基于ISSR與RAPD標記分析內蒙古赤芍的遺傳多樣性
2021-07-28 02:23:42杜亞楠孫淑英陳貴林
西北植物學報 2021年6期
杜亞楠,張 敏,孫淑英*,陳貴林
(1 內蒙古大學 生命科學學院,呼和浩特 010070;2 內蒙古自治區(qū)中蒙藥材規(guī)范化生產工程技術研究中心,呼和浩特 010070;3 牧草與特色作物生物學教育部重點實驗室,呼和浩特 010070)
遺傳多樣性是揭示物種適應環(huán)境的指標,也是物種與生俱來的特性[1]。了解植物種群的遺傳結構和變異程度,對于開展野生資源的保護和育種計劃具有重要意義[2]。在用于遺傳分析的分子工具中,隨機擴增多態(tài)性DNA(RAPD)[3]和簡單序列重復(ISSR)[4]標記與簡單序列重復(SSR)、序列相關擴增多態(tài)性(SRAP)和目標起始密碼子多態(tài)性(SCoT)等標記相比,具有節(jié)約成本、不需要序列信息、無需特定引物、操作簡便快速等優(yōu)點,被廣泛應用于植物遺傳多樣性研究中[5-7]。單一分子標記會使各項遺傳參數產生偏差,檢測多態(tài)性信息少,不能全面揭示物種的遺傳多樣性[8]。ISSR可以擴增基因組的內含子區(qū)域[9],RAPD可以同時擴增大量位點[10],這兩種分子標記的組合允許更高程度的基因組覆蓋。目前,ISSR和RAPD兩種分子標記結合廣泛用于荔枝[11]、竹子[12]、紅景天[13]、茴芹[14]、葉子花[15]及獐牙菜屬[16]等植物的遺傳變異、親緣關系和遺傳多樣性研究中。
赤芍是中國常用的大宗藥材,為毛茛科植物芍藥 (PaeoniaelactifloraPall.) 或川赤芍 (PaeoniaveitchiiLynch) 的干燥根,具有清熱涼血,散瘀止痛之功效[17]。中國赤芍廣泛分布于內蒙古、黑龍江和河北等省(區(qū)),其中內蒙古赤芍產量占據國內外供應市場近三分之一的份額,為重要的主產區(qū)[18]。隨著赤芍藥材需求日益增加,野生赤芍資源經過半個世紀的無序采挖,特別是進入21世紀之后,已日趨枯竭,市場主要以栽培赤芍為主。