海島棉GbGSTU7的克隆及其對枯萎病菌侵染的應答
2021-07-28 02:23:38陸小雙韓萬里曲延英陳全家鄧曉娟
西北植物學報 2021年6期
關鍵詞:植物
陸小雙,陳 琴,韓萬里,曲延英,陳全家,鄧曉娟
(新疆農業大學 農學院/棉花教育部工程研究中心,烏魯木齊 830052)
棉花是全球范圍內最重要的經濟作物之一。中國是目前世界上最大的棉花生產國,由于海島棉纖維比陸地棉的纖維更長、更強、更細,使其成為栽培棉中纖維品質最優的類型,并且是紡高支精梳紗和特種紡織物的關鍵原料,在紡織工業中占有重要地位[1]。新疆棉區作為中國唯一的海島棉種植及生產基地,在國民經濟中占有很重要的地位,棉花生產的興衰對中國乃至世界都會產生十分重要的影響。新疆棉區雖然具有獨特的自然條件,適合棉花的種植,但也存在許多問題。特別是棉花枯萎病 (Fusariumoxysporumf.sp.vasinfectum.Fov),近年來由于連年種植,枯萎病已嚴重影響海島棉生產,給棉花生產帶來了巨大的經濟損失[2],使新疆海島棉的種植面積和產量都大量減少。因此,因海島棉枯萎病導致減質減產的問題急需被解決,而解決這一問題的有效辦法就是棉花抗病新品種的選擇和培育。海島棉自身枯萎病抗性基因的挖掘是關鍵點,所以利用生物信息學、分子遺傳學和基因工程的方法為海島棉抗枯萎病品種的選育提供候選基因顯得尤為重要。
在長期的進化過程中,植物逐漸形成了一系列的抗逆機制,其中,谷胱甘肽途徑在此過程中發揮著重要作用[3]。谷胱甘肽轉移酶(glutathione S-transferases, GSTs)是一種多功能酶[4],是廣泛存在于自然界中的小分子多功能型水溶性蛋白[5]。GSTs一般通過促進還原型谷胱甘肽與各種含親電基團底物的相互作用,形成復合物,將有害物質運輸至體外,從而起到解毒消毒作用[6]。GST被認為是在微生物、動植物酶解毒、細胞信號轉導和其他細胞過程中起著必不可少的作用[7]。其主要功能之一是能夠提高植物細胞膜的修復能力,在基因工程中可用于提高植物的抗逆能力[8]。GST包含一個N末端域和一個C末端域。N末端結構域包括谷胱甘肽(GSH)結合和催化的催化殘基,而保守性較低的C末端結構域(包含5或6個主要螺旋)結合疏水性底物并決定GST的特異性和活性[9]。在非生物脅迫反應中,GST催化GSH的巰基與外源親電子類物質結合來降低生物細胞內ROS水平,從而保護植物免受損傷[10]。在病原菌等生物脅迫反應中,植物通過增強GST同源基因的表達,將有毒物質運送至液泡加工解毒以減輕對植物的損傷[11-12]。因此,GST基因在植物抵御各種生物和非生物脅迫的進程中均起著非常重要的作用。
課題組前期對具有高抗枯萎病的海島棉品系06-146與高感枯萎病品種‘新海14’為親本雜交構建的F7重組自交系群體中的抗感病品種進行轉錄組測序[13],測序結果提示GSTU7基因可能參與抗枯萎病過程。本研究以海島棉抗枯萎病材料06-146為試驗材料,獲得GbGSTU7基因(CottonFGD登錄號:GB_A03G0575),對其核苷酸和氨基酸序列進行分析。用抗枯萎病材料06-146與感枯萎病材料‘新海14’為試驗材料,利用實時熒光定量PCR技術對該基因在枯萎病菌侵染下的表達進行研究,旨在探討基因與棉花抗枯萎病之間的關系,為進一步研究海島棉GSTU7基因的生物學功能奠定理論基礎。
1 材料和方法
1.1 材 料
海島棉抗枯萎病材料06-146,感枯萎病材料‘新海14’,枯萎病菌(生理小種7號),均由新疆農業大學作物遺傳育種實驗室提供和保存。
1.2 方 法
