臭椿酮對植物種子萌發(fā)及相關(guān)生理生化指標(biāo)的影響
2021-08-26 10:51:34田岐震魏少鵬姬志勤
農(nóng)藥學(xué)學(xué)報 2021年4期
關(guān)鍵詞:植物
田岐震, 魏少鵬,2, 姬志勤*,,2
(1. 西北農(nóng)林科技大學(xué) 植物保護(hù)學(xué)院,陜西 楊凌 712100;2. 陜西省植物源農(nóng)藥研究與開發(fā)重點(diǎn)實(shí)驗(yàn)室,陜西 楊凌 712100)
臭椿Ailanthus altissima是原產(chǎn)于東亞的苦木科臭椿屬植物,其根皮和果實(shí)為傳統(tǒng)中藥[1]。迄今為止,已從臭椿中分離鑒定了數(shù)十個苦木苦味素類化合物,這些化合物表現(xiàn)出抗病毒、抗腫瘤、抗瘧及抗菌等多種藥理活性[2-4]。1959年,Mergen首次報道臭椿提取物具有除草活性[5];1995年,臭椿提取物中的除草活性成分被分離鑒定為臭椿酮[6]。2003年,De Feo等對臭椿根皮中的除草活性成分進(jìn)行了系統(tǒng)分離,從中得到臭椿酮、ailanthinone、chaparrine和ailanthinol B 4個具有除草活性的苦木苦味素類化合物,其中臭椿酮的活性最強(qiáng)[7]。2011年,Pedersini等比較了臭椿莖皮不同溶劑提取物的除草活性,發(fā)現(xiàn)其活性與臭椿酮的濃度呈正相關(guān)[8]。Heisey等報道了臭椿提取物對17種雜草的田間防除效果,發(fā)現(xiàn)提取物對其中13種供試雜草具有很好的防效[9]。2019年,Demasi等研究了臭椿酮苗前土壤處理防治雜草的田間持效期,發(fā)現(xiàn)其在低有機(jī)質(zhì)含量土壤中的持效期可達(dá)20~30 d,但在高有機(jī)質(zhì)含量土壤中的降解較快[10]。近年來,國內(nèi)外對臭椿酮 (或臭椿提取物)的除草活性進(jìn)行了較多的研究,證明臭椿酮具有開發(fā)成植物源除草劑的潛在價值,然而有關(guān)臭椿酮除草作用機(jī)制的研究鮮有報道。本研究測定了臭椿酮對植物種子萌發(fā)及相關(guān)生理生化指標(biāo)的影響,旨在為進(jìn)一步研究臭椿酮的除草作用機(jī)制提供參考。
1 材料與方法
1.1 供試藥劑及植物
臭椿酮 (ailanthone,純度95%,CAS: 981-15-7),本課題組從臭椿根皮中分離得到[11]。
油菜Brassica napus、小麥Triticum aestivum、馬齒莧Portulaca oleracea、苘麻Abutilon theophrasti、稗草Echinochloa crusgalli、反枝莧Amaranthus retroflexus、狗尾草Setaria viridis及馬唐Digitaria sanguinalis,以上供試作物及雜草的種子均采自西北農(nóng)林大學(xué)北校區(qū)農(nóng)場。
1.2 種子萌發(fā)活性測定