但目前生產中赤芍種質混雜,來源不清,各地間盲目引種更加重了混雜程度,根本無法保證藥材質量。為此,本研究從赤芍種群的遺傳多樣性角度解析其種群結構特征及親緣關系,為赤芍的引種選育、優(yōu)良種質挖掘和分子輔助育種提供理論依據。
1 材料和方法
1.1 實驗材料
供試赤芍樣本于2017年9月25日-10月8日分別采自內蒙古自治區(qū)、黑龍江省和吉林省等地,共計28個赤芍種群,包括12個野生種群和16個栽培種群。每個種群采集8株健康赤芍的根部組織,迅速裝入密封袋中,用硅膠干燥,帶回實驗室用去離子水清洗干凈并編號,放入-80 ℃冰箱儲存。實驗材料經內蒙古大學生命科學學院陳貴林教授鑒定為赤芍,樣品采集地信息詳見表1。
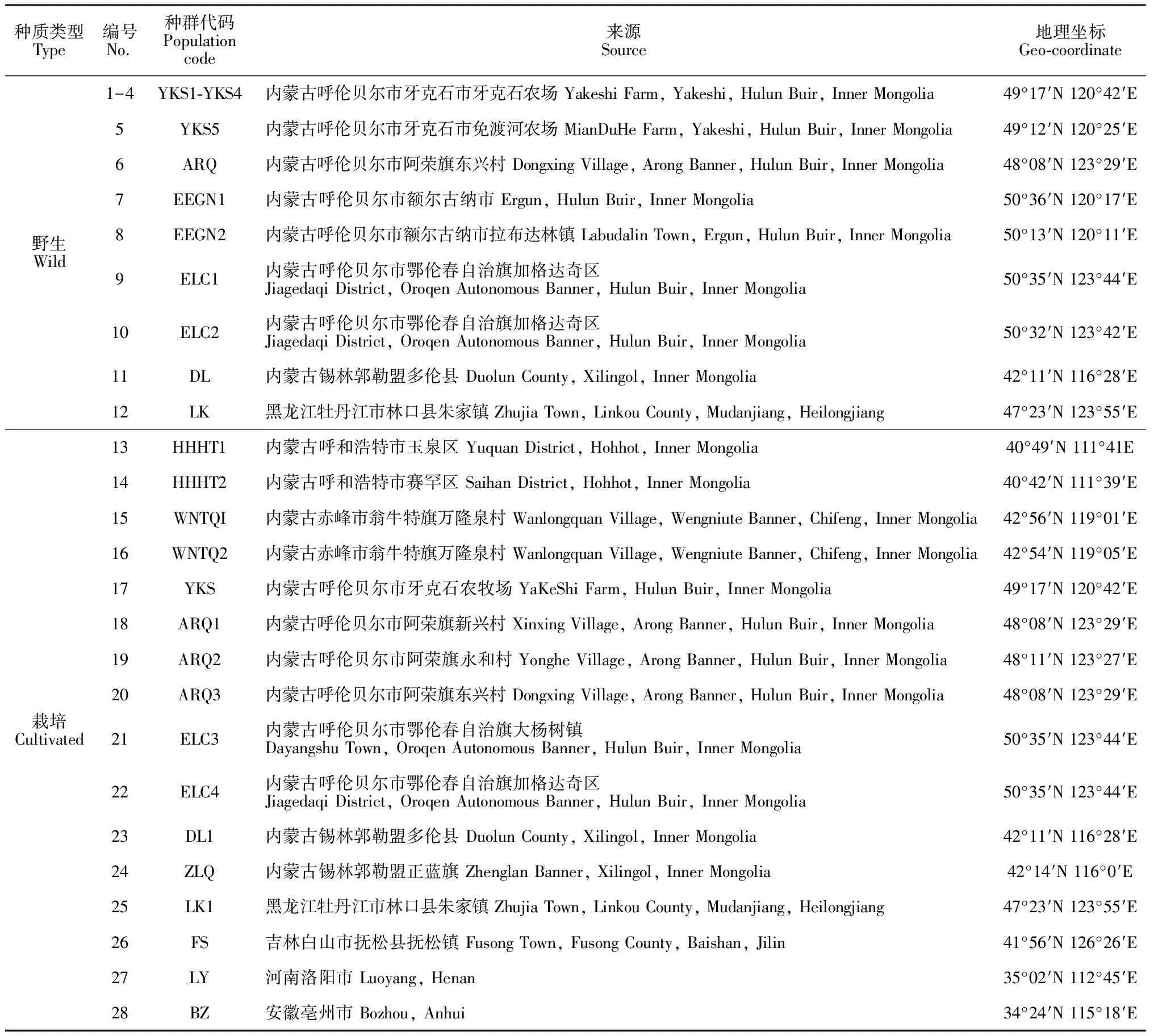
表1 赤芍種群采樣點地理信息
1.2 方 法
1.2.1 基因組DNA提取與質量檢測以單株赤芍的根組織樣本為單位,使用植物基因組DNA提取試劑盒(Tiangen)提取28個種群224株赤芍樣本的DNA。經1.3%的瓊脂糖凝膠電泳和NanoDrop核酸蛋白測定儀檢測DNA質量與濃度后,存于-20 ℃?zhèn)溆谩?/p>
1.2.2 引物篩選與其檢測本研究初步篩選的60條ISSR引物參照Zietkeiwitcz等[4]公布的序列,由上海生物工程技術公司合成。40條RAPD分子標記引物[19-20],由華大基因合成。用2個赤芍基因組DNA進行引物篩選,最后選取穩(wěn)定、清晰和多態(tài)性條帶豐富的14條ISSR引物和14條RAPD引物,并確定了其最佳退火溫度,用于本實驗研究。
ISSR分子標記擴增的方法是按照Liu等[21]方法進行,并做了一些小的修改。反應體系的總體積為25 μL,包含2 μL (20 ng/μL)DNA模板、1.5 μL(10 μmol/L)引物、9 μL ddH2O和12.5 μL 2×EsTaq Master Mix酶(TaKaRa)。PCR擴增程序為:94 ℃預變性5 min;94 ℃變性45 s,退火復性45 s,72 ℃延伸1.5 min,循環(huán)35次;72 ℃延伸10 min,4 ℃保存。
