纖枝短月蘚LEA5基因表達及耐鹽性分析
2021-07-28 02:13:28李雪寶
西北植物學報 2021年6期
李雪寶,周 璇,鄢 波
(西南林業大學 園林園藝學院, 昆明 650224)
纖枝短月蘚(Brachymeniumexile)屬于真蘚科短月蘚屬[1],能在裸露的巖壁和極端干旱的環境中生存,具有較強的耐旱能力。研究植物抗旱基因,培育耐旱植物,對改善生態環境具有重要意義。
由于非生物脅迫嚴重限制了植物的生長發育,一系列防御機制已經進化,以提高植物的抗逆性,包括組織結構、化學物質和表達蛋白的反應[2-3]。其中,晚期胚胎發生豐富蛋白(LEA)是一種高度親水性的蛋白質,最早是在棉花胚胎發育后期的子葉中發現的[4],隨后在小麥、甘藍型油菜、番茄、印度芥菜等植物中也發現LEA蛋白的存在[5-8],LEA蛋白在植物界具有廣泛的分布。但它并不是植物界所特有的,在細菌、真菌和一些無脊椎動物中也存在LEA蛋白[9-11]。
根據蛋白質氨基酸基序和保守結構特點,LEA蛋白家族的分類存在差異,按照Pfam domain分類法[12],本研究所用的LEA5蛋白屬于LEA5族(對應以往分類法中的D19[13]和group 1[14]),該LEA蛋白一般由80~153個富含甘氨酸的序列組成,其最顯著的特征是含有高度保守的20個氨基酸基元序列[15]。
此外,研究表明大部分的LEA蛋白對干旱、高滲性等各種非生物脅迫具有響應機制。例如,小麥TaEm蛋白(LEA5族)可以提高釀酒酵母對高鹽的耐受能力[16];大麥HVA1蛋白(LEA3族)的過度表達顯著提高了轉基因小麥和水稻對干旱脅迫的耐受性[17];AtLEA4-1d蛋白在芥菜中的過度表達可以顯著增強轉基因植物對干旱和鹽脅迫的適應性[8];OsLEA3-2蛋白的過表達可以改善轉基因擬南芥和水稻在鹽脅迫和干旱條件下的生長[18]。因此,LEA蛋白在植物非生物脅迫反應中的生物學功能越來越受到重視。
目前,有關苔蘚植物LEA5基因的研究較少。本研究對纖枝短月蘚LEA5基因序列及其啟動子元件進行分析,并對LEA5基因的表達及耐鹽性進行了探究,為進一步探討LEA5基因結構及其耐鹽性之間的關系提供依據,也為進一步研究LEA5基因的耐鹽保護機制奠定基礎,為提高植物耐鹽性的分子育種提供了潛在的應用前景。
1 材料和方法
1.1 植物材料
纖枝短月蘚采于西南林業大學,選取該苔蘚植株的不同發育時期及不同組織(孢子體部分及配子體部分),立即用液氮速凍后保存于-80 ℃。
1.2 方 法