采用平皿法測定臭椿酮對8種供試植物種子萌發(fā)的影響[7]。先用少量二甲基亞砜溶解臭椿酮,再用體積分?jǐn)?shù)0.1%的Tween 80分別稀釋成1.56、3.13、6.25、12.50、25.00和50.00 μg/mL 的供試溶液,二甲基亞砜在所有供試溶液中的最終體積分?jǐn)?shù)不超過0.5%。先用體積分?jǐn)?shù)1%的次氯酸鈉溶液對種子進(jìn)行表面消毒,再用去離子水沖洗3次。將50粒種子均勻地放在培養(yǎng)皿 (9 cm) 的濾紙上,每個培養(yǎng)皿中加入5.0 mL上述臭椿酮供試溶液。以含體積分?jǐn)?shù)0.5%的二甲基亞砜和0.1% Tween 80的水溶液作為溶劑對照。種子在25 ℃、16 h/8 h (光/暗) 條件下培養(yǎng)4 d。分別于24、48、72和96 h記錄萌發(fā)種子的數(shù)量 (以根長2 mm為萌發(fā)標(biāo)準(zhǔn))。所有試驗(yàn)重復(fù)3次。
1.3 種子吸水量測定
參照文獻(xiàn)方法進(jìn)行[12]。在鋪有雙層濾紙的培養(yǎng)皿 (9 cm) 中分別加入5.0 mL質(zhì)量濃度為1.56、3.13、6.25、12.50和25.00 μg/mL 的臭椿酮溶液,分別放入油菜 (1.0 g) 和小麥 (2.0 g) 種子,在25 ℃恒溫條件下分別培養(yǎng)4、8、12和16 h。將種子從溶液中取出,吸干水分,稱重,計算吸水量 (最終質(zhì)量減去初始質(zhì)量)。以含體積分?jǐn)?shù)為0.5%的二甲基亞砜和0.1%的Tween 80的水溶液作為溶劑對照。每個處理重復(fù)3次,計算平均值。
1.4 幼苗生長的生理生化指標(biāo)的測定
根據(jù)預(yù)試驗(yàn)確定臭椿酮抑制幼苗生長中濃度。將萌發(fā)的油菜和小麥種子分別置于0.50 μg/mL和2.00 μg/mL臭椿酮溶液中,在25 ℃的光照培養(yǎng)箱中培養(yǎng) (光/暗 = 12 h/12 h)。以含體積分?jǐn)?shù)為0.5%的二甲基亞砜和0.1%的Tween 80的水溶液作為溶劑對照。當(dāng)對照組種子胚芽長到約1.0 cm時,分別收集胚芽和胚根 (各0.50 g),置于預(yù)冷的研缽中,加入0.05 mol/L預(yù)冷的磷酸鹽緩沖液1.0 mL (pH 7.8),然后將胚芽和胚根在冰上磨成粉末,分別提取超氧化物歧化酶 (SOD)、過氧化氫酶 (CAT)、過氧化物酶 (POD) 和總可溶性蛋白(TSP)[13]。當(dāng)對照組種子胚芽長到約1.0 cm時,分別收集胚芽和胚根 (0.10 g),加入蒸餾水5~10 mL,煮沸30 min,提取兩次,提取液過濾入25.00 mL容量瓶,用蒸餾水定容,提取總可溶性糖 (TSS)[14]。所有粗提液于4 ℃保存?zhèn)溆谩?/p>
采用氮藍(lán)四唑光還原法[14]測定SOD活性;采用愈創(chuàng)木酚法[14]測定POD活性;采用高錳酸鉀滴定法[14]測定CAT活性;采用3,5-二硝基水楊酸法[14]、考馬斯藍(lán)G-250染色法[14]分別測定TSS和TSP含量。每處理重復(fù)3次。
1.5 對植物細(xì)胞有絲分裂的影響
參照文獻(xiàn)方法進(jìn)行[15]。將油菜種子置于含水濾紙上培養(yǎng),待根長為1.0 cm時,挑選生長一致的幼苗移至質(zhì)量濃度分別為0.01、0.05、0.10、0.50和1.00 μg/mL的臭椿酮溶液中繼續(xù)培養(yǎng)24 h。取根尖0.5 cm部分,浸入卡諾氏固定液中固定6 h。取出根尖用去離子水沖洗至無醋酸氣味,接著將根尖浸入1 mol/L的鹽酸溶液,在60 ℃下孵育10 min。取出根尖用去離子水洗滌3次,置于載玻片上,用接種針將根尖搗碎,加入卡寶品紅染色10 min。放上蓋玻片,用鉛筆輕輕敲擊,使細(xì)胞均勻分布。光學(xué)顯微鏡下進(jìn)行觀察和計數(shù)。有絲分裂指數(shù)和畸形指數(shù)分別按公式 (1) 和 (2)計算。