RAPD分子標記PCR反應體系與循環(huán)參數參照來明達等[19]報道的方法進行。反應體系總體積為25 μL,包含2 μL DNA模板(20 ng/μL)、1.5 μL(10 μmol/L)引物、10 μL ddH2O和11.5 μL 2×EsTaq Master Mix酶(TaKaRa)。PCR擴增程序為:94 ℃預變性5 min;94 ℃變性45 s,退火復性45 s,72 ℃延伸1.5 min,循環(huán)33次;72 ℃延伸7 min,4 ℃保存。1.3%瓊脂糖凝膠觀察擴增的PCR產物,通過凝膠成像系統拍照,記錄結果。
1.3 數據統計與分析
將擴增產物進行拍照,人工讀帶統計,在凝膠的相同遷移位置上將有條帶記為“0”,無條帶記為“1”,輸入到Excel中建立矩陣。利用PopGene32軟件對由野生和栽培的種質組成的兩組矩陣數據進行分組分析,計算多態(tài)性位點數(NPL)、多態(tài)性位點百分率(PPL)、觀測等位基因數(Na)、有效等位基因數(Ne)、Nei’s基因多樣性指數(He)、Shannon’s多樣性信息指數(I)、遺傳分化系數(Gst)和基因流(Nm)等遺傳系數;利用軟件NTsys Version 2.1進行UPGMA聚類分析。
2 結果與分析
2.1 擴增結果多態(tài)性分析
14條ISSR引物共擴增得到257條清晰可重復條帶,平均每條引物擴增出18.4條條帶,多態(tài)性條帶為251條,平均每條引物擴增的多態(tài)性條帶為17.9條,平均多態(tài)性比率為97.8%。其中ISSR UBC835引物擴增的條帶最多為25條,ISSR UBC860引物擴增的條帶最少為14條。