1.2.1 棉花種植與培養挑選飽滿脫絨種子,1%(體積分數)升汞消毒后,無菌水沖洗多次后將種子置于鋪有濕潤濾紙的發芽盒中,置于人工氣候室培養(25 ℃恒溫,16 h光照/8 h黑暗)培養至芽長3 cm左右,移至Hoagland’s營養液中培養。待第一片真葉完全展開時,接種稀釋至孢子數為1×107個·mL-1的枯萎病菌(生理小種7號)。選取0 h及接菌后4、8、12、24和48 h 的棉株下胚軸、莖和葉片,經液氮速凍處理,-80 ℃超低溫冰箱保存備用。
1.2.2 接種鑒定培養基制備。制備馬鈴薯固體培養基和液體培養基,用于棉花枯萎病菌的培養。
病菌培養。挑取枯萎病菌單菌落接種于馬鈴薯固體培養基上進行擴大培養,25 ℃恒溫培養5~7 d,再將培養后的枯萎病菌接種于馬鈴薯液體培養基中,25 ℃、 r·min-1震蕩培養1周以上,用細胞計數器計數菌液濃度達到1×107個·cm-2備用。
棉花枯萎病菌侵染棉花。室內種植棉花幼苗待2片子葉展平時,用傷根法完成枯萎病菌對棉花的侵染。
接種鑒定。在接種3周后開始進行室內病情的調查和統計,待病情穩定后調查發病率。
分級標準采用國家0~5級的判斷標準[14]。
1.2.3 RNA提取與cDNA合成RNA提取采用天根(TIANGEN)生化科技有限公司的RNA提取試劑盒(RNA plant Plus Reagent),提取后的RNA采用愛必夢生物科技有限公司的反轉錄試劑盒(First-Strand cDNA Synthesis Kit) 進行反轉錄,cDNA樣品用ddH2O稀釋后,置于-20 ℃備用。
1.2.4GbGSTU7基因的克隆和測序通過CottonFGD數據庫得到GSTU7基因的序列,并設計用于克隆該基因編碼區全長的引物,引物交由上海華大基因科技有限公司合成。以提取的cDNA為模板進行PCR擴增,PCR反應經94 ℃預變性5 min后,以94 ℃變性30 s,51.6 ℃退火40 s,72 ℃延伸45s為循環,循環數為35個,最后經72 ℃延伸5 min 后終止反應,4 ℃保存。采用1.5%瓊脂糖凝膠電泳對PCR產物進行分離檢測,用Tian Gen公司DNA產物純化試劑盒對含有目的DNA 片段的PCR反應液進行純化。將純化產物與pEASY-T1 (全式金)克隆載體連接,轉入Trans-T1 (全式金)感受態,均勻涂布在含有50 ng·μL-1卡那霉素的LB 固體培養基上(酵母提取物5 g,胰蛋白胨10 g,氯化鈉10 g,瓊脂粉15 g,定容至1 L),正置培養1 h后,37 ℃倒置過夜培養,挑取陽性單克隆經菌液PCR驗證正確后送華大公司測序。
1.2.5 序列生物信息學分析用NCBI-ORF FINDER找到測序序列的開放閱讀框,利用ExPAXSyProtParam (http://web.expasy.org/protparam/)分析該蛋白的物理化學特性,利用TMHMM-2.0(http://www.cbs.dtu.dk/services/ TMHMM-2.0/)和SignalP (http://www.cbs.dtu.dk/services/SignalP/)分析目的蛋白是否存在跨膜區和信號肽,使用EMBL的PFAM(https://pfam.xfam.org/search/sequence)及ExPASy的PROSITE(http://prosite.expasy.org/)對推導出的GSTU7蛋白的功能結構域進行預測。借助NCBI下載相似性高的基因序列,在Clust Omega(https://www. ebi.ac.uk/Tools/msa/clustalo/)中進行多序列比較。利用MEGA 10構建系統發育進化樹。運用PSIPRED(http://bioinf.cs.ucl.ac.uk/psipred/)和SWISS MODEL (https://www.swissmodel.expasy.org/)對蛋白質的二級、三級結構進行分析。通過在線預測軟件SoftBerryProtComp9.0(http://www.SoftBerry.com)對蛋白進行亞細胞定位。