1.2.1 纖枝短月蘚LEA5基因ORF的克隆采用OMEGA公司的總RNA提取試劑盒提取纖枝短月蘚的總RNA,利用逆轉錄試劑盒(全式金),將提取的RNA逆轉錄為cDNA。
根據本課題組已有的轉錄組測序結果,結合載體序列設計同源臂引物LEA5-PF(GATGACGATGACAAGATGGCTTCCCGGGAGGAG)和LEA5-PR(AATTCGAAGATCTCGTCACTCGTTTGTAA-ACTTAGAC)。以纖枝短月蘚cDNA第一鏈為模板,進行PCR擴增。PCR擴增程序:95 ℃預變性3 min;95 ℃變性15 s,56 ℃退火 30 s,72 ℃延伸 90 s,35個循環;72 ℃延伸10 min;4 ℃保存。
1.2.2 纖枝短月蘚LEA5基因啟動子的擴增取新鮮的纖枝短月蘚置于預冷的研缽中,加入適量的液氮充分研磨,具體操作參照北京天根公司的小量植物(葉)DNA抽提試劑盒說明書。
以纖枝短月蘚DNA為模板,結合LEA5基因序列設計啟動子引物LEA5-1R(GTACCTTCTGCGAGGTGCTCTTGGGC)、LEA5-2R(ACGATGGACTCCAGTCCGGCCGAAGCCATTCTCGTT-GTGCTC)和LEA5-3R(GATGATCAGAAGATA-AGCAAGTGTAGG),分別作為第一輪、第二輪、第三輪特異引物, LAD1-1、LAD1-2、LAD1-3、LAD1-4、AC1引物序列及HiTail-PCR擴增程序參照Liu等的研究[19],第三輪PCR產物經1%瓊脂糖凝膠電泳檢測,選擇約1 000 bp條帶進行PCR產物回收,PCR產物回收后連接到克隆載體pMD18-T中,送生工生物工程股份有限公司進行測序。
1.2.3LEA5基因的熒光定量分析選擇LEA5基因進行熒光定量PCR分析,根據獲得的LEA5基因序列,設計熒光定量引物LEA5-QF(CTCGCAGAAGGACGTAGCAAG)和LEA5-QR(CTCATCAATGTCGATGTTCTGC)。以Actin基因為內參基因,設計引物A-F(CTGTACGGCAACATCGTGCTG)和A-R(CCAGACACTGTACTTCCTCTC),以cDNA為模板,按照TB Green?Premix Ex TaqTM Ⅱ說明書,采用2-ΔΔCt法。
1.2.4 纖枝短月蘚LEA5基因原核表達分析用KpnⅠ和SacⅠ(TaKaRa公司)對pThioHis A載體進行雙酶切。將純化的酶切產物與目的基因進行重組,具體操作參照Vazyme公司的Clon ExpressⅡOne step Cloning Kit的操作說明書,獲得重組質粒,將重組質粒送生工生物工程股份有限公司進行測序。