14條RAPD引物共擴增出215條條帶,平均每條引物擴增出15.4條帶,多態(tài)性條帶為209條,平均每條引物擴增的多態(tài)性條帶為14.9條,平均多態(tài)性比率為97.2%。其中RAPD S43、RAPD S1089和RAPD S1110引物擴增的條帶數最多為18條,RAPD S75和RAPD S1114引物擴增的條帶最少為13條(表2)。兩種標記擴增效率即多態(tài)帶百分率數值相近,證明篩選的引物擴增條帶清晰度和多態(tài)性較好,可用于本研究材料的遺傳多樣性分析。
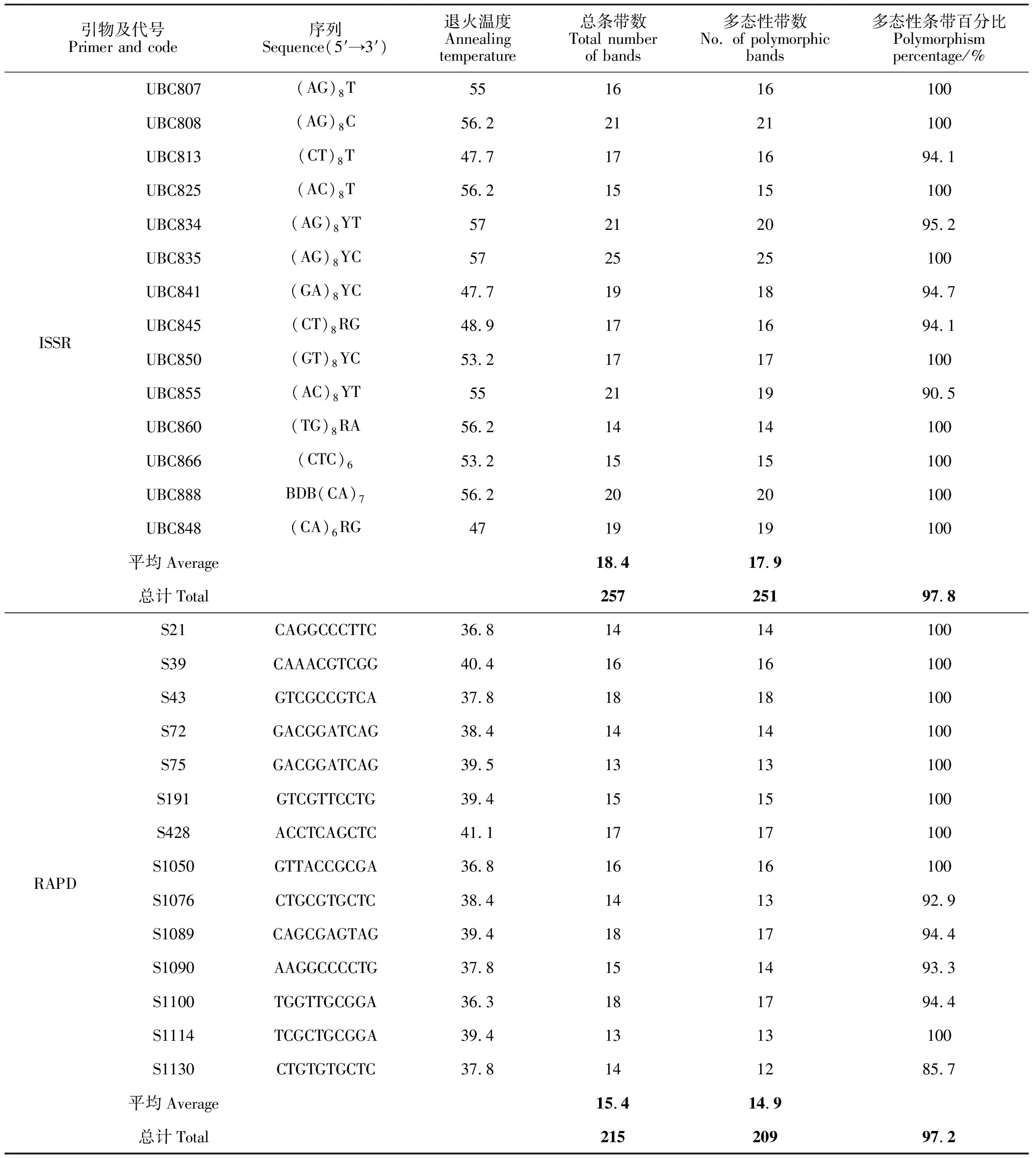
表2 引物序列及擴增條帶數
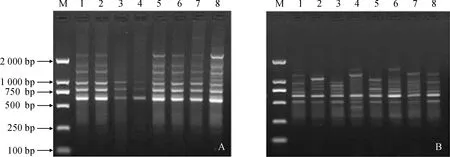
M.DL2000;1-8. 多倫(DL)種群8株植株編號圖1 引物ISSR UBC807(A)和RAPD S428(B)對多倫(DL)種群的擴增M.DL2000;1-8. No. of DL population’s 8 samplesFig.1 Effect of primers ISSR UBC807 and RAPD S428 for amplification of Duolun (DL) population of Paeoniae Radix Rubra
ISSR引物UBC807和RAPD引物S428對DL種群的擴增結果如圖1,擴增出的譜帶穩(wěn)定、清晰和多態(tài)性豐富,擴增片段長度約在 450~2 000 bp之間。由擴增結果發(fā)現,ISSR擴增出的條帶清晰且多于RAPD,對赤芍具有更強的檢測能力。
2.2 遺傳多樣性分析
根據ISSR、RAPD 兩種分子標記得到的0/1矩陣,通過 PopGene32軟件,假定遺傳平衡分別得到28個種群的遺傳多樣性參數,具體數據見表3。在28個種群中,DL(內蒙古自治區(qū)錫林郭勒盟多倫縣)種群各遺傳參數最高,ELC3(內蒙古自治區(qū)呼倫貝爾市鄂倫春自治旗大楊樹鎮(zhèn))種群最低。在野生種群中,DL種群遺傳多樣性水平仍然最高,對其生存的環(huán)境適應能力最強;而LK(黑龍江牡丹江市林口縣朱家鎮(zhèn))種群的遺傳多樣性水平最低,對環(huán)境適應能力最弱。