1.2.6 實時熒光定量PCR利用美國ABI公司7500 Fast System熒光定量PCR儀進行實時熒光定量PCR。以棉花GhUBQ7為內參基因,與目標基因一起擴增,每個處理設3次生物學重復,按照abm SYBR qPCR Master Mix試劑盒的操作說明進行實時熒光定量PCR,檢測目的基因的表達,所用引物見表1。以棉花UBQ7基因的轉錄表達水平作為內參,用2-ΔΔCT法計算目的基因的相對表達水平。
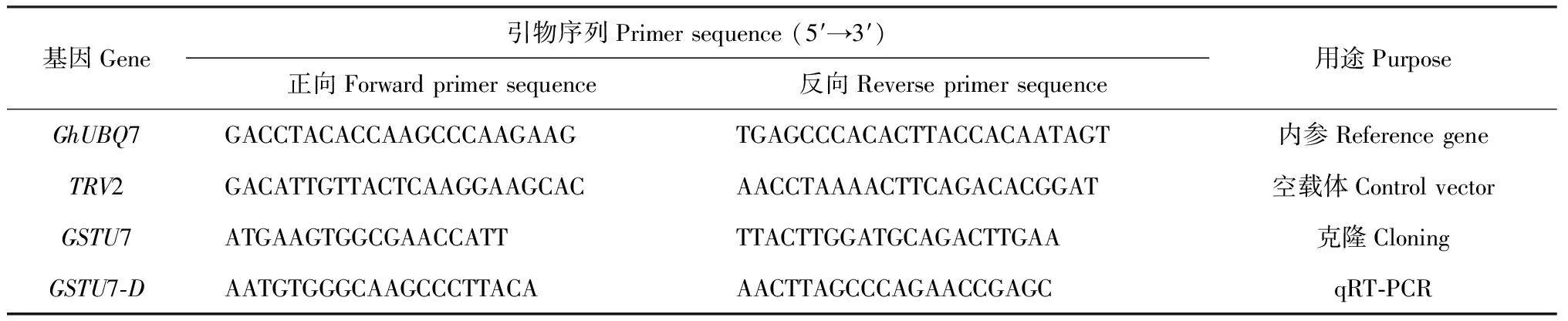
表1 本研究所用引物
2 結果與分析
2.1 室內抗枯萎病鑒定結果
對本研究所涉及的‘新海14’及06-146進行室內枯萎病接種鑒定,結果如表2所示,‘新海14’的枯萎病發病率顯著高于06-146,平均高70%。根據國家標準[14],‘新海14’屬于感病材料,06-146屬于抗病材料。

表2 室內抗枯萎病鑒定結果
2.2 目的基因片段的獲得
以06-146的cDNA為模板,利用表1中GSTU7的上下游引物進行PCR擴增,得到711 bp的目的片段(圖2)。PCR產物純化后連接到Trans-T1載體上(圖2,A),通過含有50 ng·μL-1卡那霉素的LB 固體培養基篩選獲得陽性克隆菌落(圖2,B),通過菌液PCR可以看到含有711 bp的目的條帶(圖2,C),送往上海華大基因科技有限公司測序。序列測定與分析表明,海島棉GSTU7基因含有1個711 bp的開放閱讀框,編碼236個氨基酸(圖1)。

圖1 海島棉GSTU7基因的核苷酸序列及其編碼的氨基酸序列Fig.1 The nucleotide sequence of GSTU7 gene of Island cotton and its encoded amino acid sequence
2.3 GSTU7的氨基酸序列和理化性質分析
GbGSTU7蛋白的預測分子式是 C1263H1960N324O351S5,相對分子量約為27.5 ku,等電點7.84。該蛋白中含量最多的氨基酸是賴氨酸,占氨基酸總數的9.7%,帶正電荷總體氨基酸殘基(Arg+Lys)數和帶負電荷總體氨基酸殘基(Asp+ Glu)數分別為33個和32個。親水性平均數-0.314,推測其為親水蛋白。不穩定系數33.48,是不穩定蛋白。對該蛋白進行跨膜區和信號肽分析發現 GSTU7蛋白不含信號肽,不存在跨膜區。將GbGSTU7基因的氨基酸序列進行 BLAST比對,結果顯示,該蛋白屬于tau類 GST 蛋白。將家族成員基因的蛋白序列載入MAGA-X,使用classX方式完成多序列比對,采用臨近法完成系統發生樹的構建(圖3)。由進化樹可見:該基因家族共鑒定到32個成員共分為3類。第一類共有4個基因成員,第二類共有4個基因成員,第三類共有24個基因成員,GbGSTU7(GB_A03G0575)屬于該家族第三類成員。將該基因序列進行功能預測,預測該基因可能被水楊酸誘導,與還原型谷胱甘肽與大量外源和內源性疏水親電試劑的結合有關,并且對某些除草劑具有解毒作用,與植物生長發育相關,可能參與的KEGG途徑有谷胱甘肽代謝和毒素分解代謝過程。