將構建成功的重組表達質粒以及空載體分別轉化至表達菌株E.coliBL 21 (DE 3)中。誘導表達后進行SDS-PAGE電泳分析。
1.2.5 重組菌耐鹽性分析將重組菌株與對照菌株接種到LB(100 μg/mL Amp)液體培養基中,于37 ℃,220 r/min 過夜培養。次日按照1∶100比例擴大培養至OD600為0.5時,加入終濃度為1 mmol/L IPTG進行誘導表達至OD600為0.8,做10倍梯度稀釋(1∶10;1∶100;1∶1000)。從各稀釋度的菌液取10 μL,點到添加300、500、700 mmol/L NaCl和500、700、900 mmol/L KCl脅迫培養基上,37 ℃倒置培養2 d,每個實驗重復3次,觀察菌斑生長情況,并拍照保存。
1.2.6 生物信息學分析利用ExPASy-ProtParam分析氨基酸的理化性質;利用NetPhos 3.1 Server軟件預測磷酸化位點[20];利用NetOGlyc 1.1 Server軟件預測O-糖基化位點;利用NetNGlyc 1.0 Server 預測N-糖基化位點;利用SOPMA軟件進行二級結構預測及分析;利用Swiss-Model通過同源建模建立三級模型。
2 結果與分析
2.1 纖枝短月蘚LEA5基因序列及蛋白的結構分析
轉錄組序列分析表明,該基因包含189 bp 5′-UTR和285 bp 3′-UTR,其開放閱讀框為267 bp,編碼88個氨基酸(圖1)。Pfam分析得出,LEA5蛋白在2~61氨基酸位點含有LEA5族結構域。分析表明該蛋白屬于LEA5族。

圖1 纖枝短月蘚LEA5基因的核苷酸序列及其編碼的氨基酸序列Fig.1 Nucleotide acids and deduced amino acids sequences of LEA5 gene from B. exile
利用protparam預測LEA5基因所編碼的蛋白質相對分子質量為9 157.81 Da,理論等電點4.72,分子式C377H608N120O142S2,LEA5基因所編碼蛋白的氨基酸組成中,Gly含量最高,占總氨基酸的15.9%;其次為Ala和Glu,均占總氨基酸的13.6%。脂肪系數為51.25,平均親水性為-1.011,為親水蛋白;不穩定指數為35.77,屬穩定性蛋白。
預測發現LEA5蛋白發生磷酸化修飾的位點共有6個。其中,絲氨酸的磷酸化位點最多,為4個;而蘇氨酸的磷酸化位點和酪氨酸的磷酸化位點各1個。LEA5蛋白沒有發生N-糖基化的位點,可能發生O-糖基化的修飾位點有5個。