而在栽培種群中,YKS種群(內蒙古呼倫貝爾牙克石市農牧場)遺傳多樣性最高,ELC3種群最低。同時ISSR分子標記得出的多態(tài)性位點數、多態(tài)性條帶百分率、Na、Ne、He、I等值均高于RAPD分子標記,在一定程度上表明樣品數相同、引物數相同時,ISSR檢測遺傳多樣性的能力較高,改進的ISSR可能是一種比RAPD更有效的研究赤芍植物系統發(fā)育的方法。

表3 28個赤芍種群的ISSR與RAPD遺傳多樣性參數分析
基于兩種標記對赤芍群體遺傳多樣性結果的比較(表4),28個種群的多態(tài)性位點平均百分比分別為 39.57%和35.95%;Na平均值分別為1.40和0.36;He平均值分別為0.15和0.13;I平均值分別為 0.22和0.19,由此可見赤芍種群存在較高的遺傳多樣性。根據反映研究對象遺傳多樣性水平高低指標的I和He,野生種群各遺傳多樣性指數平均值均高于栽培種群,即野生赤芍種群的遺傳多樣性水平高于栽培赤芍種群。
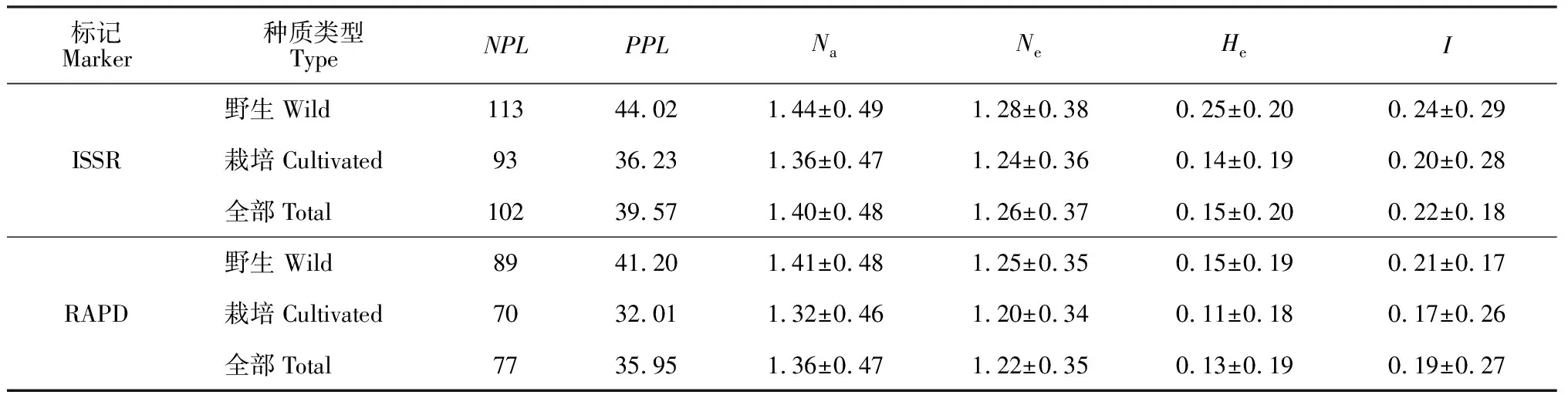
表4 赤芍群體ISSR和RAPD遺傳多樣性結果比較
2.3 遺傳分化分析
由表5可知,ISSR和RAPD得到28個赤芍種群的總遺傳多樣度(Ht)分別為0.32、0.27,種群內基因遺傳樣性(Hs)結果為0.15、0.12,那么群體之間的遺傳多樣性(Dst)大小為0.17、0.14。即遺傳多樣性在種群內占46%、47%,在種群間則為54%、53%,比較顯示赤芍種群間的多樣性水平大于種群內的。遺傳分化系數是反映種群分化程度的指標,Gst值在0至0.05之間,表示分化程度較弱;Gst值在0.05至0.15之間代表中等分化程度;Gst值在0.15至0.25之間表示較大的分化程度;大于0.25表示分化程度極大[22],兩種分子標記顯示赤芍種群間的遺傳分化程度較大, 且栽培赤芍種群間的遺傳分化程度大于野生種群。