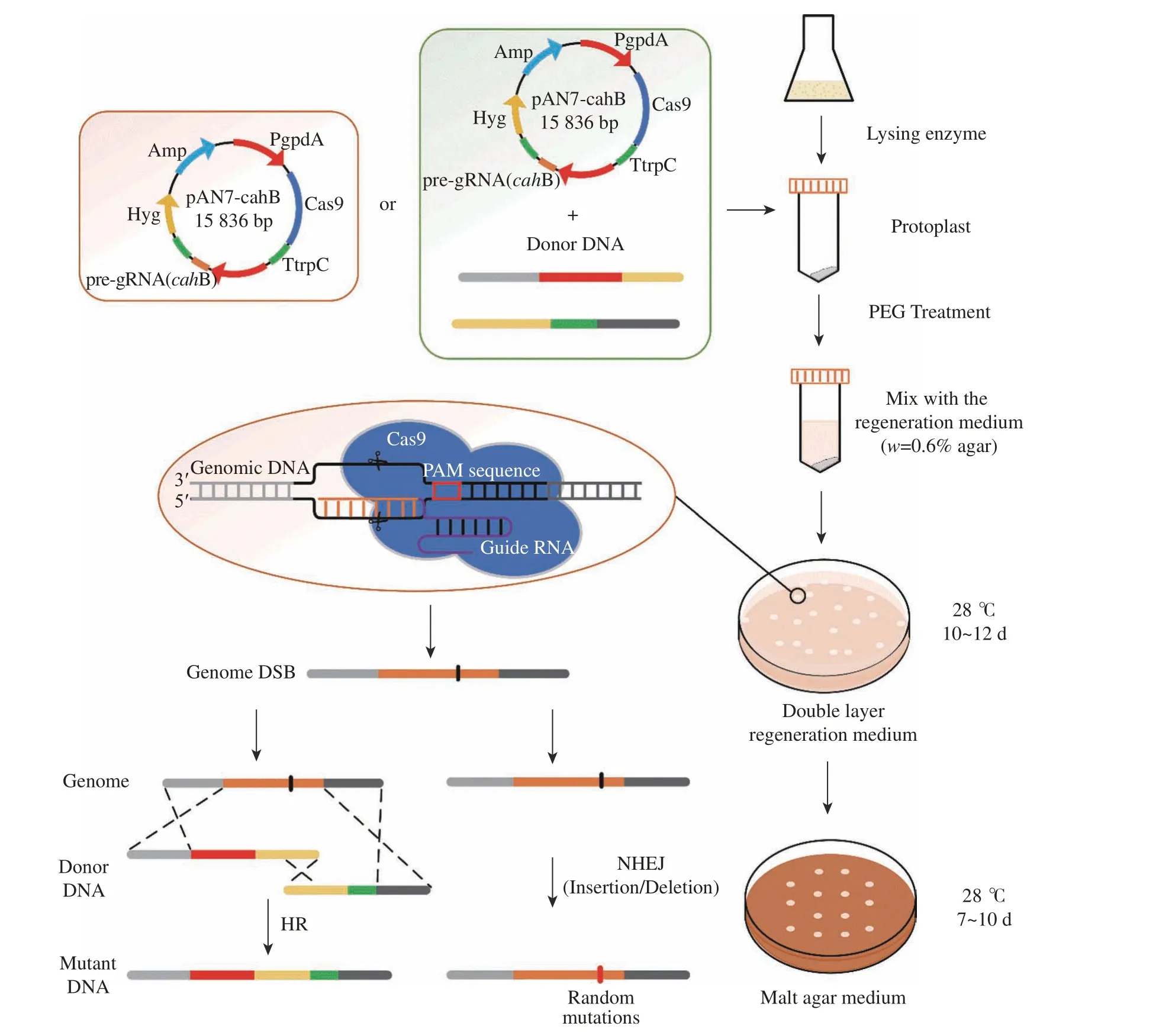
圖5 原生質體轉化實驗流程圖Fig. 5 Flow chart of protoplast transformation experiment
1.5 轉化子篩選驗證
1.5.1 Ac-ΔcahB 驗證方法 在PAM 序列上下游500~1 000 bp 處設計引物(C-veri-F,C-veri-R),以基因組為模板進行PCR,獲得的片段(1 500 bp)通過T7EI 酶切來初步驗證是否與基因組有錯配堿基,再進行測序驗證,結果見圖6。轉化子測序結果為5’-gcggcaagaccggccac--gAGG-3’,將其與原基因組進行比對,發現PAM 序列(AGG)的上游有2 個堿基缺失。將轉化子進行遺傳穩定性驗證,發現傳代至第5 代基因組時堿基缺失仍然存在,說明采用單質粒轉化方法進行敲除的菌株,其遺傳穩定性良好,沒有產生回復突變。
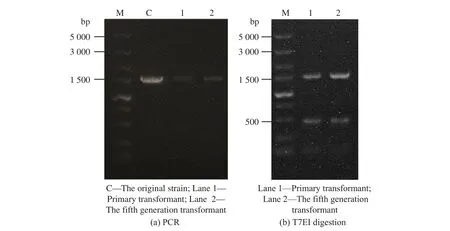
圖6 Ac-ΔcahB 轉化子驗證結果Fig. 6 Ac-ΔcahB transformant verification results