使用SOPMA軟件預測LEA5蛋白的二級結構。結果表明,該蛋白主要由α螺旋(alpha helix)、β轉角(beta turn)、無規則卷曲(random coil)和延伸鏈(extended strand)構成,其中α螺旋和無規則卷曲最多,分別占氨基酸序列的44.32%和38.64%,其次為β轉角(12.50%),延伸鏈最少(4.55%)。通過Swiss-Model對LEA5蛋白的三級結構進行同源建模,預測的LEA5蛋白三級結構中有2個α螺旋和3個β折疊。
2.2 纖枝短月蘚LEA5基因啟動子的擴增與分析
以纖枝短月蘚DNA為模板,通過HiTail-PCR擴增LEA5基因啟動子序列,選擇第4泳道約為1 000 bp條帶進行回收(圖2),最終獲得了LEA5基因啟動子序列1 053 bp。
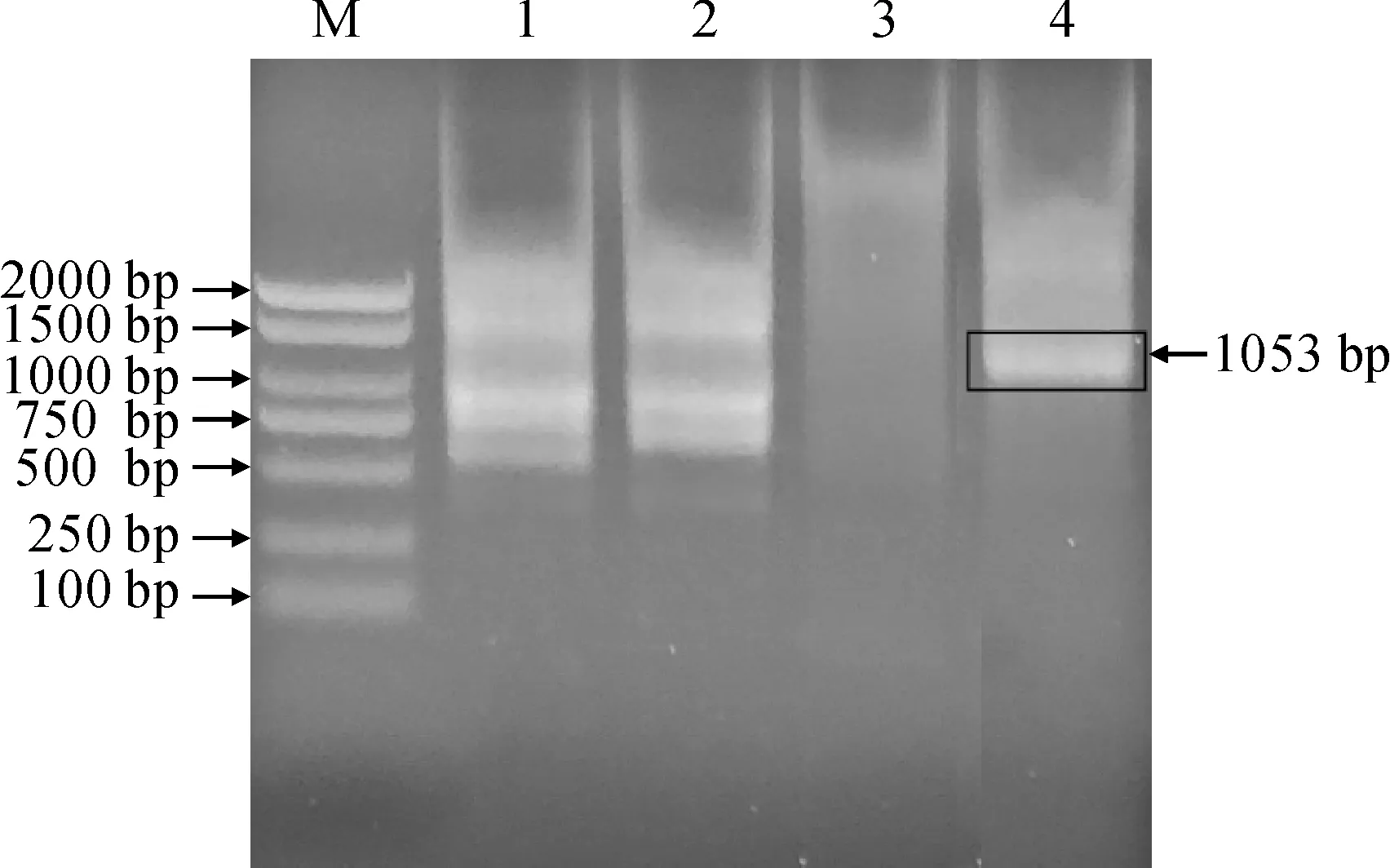
M. Marker;1-4. 分別與引物LAD1-1、LAD1-2、LAD1-3、LAD1-4結合的HiTail-PCR擴增產物圖2 纖枝短月蘚LEA5基因啟動子HiTail-PCR擴增M. Marker; 1-4. HiTail-PCR production combined with LAD1-1, LAD1-2, LAD1-3 and LAD1-4, respectivelyFig.2 HiTail-PCR production of LEA5 gene promoter from B. exile
啟動子元件分析結果顯示,LEA5基因啟動子具有典型的CAAT box元件。在LEA5基因啟動子區含大量非生物脅迫響應順式作用元件,主要包括脫落酸響應元件(abscisic acid responsive element,ABRE)、茉莉酸甲酯響應元件(methyl jasmonate response element,CGTCA-motif)、赤霉素響應元件(gibberellin re-sponse element,GARE-motif)以及MYB結合位點(MYB binding site,MBS),且含有多個MYB和MYC元件,這2類元件都屬于轉錄因子作用元件,與干旱或ABA誘導的表達調控有關,此外,還含有G-box、Wbox、TCCC-motif、TCT-motif、TGACG-motif 等元件。
2.3 纖枝短月蘚LEA5基因熒光定量分析
