脯氨酰寡肽酶抑制劑S17092 對脂質超載人肝細胞株LO2脂質沉積的改善作用及其機制
2021-08-25 10:16:10舍玲丁永年紀文靜阿孜古力阿不來提
山東醫藥 2021年21期
關鍵詞:模型
舍玲,丁永年,紀文靜,阿孜古力·阿不來提
新疆醫科大學第二附屬醫院消化科,烏魯木齊830011
非酒精性脂肪肝病(NAFLD)是一種由遺傳-環境-代謝應激相關因素所致的以肝細胞內脂肪堆積為主要表現的臨床病理綜合征[1]。NAFLD的發病機理尚不清楚,但已知脂質的合成和代謝在其發病中起著重要作用[2]。在肝臟中,固醇調節元件結合蛋白-1c(SREBP-1c)是脂質代謝的關鍵調控者,可調控與脂肪酸(FFA)、甘油三酯(TG)合成相關的靶基因,如硬脂酰輔酶A 去飽和酶(SCD1)、脂肪酸合成酶(FAS)、乙酰輔酶A 羧化酶1(ACC1)等,激活脂質合成的轉錄過程[3]。脯氨酰寡肽酶(POP)是一種可以裂解短肽中脯氨酸殘基羧基端肽鍵的絲氨酸蛋白酶,參與多肽的成熟和降解,可調控炎癥因子、細胞增殖、分化、凋亡等[4-6]。研究[7-8]發現,在單純脂肪變細胞及NAFLD 動物模型中POP 表達增高,提示POP在NAFLD 進展中起重要作用,但具體調節機制尚待進一步研究。2019年11月1日—2020年12月1日,本研究觀察了POP 抑制劑S17092 對脂質超載人肝細胞株LO2 脂質沉積的改善作用,并探討其作用機制。
1 材料與方法
1.1 細胞、試劑 人肝細胞株LO2購自購自中國科學院細胞庫。POP 抑制劑S17092 購自上海源葉生物科技有限公司,RPMI 1640 培養液購自Hyclone 公司,胎牛血清購自杭州四季青生物有限公司,棕櫚酸鈉(P9767)、油酸鈉(O7501)、油紅O 試劑盒購自Sig?ma-Aldrich 公司,GAPDH 抗體購自美國Santa Cruz公司,POP 抗體購自Abcam 公司,蛋白提取試劑盒、蛋白檢測試劑盒購自凱基生物公司,甘油三酯試劑盒購自中生北控生物技術有限公司,SDS-PAGE 凝膠制備試劑盒及組織細胞RNA 微量提取試劑盒購自索萊寶公司,SYBR Green 熒光實時定量PCR 試劑盒購自Life Technologies,RT-PCR 試劑盒購自TaKa?Ra 公 司,AcSDKP ELISA 試 劑 盒 購 自SPI Bio and CEA公司,PCR引物由生工生物工程公司合成。
1.2 脂質超載模型制備、分組及POP 抑制劑給予方法 參照文獻[9]方法,將LO2 細胞在含10%胎牛血清的1640 培養基中培養,每2~3 d 更換一次培養基,待細胞處于良好生長狀態且長至80%匯合時,將配置好的油酸和棕櫚酸(2∶1)的混合物按1 mmol/L加入培養液中,繼續培養24 h,建立脂質超載模型。綜合前期試驗結果及相關文獻[8],本研究選擇10μg/mL S17092 處理細胞。將LO2 細胞隨機分為3 組,對照組LO2 細胞用1%無FFA 的1640 培養基培養,模型組LO2 細胞制備脂質超載模型后再用1640 培養基培養,實驗組LO2 細胞制備脂質超載模型后再加入含S17092 的1640 培養液培養。各組細胞繼續培養24 h,用于后續實驗。
1.3 各組細胞內脂質沉積觀察 采用油紅O 染色法。將三組細胞用PBS 洗滌3 次并用4%多聚甲醛固定15 min,然后再用PBS 沖洗3 次,加入丙二醇試劑,室溫下脫水5 min,然后用新鮮稀釋的油紅O 溶液染色10 min,將細胞用60%異丙醇洗滌兩次,每次1 min,用水沖洗,并用蘇木精復染30 s,用PBS 洗滌后,顯微鏡下觀察細胞內脂質沉積情況。
1.4 各組細胞內TG 檢測 采用Bio-Rad 蛋白質測定法。取三組細胞,PBS洗滌兩次后,用0.25%胰酶消化,添加100μL 細胞裂解緩沖液孵育10 min 后離心5 min,收集上清液,按照TG 測定試劑盒說明書要求檢測細胞內TG。實驗重復3次,取平均值。