式中:A,有絲分裂指數(shù);C,分裂細(xì)胞數(shù);D,頂端分生組織細(xì)胞總數(shù);B,畸形指數(shù);E,分裂異常數(shù)。
1.6 活性氧含量的測定
參考文獻(xiàn)方法[16]并略作修改。將油菜種子置于含水濾紙上催芽,待根長為1.0 cm時,挑選長勢相近的油菜幼苗,浸入0.10、1.00和10.00 μg/mL的臭椿酮溶液,于25 ℃黑暗培養(yǎng)12 h。以體積分?jǐn)?shù)為0.1%的二甲基亞砜溶液作為溶劑對照。每個處理設(shè)3個重復(fù)。將幼苗分別轉(zhuǎn)置于3,3-二氨基聯(lián)苯胺 (DAB) 染色液和氯化硝基四氮唑藍(lán) (NBT)染色液中,于25 ℃黑暗培養(yǎng)箱中浸染2 h。取出幼苗觀察拍照,用Image J軟件進(jìn)行量化處理。按公式(3)計算染色強(qiáng)度。
式中:R,染色強(qiáng)度;F,未染色根尖灰度值;H,染色根尖灰度值。
1.7 細(xì)胞死亡率測定
參考文獻(xiàn)方法[15]并略作修改。將油菜種子置于含水濾紙上催芽,待根長為1.0 cm時,挑選長勢相近的油菜幼苗,浸入0.10、1.00和10.00 μg/mL的臭椿酮溶液。以體積分?jǐn)?shù)為0.1%二甲基亞砜溶液為對照,于28 ℃黑暗培養(yǎng)24、48和72 h,幼苗煮沸30 min作為死亡對照,將整株油菜浸入2.50 μg/mL的伊文思藍(lán)水溶液中,于旋轉(zhuǎn)搖床上25 ℃孵育30min,取出植株用去離子水洗滌,除去未結(jié)合的染料,并觀察拍照。每處理重復(fù)3次。運(yùn)用Image J對照片進(jìn)行量化處理。按公式(4)計算細(xì)胞相對死亡率。
式中:X,細(xì)胞相對死亡率;M,0.1%二甲基亞砜組灰度值;L,處理組灰度值;K,CK組灰度值;Z,煮沸對照組灰度值。
1.8 數(shù)據(jù)處理
采用SPSS 23.0軟件計算臭椿酮對種子萌發(fā)抑制IC50值,使用鄧肯氏新復(fù)極差法 (P< 0.05) 進(jìn)行差異顯著性分析;使用Image J量化處理染色強(qiáng)度;使用Excel 2010完成圖表繪制;使用Photoshop 6.0處理圖片。
2 結(jié)果與討論
2.1 臭椿酮對8種植物種子萌發(fā)的影響
從表1可以看出,臭椿酮對油菜、稗草、苘麻、狗尾草和馬唐的種子萌發(fā)具有較強(qiáng)的抑制作用,其IC50值分別為3.74、9.86、9.71、3.87和3.76 μg/mL。臭椿酮對小麥和反枝莧種子萌發(fā)也有較強(qiáng)的抑制作用,其IC50值分別為24.70和32.60 μg/mL。在供試濃度下,臭椿酮對馬齒莧種子萌發(fā)的抑制作用較弱。從結(jié)果可以看出,臭椿酮的除草活性在單子葉和雙子葉植物間無選擇性,但是個別雜草對臭椿酮的敏感性較低。臭椿酮對油菜和小麥的種子萌發(fā)有明顯抑制作用,有文獻(xiàn)也報道了臭椿提取物在采用莖葉噴霧法防治雜草時對番茄、白蘿卜、黃豆、向日葵、玉米、燕麥和南瓜有嚴(yán)重的藥害[9],因此不建議將其用于大田作物及蔬菜生產(chǎn)中,但在園藝作物中作為一種非選擇性除草劑具有潛在的應(yīng)用價值,相關(guān)應(yīng)用技術(shù)值得進(jìn)一步研究。
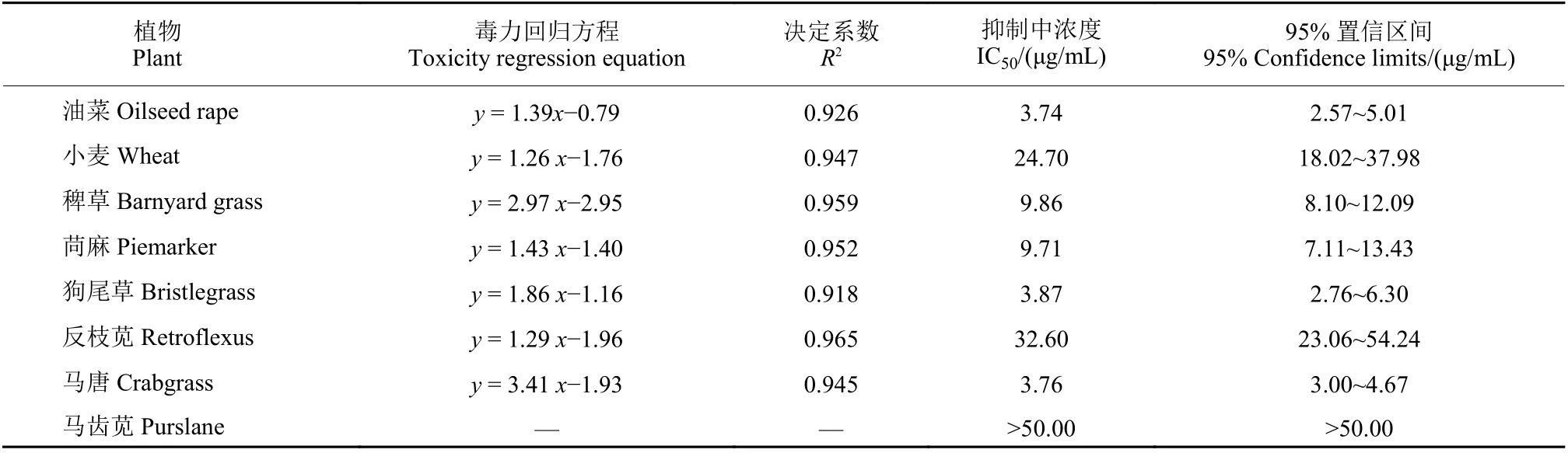