熒光定量結果顯示,在正常情況下,LEA5基因在纖枝短月蘚的不同時期及不同部位中皆有表達,在纖枝短月蘚無性世代的孢子體中表達量最高,有性世代的配子體次之,無性世代的配子體最低,不同組織的F檢驗非常顯著(P<0.05),說明正常情況下不同組織的表達量不同。Tukey檢驗可以得出,無性世代的孢子體和無性世代的配子體間表達量差異顯著,有性世代的配子體與其他2個組織間差異不顯著(圖3)。
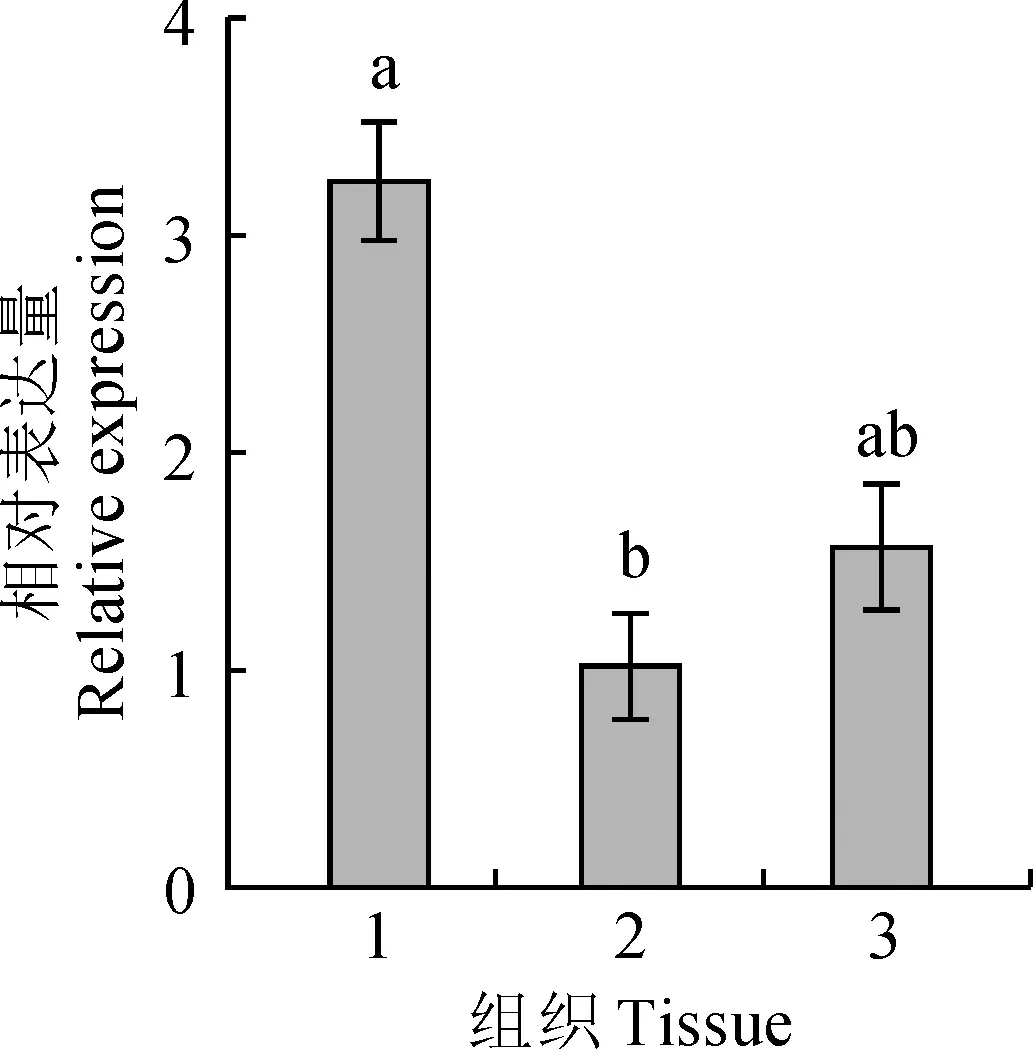
1.無性世代的孢子體;2.無性世代的配子體;3.有性世代的配子體。不同字母表示組織間差異顯著圖3 纖枝短月蘚不同組織中LEA5基因的表達分析1. Sporophyte of the asexual generation; 2. Gametophyte of the asexual generation; 3. Gametophyte of the sexual generation. Different letters indicate significant difference in different tissues Fig.3 Expression analysis of LEA5 gene in different tissues of B. exile
2.4 纖枝短月蘚LEA5基因原核表達分析
將對照菌(BL21/pThiohis A)和重組菌(BL21/LEA5)在37 ℃培養至OD600為0.5,加入終濃度為1 mmol/L的IPTG進行誘導,其中對照菌誘導6 h,重組菌分別誘導1、2、4和6 h。SDS-PAGE結果顯示:與對照菌相比,經IPTG誘導的重組菌在約22 kD處有一條目標蛋白表達條帶,并且隨著誘導時間的延長表達量增加,而僅含有空載體的對照菌則在約16 kD處有一條較小的硫氧還蛋白(為pThiohis A載體中的蛋白)表達條帶 (圖4)。
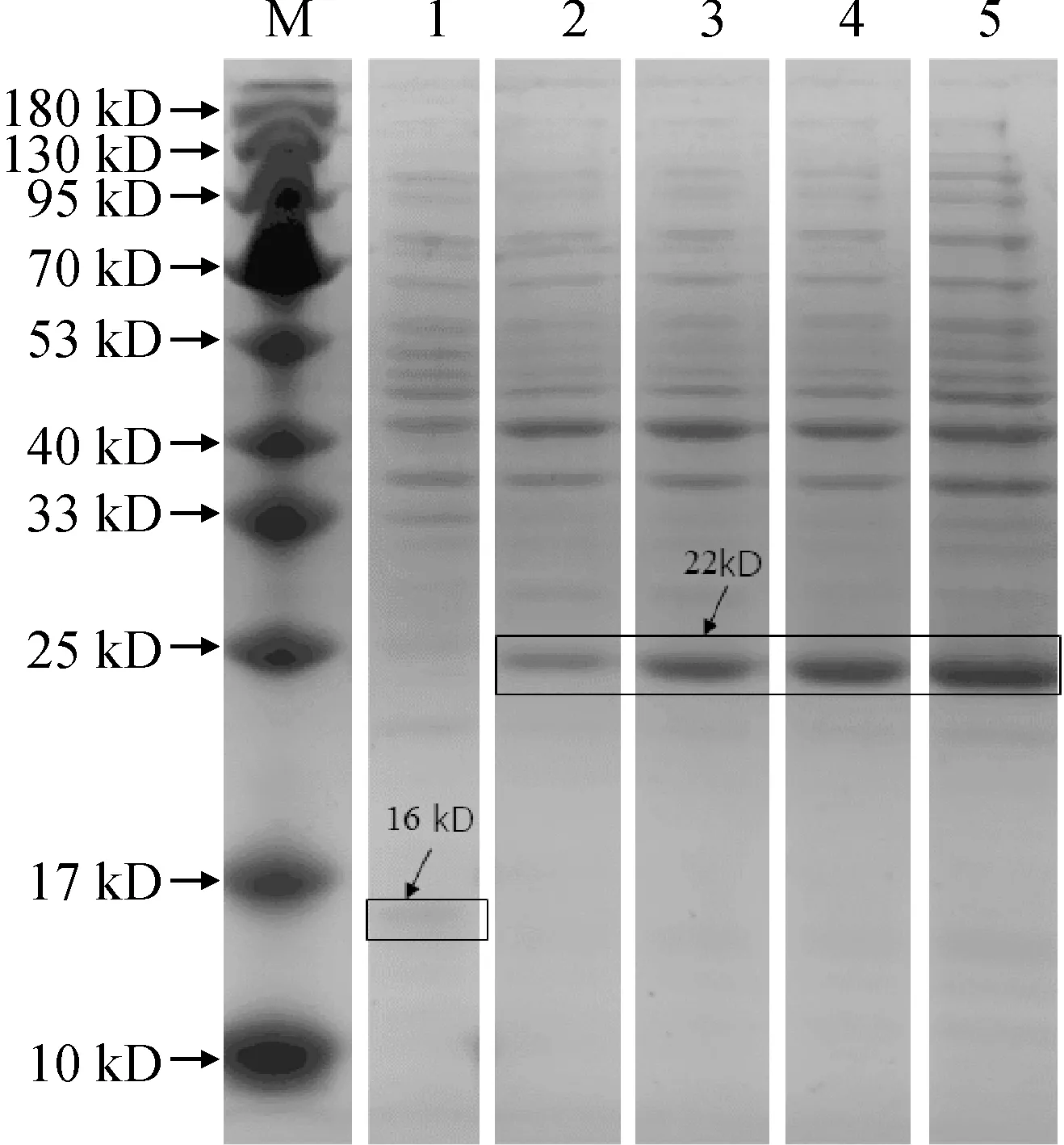
M. Marker;1.誘導6 h的BL21/pThiohis A;2-5為分別誘導1、2、4、6 h的BL21/LEA5圖4 BL21/pThiohis A和BL21/LEA5的原核表達M. Marker; 1. BL21/pThiohis A induced for 6 h; 2-5. BL21/LEA5 induced for 1, 2, 4 and 6 h, respectivelyFig.4 Prokaryotic expression of BL21/pThiohis A and BL21/LEA5
2.5 重組菌耐鹽性分析