1.5 各 組 細 胞 內POP、SREBP-1c、FAS、ACC1、SCD1 mRNA 檢測 采用實時熒光定量PCR 法。取三組細胞,提取RNA 后反轉錄合成cDNA,以cDNA為模板,以GAPDH 為內參,配置20 μL 的PCR 反應液,于實時熒光定量PCR 儀上進行PCR 反應。PCR反應條件:50 ℃2 min、95 ℃2 min 預變性,95 ℃15 s、60 ℃15 s、72 ℃1 min,40個循環。引物序列如下:POP 正 向 引 物 為5′-CATCTCCCAAGAGGCT?GACTA-3′,反向引物為5′-GGGCAATAACACAAC?CAAAGA-3′;SREBP-1c 正 向 引 物 為 5′-GA?CAGCCCAGTCTTTGAGGA-3′,反向引物為5′-CAG?GACAGGCAGAGGAAGAC-3′;FAS 正向引物為5′-TTCCGAGATTCCATCCTACG-3′,反 向 引 物 為5′-AGGCTCACAAACGAATGGAC-3′;ACC1 正向引物為5′-TCACACCTGAAGACCTTAAAGCC-3′,反向引物 為5′-AGCCCACACTGCTTGTACTG-3′;SCD1 正向引物為5′-TTCCTACCTGCAAGTTCTACACC-3′,反 向 引 物 為5′-CCGAGCTTTGTAAGAGCGGT-3′;GAPDH 正 向 引 物 為 5′-GAAGGTGAAGGTCG?GAGTC-3′,反 向 引 物 為5′-GAAGATGGTGAT?GGGATTTC-3′。以2-ΔΔCt法表示細胞中目的mRNA的相對表達量。
1.6 統計學方法 采用SPSS21.0 統計軟件。計量資料以±s表示,多組間比較用單因素方差分析,兩兩比較用LSD-t檢驗。P<0.05 為差異有統計學意義。
2 結果
2.1 各組細胞內脂質沉積情況比較 油紅O 染色結果顯示,與對照組相比,模型組LO2細胞內含有大量的紅染顆粒,而實驗組較模型組紅染顆粒相對較少,見圖1。
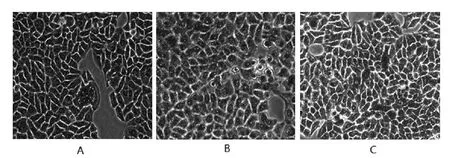
圖1 顯微鏡下觀察油紅O染色的LO2細胞(×200)
2.2 各組細胞內TG 水平比較 對照組細胞內TG水平為(1.995 ± 0.221)pmole/μL,模型組細胞內TG 水平為(2.910 ± 0.030)pmole/μL,實驗組細胞內TG 水平為(2.573±0.113)pmole/μL,組間相比,P均<0.05。
2.3 各 組 細 胞 內POP、SREBP-1c、FAS、ACC1、SCD1 mRNA 相對表達量比較 各組細胞內POP、SREBP-1c、FAS、ACC1、SCD1 mRNA 相對表達量比較見表1。由表1 可知,與對照組相比,模型組、實驗 組 細 胞 中POP、SREBP-1c、FAS、ACC1、SCD 1 mRNA 相對表達量明顯升高(P均<0.05);與模型組 相 比,實 驗 組 細 胞 中POP、SREBP-1c、FAS、ACC1、SCD1 mRNA 相對表達量明顯降低(P均<0.05)。
表1 各組細胞內POP、SREBP-1c、FAS、ACC1、SCD1 mRNA相對表達量比較(±s)

表1 各組細胞內POP、SREBP-1c、FAS、ACC1、SCD1 mRNA相對表達量比較(±s)
注:與對照組相比,*P<0.05;與模型組相比,#P<0.05。
組別實驗組模型組對照組POP mRNA 1.369±0.0819*#1.595±0.040*1.060±0.021 SREBP1c mRNA 1.381±0.083*#2.626±0.066*1.043±0.032 FAS mRNA 1.639±0.113*#2.955±0.095*1.051±0.019 ACC1 mRNA 1.376±0.045*#1.821±0.066*1.062±0.036 SCD1 mRNA 1.311±0.019*#1.869±0.109*1.073±0.014