基因流(Nm)指生物個體在遷移過程中致使基因從一個種群到另一個種群的過程,Nm在0.44~0.47<1,說明種間產生了明顯的遺傳分化,28個赤芍種群之間的基因流較為豐富。同時觀察到野生種群比栽培種群Nm值高,且野生赤芍種群的遺傳分化主要發(fā)生在種群內,這可能是由遺傳漂變引起;而栽培赤芍種群之間的遺傳分化較為明顯,種群間的基因交流較少導致Nm值低。
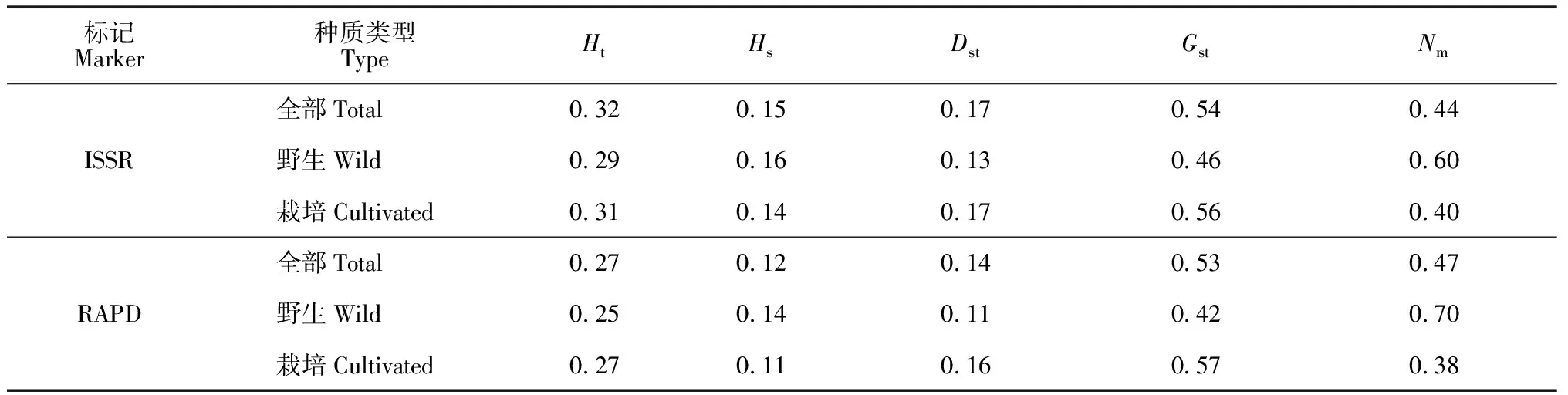
表5 基于ISSR和RAPD的赤芍種群的遺傳分化分析
2.4 聚類分析
根據ISSR的擴增結果得出0/1矩陣,經過NTSYS2.10軟件計算的遺傳距離構建聚類分析圖,遺傳距離變化范圍為0.115 1~0.343 8(圖2,A),28個赤芍種群可約在遺傳距離0.234處被分為5大類。第一類包括除了LK野生種群外的其余11個野生種群,還有YKS、ARQ1、ARQ2、WNTQ2和ZLQ 5個栽培種群組成;第二類包括LK、LK1和FS種群,這3個種群分別來自黑龍江省和吉林省;第三類只包括ELC3種群;第四類包括WNTQI、DL1、BZ、ELC4、LY、HHHT1、HHHT2種群;第五類只包括ARQ3種群。
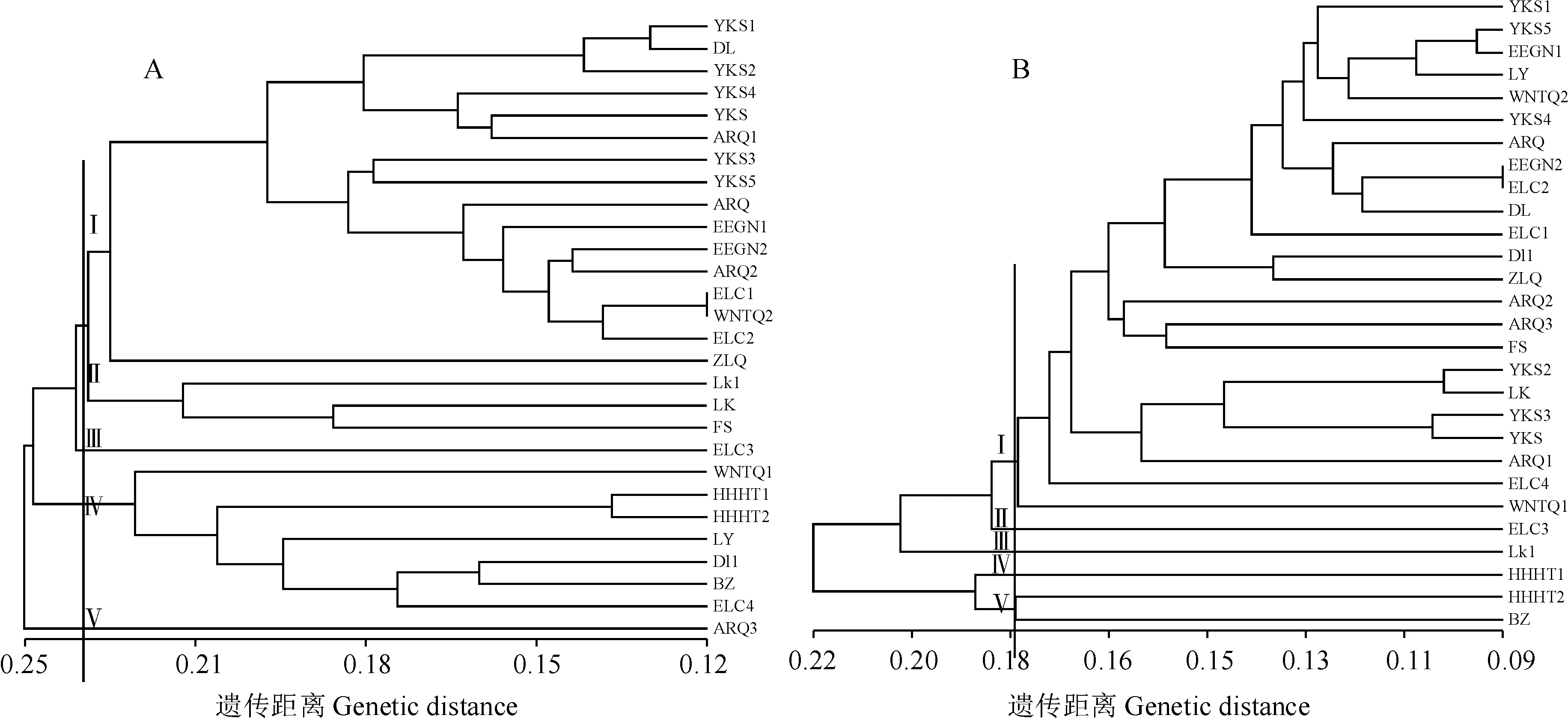
種群編號同表1圖2 基于ISSR (A)和RAPD (B)的UPGMA 聚類結果圖The population codes are same as Table 1Fig.2 UPGMA dendrogram based on ISSR (A) and RAPD (B) markers
基于RAPD分子標記與ISSR方法得到的結果相似,遺傳距離變化范圍為0.095 5~0.286 2,在遺傳距離0.179處同樣將28個赤芍種群分為5類(圖2,B)。第一類包括23個種群,其中包括了全部12個野生赤芍種群和大部分栽培赤芍種群,野生赤芍種群和栽培赤芍種群沒有完全分開;第二類由ELC3種群單獨聚為一類;第三類只有LK1種群;第四類HHHT1種群單獨聚為一類;第五類由HHHT2和BZ2組成。