為檢驗LEA5蛋白的重組菌對高鹽的耐受性,首先確定合適的鹽(NaCl或KCl)脅迫濃度。將空載體對照菌(BL21/pThiohis A)和重組菌(BL21/LEA5)進行IPTG誘導。再將等份的相同細胞密度的菌液分別涂布在添加外源NaCl(300、500和700 mmol/L)和KCl(500、700和900 mmol/L)的培養基上。結果顯示,當稀釋1 000倍時,BL21/pThiohis A和BL21/LEA5在300 mmol/L NaCl培養基平皿上菌斑數目超過1 000,二者的生長情況無明顯差別;在500 mmol/L NaCl培養基上,BL21/pThiohis A和BL21/LEA5的菌斑數目為10~600,二者的生長情況差別較小;在700 mmol/L NaCl培養基上,BL21/pThiohis A和BL21/LEA5菌斑數目為0~150,BL21/LEA5菌斑數目明顯多于BL21/pThiohis A。在700 mmol/L KCl脅迫下,BL21/pThiohis A和BL21/LEA5表現出與500 mmol/L NaCl相似的生長情況(表1)。因此,選擇500 mmol/L NaCl和700 mmol/L KCl作為脅迫培養基的鹽濃度。

表1 BL21/pThioHis A和BL21/LEA5在鹽脅迫條件下的菌斑數目
將BL21/pThiohis A和BL21/LEA5進行IPTG誘導,按0、10、100和1 000倍進行稀釋,將等份的菌液分別點在普通培養基和鹽脅迫(500 mmol/L NaCl和700 mmol/L KCl)培養基上,2 d后觀察平板上菌斑生長情況。結果顯示,在普通培養基上,不同稀釋倍數的BL21/pThiohis A和BL21/LEA5的菌斑數相近。在含500 mmol/L NaCl的培養基上,未稀釋和稀釋10倍的BL21/pThiohis A和BL21/LEA5菌斑數目較多,二者生長情況無明顯差別;在菌液稀釋100倍和1 000倍時,BL21/pThiohis A和BL21/LEA5菌斑數目差異較大, BL21/LEA5菌斑數目明顯多BL21/pThiohis A。在700 mmol/L KCl的培養基上,BL21/pThiohis A和BL21/LEA5表現出與500 mmol/L NaCl類似的生長情況。由此可知,LEA5蛋白的表達使大腸桿菌獲得了對高鹽(500 mmol/L NaCl和700 mmol/L KCl)的耐受能力(圖5)。