3 討論
近年來,隨著肥胖等代謝性疾病的增加,NAFLD的發病率明顯增高。研究[11]發現,NAFLD是終末期肝病和肝癌的重要病因,即使是肝臟輕度脂肪變性,也會增加71%的死亡風險,而高循環FFA濃度可通過破壞NAFLD 患者的脂質代謝而加重肝臟脂肪的積累[12],因此研究FFA 如何影響代謝調節將進一步增進我們對NAFLD 發病機制的了解。本研究的目的是在FFA 誘導的脂質超載細胞模型中研究POP抑制劑對NAFLD 的影響,這可能有助于闡明與疾病進展有關的機制。在正常人和NAFLD 患者中,棕櫚酸和油酸都是肝甘油三酯中最豐富的FFA。根據GOMEZ-LECHON 等的報告,載有含油酸/棕櫚酸(比例為2∶1)的FFA(1 mmol/L)混合物的肝細胞模擬了人類的肝臟慢性脂肪變性[9]。因此,我們通過將LO2 細胞與1 mmol/L FFA 一起孵育,建立了細胞脂肪變性的脂質超載模型,油紅O 染色顯微觀察可見細胞內脂質液泡中大量脂質沉積證實了建模成功。
目前研究[13-14]發現,POP 與糖尿病、多發性硬化、肝硬化、抑郁癥、神經退行性疾病和癌癥相關,在肝臟中發現POP 可促進肝臟的再生和修復。此外有研究[13]表明,POP 可作為肝硬化嚴重程度的評估指標。而ZHANG 等[7-8]研究發現,在高脂飲食誘導的NAFLD模型中POP表達上調,抑制POP可降低肝脂肪變性的嚴重性和炎癥反應,而抑制POP 對肝臟炎癥的保護作用與基質金屬蛋白酶(MMPs)和脯氨酸-甘氨酸-脯氨酸(PGP)的產生受抑制以及嗜中性粒細胞浸潤的抑制有關。而另一項細胞實驗[15]表明,POP 過表達可導致Smad7 蛋白和PPAR-γ 上調,減弱大鼠肝星狀細胞的活化,進而延緩肝纖維化。目前POP 在肝臟脂肪變性過程中的機制尚未闡明。一般認為,脂肪肝是由肝臟攝取FFA、TG 合成和排泄之間的不平衡引起的,與脂質代謝相關基因失調密切相關。本研究發現,與對照組相比,模型組POP mRNA 相對表達量更高,而實驗組則表現出細胞脂肪變性減輕。SREBP-1c 是調節肝臟新生脂肪形成的最重要轉錄因子,可以通過調節內源性TG、膽固醇、FFA 等合成所需的酶的表達來維持脂質的動態平衡。SREBP-1c 及其靶基因的異常可引起胰島素抵抗、糖尿病、心臟功能障礙、血管并發癥和肝脂肪變性等一系列代謝性疾病。研究[16]發現,NAFLD 患者肝臟內的SREBP-1c 含量明顯升高,表明SREBP-1c在NAFLD 發生進展中起重要作用。本研究中,FFA 刺激下的L02 細胞中SREBP-1c 基因表達顯著上升,其調控的靶基因FAS、ACC1、SCD1 的表達同步上調,細胞內TG 含量亦增加,并伴有肝細胞內大量脂滴生成,表明SREBP-1c基因表達的上調啟動了相關脂質生成的級聯過程,促進NAFLD 的發生及進展。而與模型組比較,實驗組肝LO2 細胞抑制POP 后,內源性長鏈脂肪酸合成的關鍵基因SREBP-1c及其靶向基因FAS、ACC1、SCD1 mRNA相對表達量亦顯著降低,細胞內TG含量和脂滴的生成減少。這些發現表明,在L02 細胞中進行FFA 處理導致脂質蓄積可能是由POP介導的,而SREBP-1c及其靶基因FAS、ACC1、SCD1 相對表達量升高是POP誘導脂肪變性的機制的重要組成部分。
總之,POP 抑制劑S17092 可改善脂質超載LO2細胞的脂質沉積,其機制可能與抑制SREBP-1c及其靶基因的表達有關。細胞中POP 表達與脂肪合成相關基因之間的密切關系可能是治療干預NAFLD的潛在目標。然而,POP 促進肝脂肪變性的確切機制仍有待闡明。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