單質粒轉化在驗證時發現,部分轉化子無法進行PCR 獲得驗證片段,這可能是由于Cas9 蛋白切割造成的雙鏈斷裂所引發的非同源性末端接合(NHEJ),導致基因組其他部分的大片段插入與缺失,使驗證引物無法與基因組匹配。但是由于其插入與缺失長度的不確定性,無法很好通過調整引物與片段長度解決,故采取單質粒與Donor DNA 共轉方式以較好解決突變株驗證問題。
1.5.2 Ac-ΔcahB:cefG 驗證方法 轉化子驗證原理見圖7。首先,以C1-1、C1-2 為引物擴增靶基因序列,對照組1-D1 有擴增片段,實驗組應無擴增片段;而以C2-1、C2-2 為引物擴增同源修復片段中cefG基因表達盒的部分序列,對照組1-D1 應無擴增片段,實驗組應有擴增片段;再以C3-1、C3-2、C4-1、C4-2 為引物分別擴增上游同源臂B1 與表達盒啟動子部分、表達盒終止子與下游同源臂B2 部分,對照組1-D1應無擴增片段,而實驗組應有擴增片段。最后用C3-1、C4-2為引物擴增時,對照組與實驗組條帶長度有明顯差異,電泳結果見圖8。
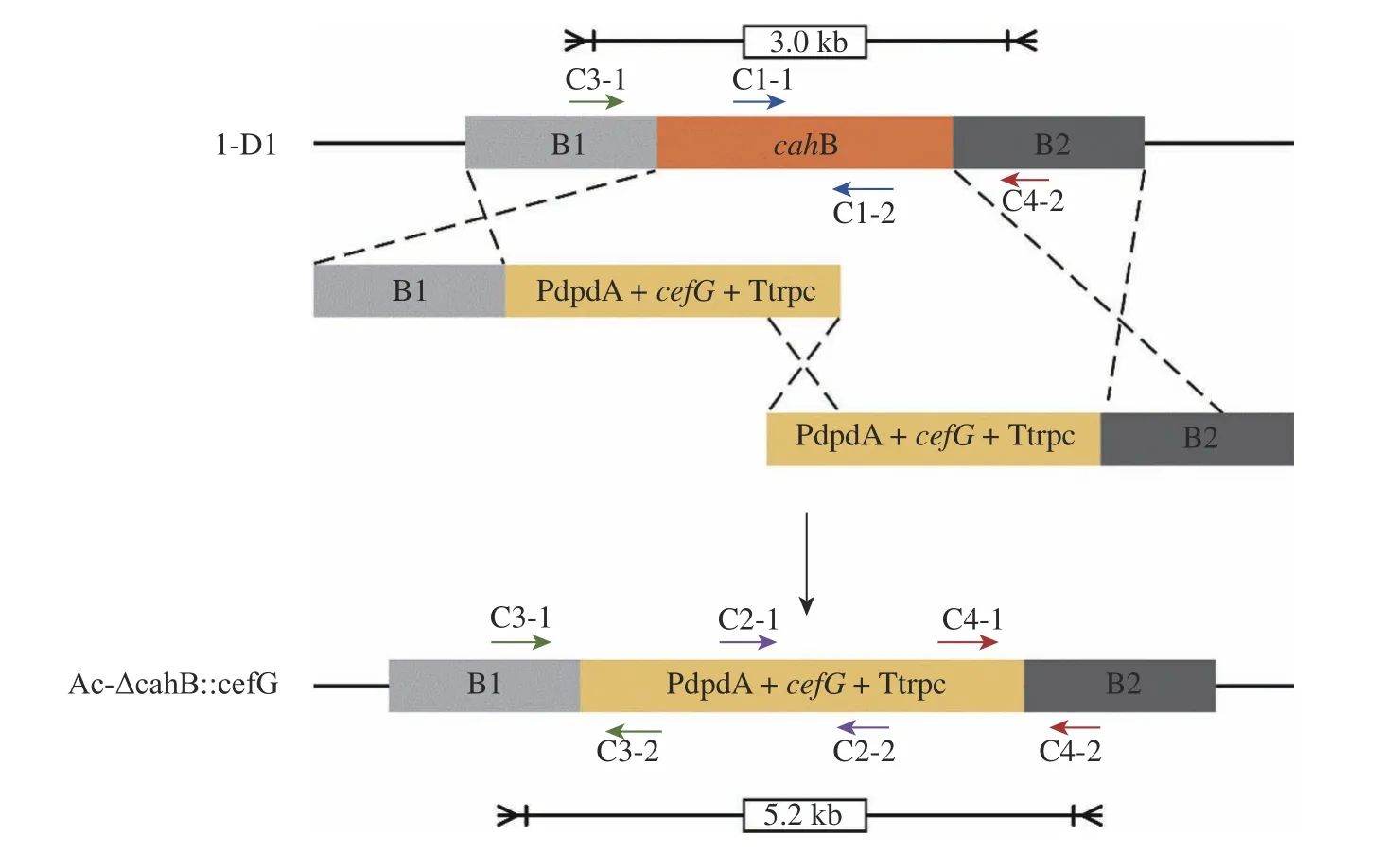
圖7 Ac-ΔcahB::cefG 驗證原理圖Fig. 7 Ac-ΔcahB::cefG verification principle diagram
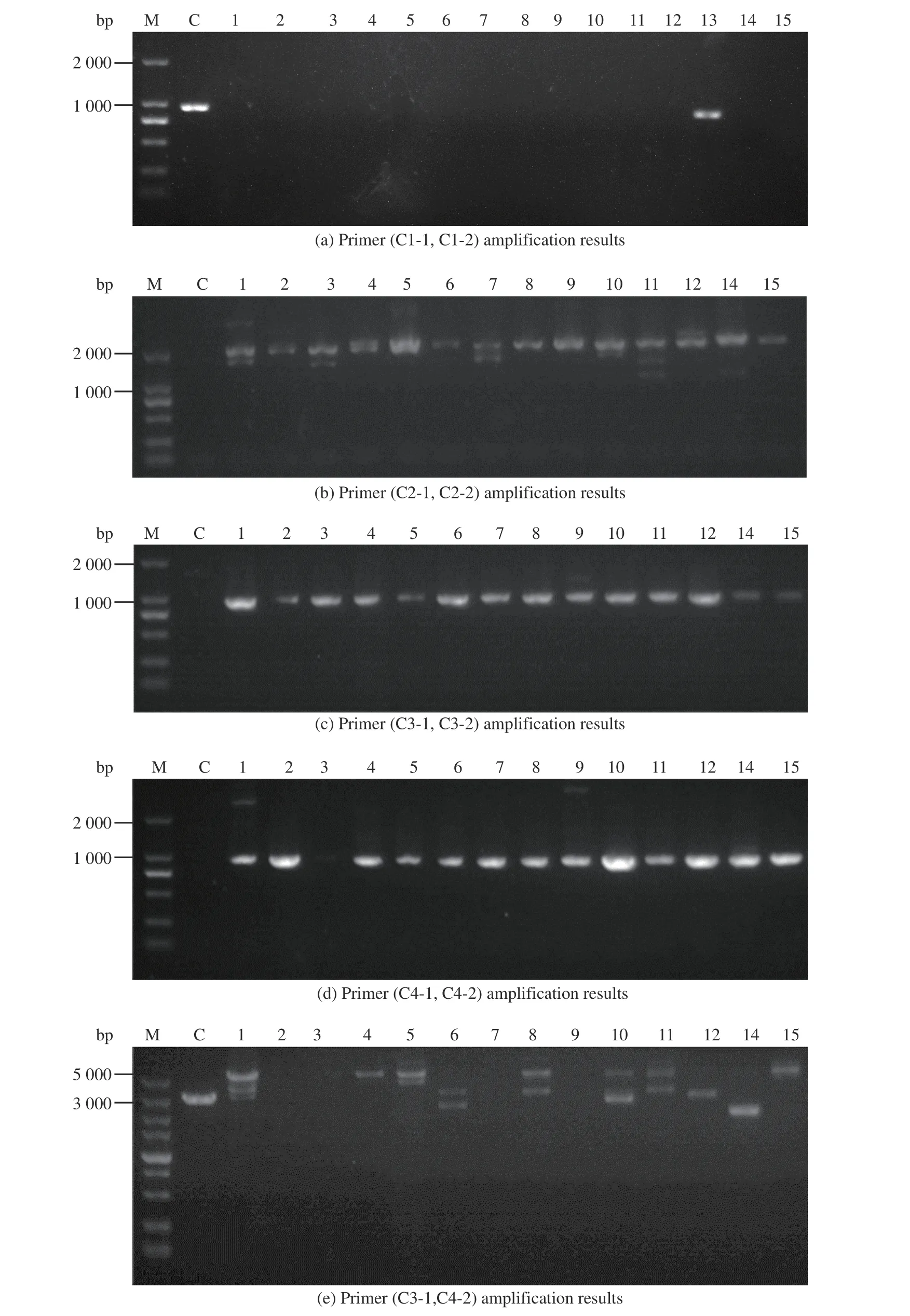