表1 臭椿酮對8種植物種子萌發(fā)的影響Table 1 Effects of ailanthone on seed germination of eight plants
2.2 臭椿酮對油菜和小麥種子吸水量的影響
萌發(fā)是種子從靜止干燥狀態(tài)到代謝活躍狀態(tài),然后生長成幼苗的過程[17]。這一過程始于干燥種子對水分的吸收,外源化合物可以抑制種子蛋白酶的活性,造成種子吸水能力下降,最終導(dǎo)致種子不能正常萌發(fā)[12,18]。臭椿酮處理后的油菜和小麥種子吸水量如圖1所示。可以看出,不同濃度臭椿酮處理的種子在吸水能力上并無明顯差異,與空白對照相比也沒有顯著差異,這說明臭椿酮并未影響種子的吸水能力,由此可以推測臭椿酮不影響種子蛋白酶的活性。
2.3 臭椿酮對幼苗生長的影響
SOD在保護(hù)細(xì)胞免受超氧化物自由基損傷方面發(fā)揮重要作用,它催化超氧化物自由基的降解生成過氧化氫[19]。細(xì)胞內(nèi)過氧化氫水平受多種酶調(diào)控,CAT是多酶體系中的一種[20]。POD是植物中的抗氧化酶,通過將過氧化氫還原為水來催化多種底物的氧化[21]。SOD、CAT和POD活性的增加是植物組織提高抗氧化能力的表現(xiàn)。
對油菜和小麥胚芽和胚根中SOD、CAT和POD的酶活性測定結(jié)果 (圖2) 表明:溶劑對照處理油菜胚根SOD活性為106.27 U/g, 臭椿酮處理后SOD活性增加至173.56 U/g。同樣,用臭椿酮處理后,小麥胚芽中SOD活性從1 851.43 U/g增加到2 159.61 U/g,小麥胚根中SOD活性從167.50 U/g 增加到441.54 U/g。臭椿酮也導(dǎo)致了CAT活性的輕微增加,小麥胚芽和胚根的CAT活性值從1.79 mg/(g?min)增加到2.20 mg/(g?min),胚根的CAT活性值從0.41 mg/(g?min)增加到0.56 mg/(g?min);處理過的油菜和小麥幼苗POD活性也高于對照組。
與對照組相比,臭椿酮提高了油菜和小麥胚中TSS和TSP的含量,說明低劑量的臭椿酮能提高植物的整體代謝水平。
活性氧 (ROS) 的產(chǎn)生和消除是一個動態(tài)平衡,在植物體內(nèi)活性氧的消除主要依賴于細(xì)胞的抗氧化酶系,當(dāng)細(xì)胞內(nèi)產(chǎn)生的活性氧增加時,為維持細(xì)胞的正常生長,植物會提高抗氧化酶活性消除活性氧[22]。上述生理指標(biāo)試驗(yàn)證實(shí)臭椿酮的存在可以提高植物的抗氧化能力,但臭椿酮是否會誘導(dǎo)ROS的積累仍需通過活性氧染色試驗(yàn)進(jìn)行探究。
2.4 臭椿酮對細(xì)胞有絲分裂的影響
通過對油菜根分生組織細(xì)胞的有絲分裂指數(shù)分析,評價了臭椿酮對油菜根分生組織細(xì)胞分裂的影響。暴露于臭椿酮 (0.01、0.05、0.10、0.50和1.00 μg/mL) 24 h后,較高濃度的臭椿酮 (0.50和1.00 μg/mL) 明顯抑制了油菜根的生長 (圖3 Ⅰ),其中1.00 μg/mL的臭椿酮抑制率為40.60%。處理組根尖分生區(qū)有絲分裂指數(shù)均低于對照組,且臭椿酮對油菜細(xì)胞分裂的抑制作用呈劑量依賴性 (圖3 Ⅱ)。暴露于1.00 μg/mL的臭椿酮后,有絲分裂指數(shù)僅為0.48%,而對照組為5.85%。此外,臭椿酮還能導(dǎo)致畸形細(xì)胞的形成,其中0.10 μg/mL的臭椿酮處理后畸形指數(shù)最高,約為0.45% (圖3 Ⅱ、Ⅲ)。