依據氨基酸序列多重比對結果將該蛋白分別與古巴棉(Gossypiumbarbadense) KAB2038401.1、澳洲棉(Gossypiumaustrale)KAA3456760.1、橡膠(Heveabrasiliensis)XP_021652560.1、等植物中的GSTU7基因編碼蛋白做多序列比對并進行同源性分析,發現海島棉GSTU7基因編碼的蛋白序列與亞洲棉、陸地棉、雷氏棉等GSTU7氨基酸序列相似性較高,分別是100%、98.23%、97.88%,與黃連、荔枝、橡膠等物種的GSTU7氨基酸序列相似性較低,均在65%以下(圖4)。
2.4 海島棉GSTU7的生物信息學分析
對海島棉GSTU7基因推導的氨基酸序列進行疏水性/親水性分析,結果表明該蛋白為親水蛋白。該蛋白第174位的纈氨酸(Val) 親水性最強,其次為第170和第171位的丙氨酸(Ala) 和異亮氨酸(Ile);疏水性最強的位點為第16位的谷氨酸(Glu),其次為第14的天冬氨酸(Asp) (圖5)。該蛋白二級結構包含5個β-折疊和9個α-螺旋,三級結構空間結構像呈球狀。海島棉GSTU7基因與毛棉、陸地棉,亞洲棉,雷蒙德氏棉親緣關系較近,與荔枝、柑橘等親緣關系較遠。海島棉、毛棉的GST屬于同一分支,古巴棉、陸地棉、雷氏棉、海島棉、毛棉、亞洲棉、澳洲棉均屬于同一分支,荔枝、柑橘的GST屬于同一分支,橡膠、黃連的GST屬于同一分支(圖6)。亞細胞定位預測結果顯示,發現該蛋白既可在細胞質中表達,也可在細胞核中表達。

M.DL2000;A.1.PCR純化產物;2.目的基因連接Trans-T1載體之后的質粒;B.1-3.陽性克隆菌落;C.1-8.菌液PCR結果;D.1.目的片段 圖2 海島棉GSTU7 基因 PCR 產物電泳分析M. DL2000; A. 1. PCR purified products; 2. The plasmid after the target gene was attached to the trans-T1 vector; B.1-3. Positive colonies; C.1-8. PCR results of bacterial fluid; D.1. Target fragmentFig.2 Electrophoresis analysis of PCR products of Island cotton GSTU7 gene

海島棉H7124數據庫中GST_N_3基因家族成員圖3 家族系統發生樹Registration numbers are GST_N_3 gene family members in H7124 database of Island cottonFig.3 Family phylogenetic tree
2.5 枯萎病菌侵染后海島棉GSTU7基因的表達分析
由圖7可知,正常條件下,海島棉GSTU7基因在棉花的根、莖和葉中均有表達。
圖7,A、B顯示,在根中,‘新海14’(感病材料)該基因響應緩慢,06-146(抗病材料)接菌后立即上調響應,而‘新海14’(感病材料)到24 h才上調表達對抗。接菌后,該基因的表達量在06-146(抗病材料)的各組織中表現出有規律的先上升后下降,根、莖、葉中依次表達,表現出與病菌傳播途徑一致的方向,表明該基因起到了正面抵抗作用。‘新海14’(感病材料)則不同,在根中響應緩慢,在莖和葉中呈上升-下降-上升-下降的變化,表現出盲抗反應,對病菌擴散未起到持久抑制作用。