圖8 Ac-ΔcahB::cefG PCR 驗證圖Fig. 8 Ac-ΔcahB::cefG PCR verification chart
1.6 RNA 提取以及RT-PCR(Red Time Quantitative PCR)分析相關基因轉錄水平
取培養72、96 h 后的頂頭孢霉改造菌株Ac-ΔcahB、Ac-ΔcahB::cefG 與出發菌株1-D1 的發酵液,去盡上清并用DEPC(焦碳酸二乙酯)水洗滌3 次后,進行液氮研磨,按照真菌RNA 提取試劑盒說明書要求進行RNA 提取。確認RNA 質量與濃度后,使用Evo M-MLV Reverse Transcriptase 反轉錄試劑盒說明書進行反轉錄制備cDNA。以此為模版,以cefG-F1、cefG-R1 為引物,使用SYBR? Green Premix qPCR 試劑盒進行RT-PCR 擴增。
2 結果與討論
2.1 出發菌株與突變菌株的CPC 產量比較
將突變株Ac-ΔcahB、Ac-ΔcahB::cefG 與對照工業菌株1-D1 進行168 h 的液體搖瓶發酵,對CPC產量與雜質DCPC 產量進行比較研究。CPC 產量如圖9所示,突變株Ac-ΔcahB的CPC產量為4 665 μg/mL,出發菌株的CPC 產量為4 578 μg/mL,兩者發酵水平相接近。