圖5 BL21/LEA5和BL21/pThioHis A在普通和鹽脅迫下的生長情況Fig.5 Growth situation of BL21/LEA5 and BL21/pThiohis A under normal and salt stress
3 討 論
順式作用元件是重要的分子開關,有助于了解基因表達調控信息。研究表明多數LEA家族基因啟動子具有大量與非生物脅迫的調控有關的元件[21]。ABA在植物逆境脅迫響應過程中發揮著信號分子的作用,ABRE元件是響應ABA的主要順式作用元件,MYC和MYB與ABA誘導基因的表達調控有關[22-23]。本研究中LEA5基因啟動子含有ABRE、MYC和 MYB等順式作用元件,表明LEA5基因在植物逆境脅迫中發揮重要作用。同時,LEA5啟動子區還含有茉莉酸甲酯響應元件(CGTCA-motif),推測LEA5基因可能對茉莉酸甲酯具有響應機制。此外,LEA5基因啟動子含有MYB結合位點(MYB binding site,MBS)、TGACG-motif、W-box等元件,W-box是WRKY轉錄因子的結合元件,而WRKY是植物參與各種生物與非生物脅迫應答的主要轉錄因子[24]。這些都表明,LEA5基因在纖枝短月蘚的抗逆性中起著非常重要的作用,有深入研究的價值。
熒光定量結果顯示,LEA5基因在不同時期及不同部位中皆有表達,且在無性世代的孢子體中表達量最高,有性世代的配子體次之,無性世代的配子體最低,揭示LEA5基因可能在無性世代孢子體的生長發育過程中起重要作用,其具體的機制還需進一步探究。
高鹽脅迫引起細胞脫水,損傷細胞內的蛋白和細胞膜。大量研究表明LEA基因的表達能夠提高植物的耐鹽性,有研究者將大豆的PM11、PM30、PM2基因分別轉化至大腸桿菌中,重組菌在高鹽脅迫下的存活率均明顯提高[25-26];還有研究表明SmLEA、SmLEA2基因的過表達提高了大腸桿菌對鹽脅迫的耐受性[27-28]。本研究結果顯示,重組菌(BL21/LEA5)的耐鹽性高于對照菌(BL21/pThioHis A),說明LEA5基因提高了大腸桿菌的耐鹽功能,這與前人的研究結果一致。
綜上所述,纖枝短月蘚LEA5基因啟動子區域含有與非生物脅迫的調控有關的順式作用元件,且在纖枝短月蘚不同時期及不同部位均有表達,LEA5基因的異源表達提高了大腸桿菌的耐鹽性,可見LEA5基因在纖枝短月蘚抗逆性中發揮重要功能。LEA5基因作為一個普遍存在于植物中的抗逆基因,與植物的抗逆性密不可分,將外源LEA5基因轉入常見的植物中,培育具有抗旱、耐鹽堿等逆境的植物新品種,這對實際的生產具有重要意義。