GB_A03G0575.1. 海島棉;KAB2038401.1. 古巴棉;KAA3456760.1. 澳洲棉;XP_021652560.1. 橡膠;KDO66687.1. 柑橘;XP_031286334.1. 黃連;EOY33900.1. 可可;XP_012471261.1. 雷氏棉;ALY05896.1. 荔枝;XP_016740535.1. 陸地棉;TYI35251.1. 毛棉;XP_017641138.1. 亞洲棉;KAE8705650.1. 木槿;AT2G29420.1. 擬南芥; 圖6同圖4 海島棉GSTU7與其他植物 GSTU7氨基酸序列的多重比對GB_A03G0575.1. Gossypium barbadense L; KAB2038401.1. Gossypium barbadense; KAA3456760.1. Gossypium australe;XP_021652560.1. Hevea brasiliensis; KDO66687.1. Citrus sinensis; XP_031286334.1. Pistacia vera; EOY33900.1. Theobroma cacao; XP_012471261.1. Gossypium raimondii; ALY05896.1. Litchi chinensis; XP_016740535.1. Gossypium hirsutum;TYI35251.1. Gossypium tomentosum; XP_017641138.1. Gossypium arboreum; KAE8705650.1. Hibiscus syriacus; AT2G29420.1. Arabidopsis thaliana; The same as Fig.6Fig.4 Multiple alignment of the amino acid sequences of Island cotton GSTU7 and other plant GSTU7
圖7,C-E顯示,在06-146(抗病材料)中,隨著接菌時間的延長,GbGSTU7基因在根中的表達量呈下降趨勢,在莖和葉中的表達量均呈先上升后下降的趨勢,且在處理24 h時莖的表達量達到最高。接菌24 h時,海島棉GSTU7基因在莖和葉中的表達量較接菌4 h有明顯上升,但根中的表達量明顯下降。在‘新海14’(感病材料)中,枯萎病菌侵染后4 h時,海島棉GSTU7基因在根和莖中的表達量較0 h無明顯變化,但在葉片中的表達量達到最高,隨著接菌時間的延長,海島棉GSTU7基因在‘新海14’(感病材料)根中的表達量呈下降-上升的趨勢,葉中的表達量呈上升-下降-上升的趨勢,且在處理8 h時莖的表達量達到最高。
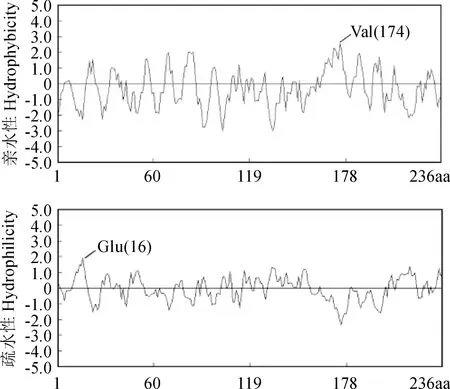
Val. 纈氨酸;Glu. 谷氨酸圖5 海島棉GSTU7 氨基酸序列的親水性和疏水性分析Val. Valine acid;Glu. Glutamic acidFig.5 Hydrophilicity and hydrophobicity analysis of GSTU7 amino acid sequence of Island cotton
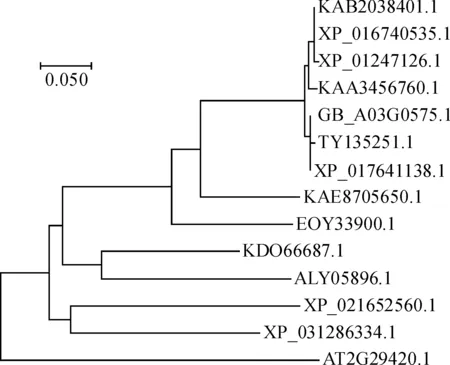
圖6 海島棉與其他植物 GSTU7 氨基酸序列的系統進化樹Fig.6 Phylogenetic tree of GSTU7 amino acid sequences of Island cotton and other plants
3 討 論