而突變株Ac-ΔcahB::cefG 的CPC 產量為6 072 μg/mL,相較于出發菌株,總體提高了32.6%,差異顯著(p=0.025 9)。以上結果可推測,插入強化表達的cefG基因可使DCPC 得到有效轉化,減少了菌體中的積累,在降低雜質的同時進一步提高了CPC 的產量。
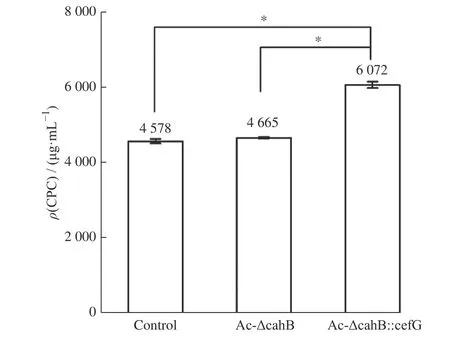
圖9 出發菌株與突變株CPC 產量比較Fig. 9 CPC production comparison of starting strain and mutant strain
cahB基因是CPC 乙酰水解酶基因,可將CPC 水解成為無經濟價值的DCPC。但是在此結果中發現,敲除該基因后CPC 并未達到預期中明顯提高的效果,這可能由于水解的CPC 在總產量中所占比重比較小。有學者對CPC 乙酰水解酶進行了酶動力學反應研究,結果表明,純化至均質的酶蛋白與CPC 的親和力不高(Km=33.7 mmol/L),同時其酶活性在發酵至120 h 時開始逐漸下降[6]。底物CPC 濃度較低時,與乙酰水解酶結合轉化的效率較低,水解能力較弱,因乙酰水解酶減少的CPC 在總產量中所占比重比較小,故敲除后CPC 產量沒有明顯提升。