兩種分子標記的聚類結果顯示YKS、ARQ1、ARQ2、WNTQ2和ZLQ等5個栽培種群總是與野生種群聚在一類,說明這5個栽培赤芍種群與野生赤芍種群的親緣關系較近;且均顯示栽培赤芍種群ELC3是獨立聚類的,該種群與其他栽培赤芍種群相比差異顯著,親緣關系較遠,具有獨特的遺傳背景。ISSR結果中LK、LK1和FS種群聚為一類,這3個種群分別來自黑龍江省和吉林省,證明種質聚類與地理分布有一定的相關性。但不完全一致,RAPD聚類中同是栽培種的安徽亳州BZ和內蒙古呼和浩特HHHT2種質聚在一起,以及ISSR聚類結果中ELC1和WNNTQ1種群、RAPD聚類結果中EEGN2和ELC2種群也未完全分開,這可能存在人為引種等原因導致它們在遺傳背景上相似。
3 討 論
分子標記被認為是最可靠、最有力的種質資源鑒定手段[23]。不同分子標記的結合使用可以根據各自的特點檢測基因組的不同部分,提高多態(tài)性位點的覆蓋率和均勻度,彌補一種分子標記帶來的局限性和弊端[24, 25]。在評價趕黃草[26]、野生艾蒿[27]和紅花[28]種質遺傳多樣性的研究中發(fā)現多種分子標記技術的使用可以互相驗證結果,從多方面、多角度地了解植物資源的遺傳信息,有效地揭示種質之間的遺傳多樣性。近年來, 國內外學者對芍藥屬進行了大量的遺傳多樣性研究,主要是探討牡丹的種間親緣關系[29, 30]和觀賞芍藥的群體遺傳關系[31-32],而關于藥用芍藥遺傳結構的研究多集中于白芍[33-34]。內蒙古作為赤芍的道地產區(qū),因其優(yōu)越的自然地理條件及退耕還林政策,赤芍的蘊藏量十分豐富,目前對該地區(qū)赤芍道地藥材的分子遺傳研究鮮有報道。本研究應用ISSR和RAPD兩種分子標記評估不同赤芍種群遺傳多樣性水平、遺傳變異以及種群之間的親緣關系,對于就地保護野生赤芍種質資源和內蒙古赤芍產業(yè)的發(fā)展具有重要意義。