前人研究表明,植物在遭遇逆境脅迫時,其體內的活性氧含量會大量升高,會對細胞結構和代謝物質造成氧化傷害,嚴重影響甚至擾亂植物的正常生長、發育和代謝[15-16]。清除活性氧的過氧化物酶系在生物體內普遍存在,包括谷胱甘肽過氧化物酶(glu-tathione peroxidase,GPX)、超氧化物歧化酶(superoxide dismutase,SOD) 、過氧化氫酶(catalase, CAT)和抗壞血酸過氧化物酶(ascorbate peroxidase,APX),能夠有效地清除植物體內的活性氧及其他自由基,消除細胞內的有害代謝物質,從而保護植物免受或減緩傷害[17]。其中,GST作為一個大的基因家族,在植物生長發育、響應環境變化的過程中發揮重要的調節作用[18]。GSTs對生長素和細胞分裂素有很高的親和力,在茄屬植物中,受病原真菌侵染的植物表現GST含量隨著細胞內生長素水平的增加而快速增加[19],這表明GSTs在激素穩態和植物抵御病原菌中起著重要作用[20]。抗黃萎病棉花品種‘農大601’,在病毒誘導基因沉默抑制GST基因簇后極易感病,這表明GST簇在黃萎病抗性中起重要作用[21]。因此,研究GST有助于闡明植物生長和抵御脅迫的分子機制,對改善脅迫下植物的生存能力具有實際意義。

A.06-146;B. 新海14;C.根;D.莖;E.葉; *表示P<0.05,**表示P<0.01圖7 海島棉GSTU7基因在枯萎病菌侵染后不同材料及組織的表達A. 06-146; B. Xinhai 14; C.Root;D.Stem; E. Leaves. * Indicates P<0.05, ** Indicates P<0.01Fig.7 Expression of GSTU7 gene of Island cotton in different materials and tissues after infection by Fusarium wilt
GSTs一般以25~27 kD的2條亞基的同源或異源方式聚合而成,每個亞基都含有2個空間結構不同的結構域:N端由β折疊和α螺旋構成;C端主要由α螺旋構成[18]。本研究克隆獲得的海島棉GSTU7基因屬于tau類GST家族成員,其編碼的蛋白質與其他物種中GST序列相似性較低,但在結構上相對保守,N端具有保守的結合谷胱甘肽的G位點,C端的H位點可結合疏水底物[5],在空間結構上與前人研究結果類似,N端主要由β折疊和α螺旋構成(βαβαββα),C端由6個α螺旋構成,通過1個約11個氨基酸的短序列與N端結構域連接。
研究發現,玉米中大多數GST基因在根中表達水平較高,而在其他組織中表達相對較低[22]。本研究中,海島棉GSTU7基因在棉花各組織中表達差異明顯,莖中的表達量最高,其次是葉,在根中的表達量較低,呈現明顯的組織特異性。前人研究表明,干旱、真菌攻擊、鹽脅迫等均能誘導GSTs的表達[21, 23-25]。過表達大豆GmGSTL基因提高了煙草和擬南芥在鹽脅迫下的存活率[7]。擬南芥中過表達番茄GST基因增強了其在鹽脅迫和干旱脅迫條件下的生長勢[26]。煙草過表達GhGST結果表明,轉基因煙草植株對黃萎病的抗性明顯增強,棉花抗病品種在病毒誘導基因沉默抑制GST基因簇后極易感病[21]。這表明,GSTs 在植物抵御不良環境和損傷等方面也起著重要作用。
本研究結果表明,克隆的海島棉GSTU7基因屬于GST家族tau類成員,枯萎病菌侵染后,該基因在抗病材料06-146和感病材料‘新海14’的各組織均可響應誘導,并且在根、葉中均存在顯著差異,莖中存在極顯著差異。在抗病材料06-146莖中的表達量最高,其次是葉,在根中的表達量較低,具有明顯的組織特異性。說明該基因可在植物體遭受枯萎病菌侵染時,各組織隨著病原菌的擴散做出應答,推測其可能與棉花抗枯萎病有著密切的關系,可能在棉花抵御病菌侵染的過程中發揮重要的調控作用,但其具體的作用機制還有待進一步研究。本研究結果為鑒定海島棉GSTU7基因功能奠定了一定的理論基礎。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