此外,在對基因組進行分析時發現,菌體中有可能存在其他CPC 水解酶,如GME4736_g 與GME1825_g(圖10),其分別注釋為堿性蛋白酶(Alkaline proteinase)與角質層降解蛋白酶(Cuticle-degrading protease),均具有絲氨酸型肽鏈內切酶活性,而cahB與枯草芽孢桿菌的絲氨酸蛋白質家族基因具有高同源性。相較于cahB,這些注釋為絲氨酸家族的蛋白質基因對CPC 的水解可能起主導作用。
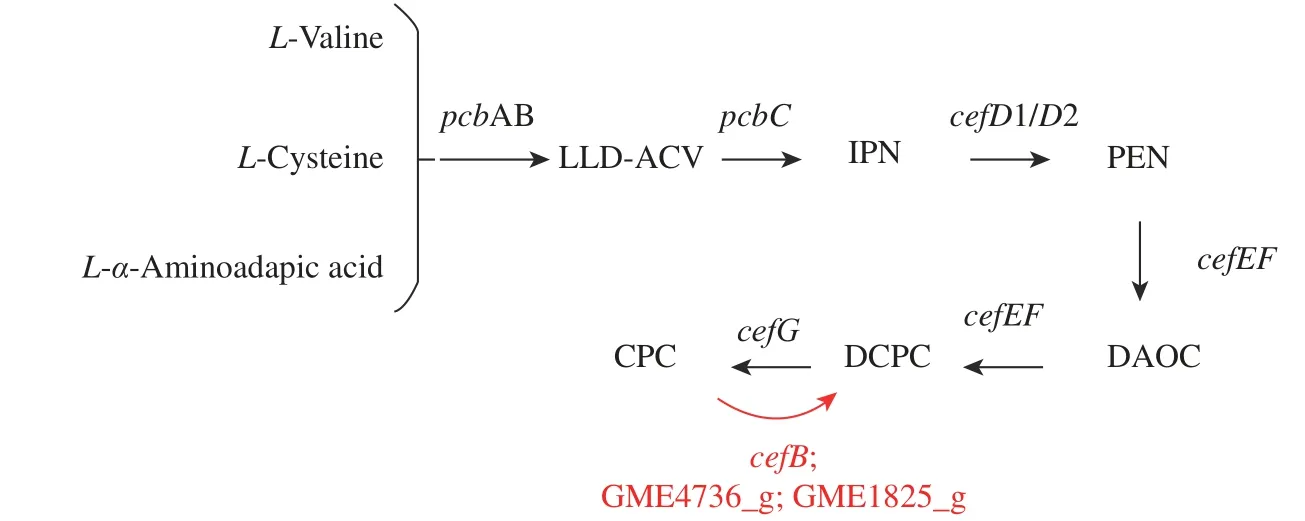
圖10 CPC 合成與分解過程的相關基因Fig. 10 Genes involved in the process of CPC synthesis and decomposition
2.2 出發菌株和突變菌株的DCPC 產量比較

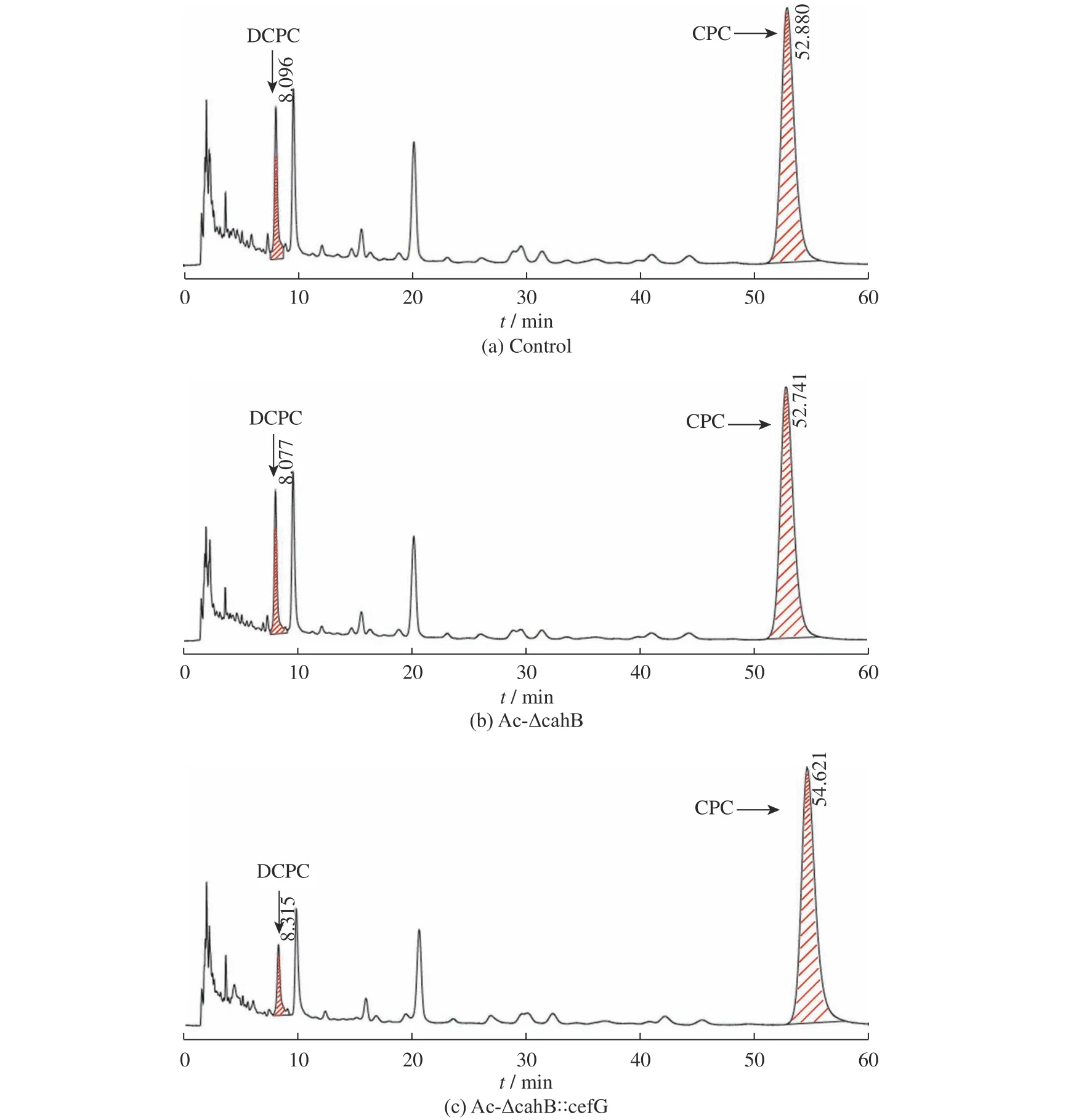
圖11 出發菌株與突變株發酵液高效液相色譜圖Fig. 11 HPLC of the fermentation broth of starting strain and mutant strain
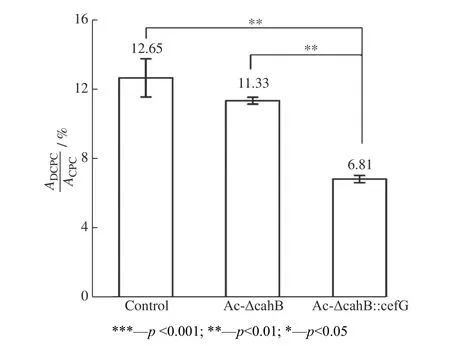
圖12 出發菌株與突變株DCPC 的含量比較Fig. 12 DCPC content comparison of starting strain and mutant strain