在本研究中,ISSR和RAPD 兩種分子標記分析均發(fā)現內蒙古錫林郭勒盟多倫縣種群(DL)的赤芍遺傳多樣性水平最高,而近代以來一直推崇內蒙古多倫縣所產的野生赤芍為最佳道地藥材,使得多倫亦處于生產性赤芍資源瀕危狀態(tài)。因此,亟需就地建立自然保護區(qū),制定赤芍資源保護策略,盡快開展優(yōu)良種質基因庫的保護、研究及科學引種工作。通過歷代本草的記載發(fā)現,野生赤芍產區(qū)隨著野生資源的枯竭逐漸北移,目前主產區(qū)已轉至呼倫貝爾等地[35]。來自呼倫貝爾市的大部分赤芍野生種群具有較豐富的遺傳多樣性,是選育優(yōu)良藥用和觀賞品種的物質基礎;而栽培赤芍種群遺傳多樣性降低,可能是由于栽培赤芍所用繁殖材料主要取自野生資源并以分根的方式克隆繁殖,且藥農多從經濟價值角度出發(fā),主觀選擇根較粗壯的植株種植,使得群體的種質趨于單一;另外在栽培種群引種馴化過程中,參與馴化的野生種群偏小影響其基因頻率的變化,遺傳多樣性逐漸變窄。
兩種標記的聚類結果雖都將野生種群聚在一類,但栽培赤芍種群和野生赤芍群體之間有所交叉,未完全分開,可能是由于栽培赤芍的種源多來源于野生種,且初期栽培種多用種苗和芽頭無性繁殖[36]。在雪蓮果遺傳研究中認為,無性繁殖的營養(yǎng)性質會使其遺傳變異性變低[37],使得栽培種與野生種之間的性狀差異并不顯著,因而難以按產地聚類。獨自聚類的ELC3種群在所有種群中具有最低水平的遺傳多樣性,根據第四次中藥資源普查了解到內蒙古鄂倫春自治旗(縣)赤芍種植歷史悠久,ELC3可能是在人工馴化過程中形成了獨特的基因型種質,可作為優(yōu)質赤芍新品種進行進一步培育。對藥用植物而言,次生代謝物是臨床療效的物質基礎,豐富的化學成分多樣性使其具有更高的進化潛力和應用價值[38]。次生代謝物的合成和積累主要受內部遺傳基礎和外部環(huán)境因素的影響。因此,基于本研究基礎,擴大采樣區(qū)域,結合化學成分及環(huán)境因子對赤芍資源進行更加全面系統的分析和評價,以保存和挖掘優(yōu)良的基因型和化學型,為赤芍原位保護、遺傳改良和品種選育提供科學依據。