2.3 基因轉錄水平測定
以出發菌株1-D1 為對照,提取發酵72 h 與96 h的菌體RNA 進行熒光定量PCR(RT-qPCR)分析。分別以菌株中γ-actin 為內參基因進行矯正,對DCPC乙酰轉移酶基因cefG進行表達量測定,結果見圖13所示。在頂頭孢霉中,DCPC 乙酰轉移酶基因cefG內源性啟動子較弱,基因表達強度較低一直是CPC 生產過程中的限制性步驟。在Ac-ΔcahB::cefG菌株中,強啟動子PgpdA提高了cefG表達量,且上調倍數高達5 倍。DCPC 乙酰轉移酶基因轉錄量的提升可能增加了菌體內乙酰轉移酶的含量,從而提高了菌體對中間產物DCPC 的轉化效率,促進了CPC 合成。
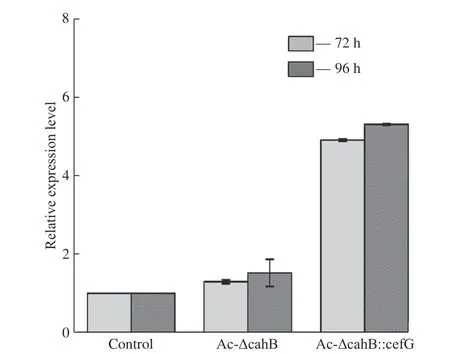
圖13 DCPC 乙酰轉移酶基因cefG 相對表達量測定Fig. 13 Determination of cefG expression level of DCPC acetyltransferase gene
3 討 論
頭孢菌素類抗生素因其毒性低、抗菌譜廣、作用效果好等特點被廣泛應用于臨床治療中,而CPC 作為頭孢菌素類抗生素的重要原料,其生產菌株頂頭孢霉的發酵卻常有中間產物與類似物作為雜質殘留,對CPC 產量以及分離純化造成影響。
本文運用CRISPR/Cas9 基因編輯系統構建了Ac-ΔcahB 菌株,并通過結合Donor DNA,優化了敲除效率,構建了Ac-ΔcahB::cefG 菌株。cahB基因的敲除雖不能有效提高CPC 產量,但是從適宜種齡以及平板培養等其他生長狀況而言,該基因的敲除對菌體生長并沒有過多影響。或許,相較于以往隨機插入的過表達方法,將cahB基因作為定點整合位點,是一個便于驗證的不錯選擇。此外,在對基因組進行分析時發現,菌體中有可能存在其他CPC 水解酶,如堿性蛋白酶(Alkaline proteinase)與角質層降解蛋白酶(Cuticle-degrading protease),均具有絲氨酸型肽鏈內切酶活性,并且cahB與枯草芽孢桿菌的絲氨酸蛋白質家族基因具有高同源性。相較于cahB,可能這些注釋為絲氨酸家族的蛋白質基因對CPC 的水解起著主導作用。插入由強啟動子PgpdA過表達的cefG基因,使突變菌株Ac-ΔcahB::cefG 在轉錄水平表達量提高了5 倍,并且在CPC 生產與DCPC 雜質方面較出發菌株均差異顯著,既使CPC 產量提高了32.6%,又使DCPC含量控制在6.81%。此外,發酵培養基的有效利用,提高了經濟效益,同時由于DCPC 含量的降低減少了廢液中抗生素的殘留,從而減小了廢液排放對環境的影響。