2.5 臭椿酮對植物組織中活性氧積累的影響
活性氧是植物進(jìn)行代謝的副產(chǎn)物,在生長發(fā)育過程中可以作為信號分子參與調(diào)控,但是活性氧的過度積累會導(dǎo)致脂質(zhì)過氧化反應(yīng),從而對植物的生長造成嚴(yán)重的損傷[23]。植物體內(nèi)的活性氧主要包括超氧陰離子 (O2?)、過氧化氫 (H2O2) 和羥基自由基 (?OH) 等。
由圖4可以看出,油菜根浸入臭椿酮后,不僅根長受到了抑制,根部的活性氧含量也大幅升高,且其含量隨著臭椿酮質(zhì)量濃度的增加而升高,在10.00 μg/mL臭椿酮劑量下,油菜根長幾乎沒有增長,但是過氧化氫和超氧陰離子含量分別是溶劑對照的2.53倍和2.08倍。由試驗(yàn)結(jié)果得知,臭椿酮的存在可以在短時間內(nèi)誘導(dǎo)植物體內(nèi)大量活性氧的累積。抗氧化酶活性測定結(jié)果表明,在臭椿酮的影響下,SOD、CAT和POD的活性都呈升高的趨勢,這與臭椿酮可以在短時間內(nèi)誘導(dǎo)大量活性氧積累相關(guān),體內(nèi)活性氧含量升高會損害細(xì)胞正常生命活性,機(jī)體便會調(diào)節(jié)抗氧化酶的活性來消除活性氧。
2.6 細(xì)胞死亡率測定
通過圖5 (Ⅰ、Ⅱ、Ⅲ) 可以看出,染色較深的是根毛茂盛部位和根尖,對根尖區(qū)域的相對死亡率測定發(fā)現(xiàn),在臭椿酮相同質(zhì)量濃度下,根尖細(xì)胞的相對死亡率并沒有隨著時間的推移而產(chǎn)生較大變化。1.00 μg/mL臭椿酮處理24、48和72 h的細(xì)胞相對死亡率分別為8.56%、9.58%和10.23%,但是,在同一時間段,根尖細(xì)胞相對死亡率會隨著臭椿酮質(zhì)量濃度的增加而增大,0.10、1.00和10.00 μg/mL臭椿酮處理72 h時的細(xì)胞相對死亡率分別為6.01%、10.23%和21.36%。表明1.00 μg/mL臭椿酮在24 h內(nèi)就已經(jīng)造成細(xì)胞死亡,但是直至72 h細(xì)胞死亡的數(shù)量并沒有顯著增加,說明受到臭椿酮影響失去活力的細(xì)胞占根尖細(xì)胞的比例變化不大。細(xì)胞有絲分裂周期為G1、S、G2和M4個時期,在外部條件相同的情況下,處于一個時期的細(xì)胞數(shù)量幾乎不變,例如,處于M期的細(xì)胞占細(xì)胞總數(shù)5%~10%[24]。結(jié)合根尖分生區(qū)有絲分裂現(xiàn)象觀察,可以推測:臭椿酮的存在可以引起部分植物根尖分生區(qū)細(xì)胞死亡,使這一部分細(xì)胞無法進(jìn)入M期,或進(jìn)入M期后引起有絲分裂畸形,從而減少新生細(xì)胞數(shù)量,導(dǎo)致根無法正常生長。
3 結(jié)論
臭椿酮對多種植物的種子萌發(fā)和幼苗生長具有很強(qiáng)的抑制作用,且在單子葉和雙子葉植物之間沒有選擇性。臭椿酮對植物種子吸水能力無明顯影響。植物生長量的減少可能與臭椿酮抑制其細(xì)胞分裂有關(guān)。SOD、CAT和POD的酶活試驗(yàn)結(jié)果表明,臭椿酮提高了植物的抗氧化能力。DAB和NBT染色結(jié)果表明,臭椿酮誘導(dǎo)了植物組織中活性氧的積累,伊文思藍(lán)染色說明臭椿酮可以引起一部分細(xì)胞死亡,但并未隨著時間的推移而增長,推測臭椿酮的存在可以引起部分植物根尖分生區(qū)細(xì)胞死亡,并使這一部分細(xì)胞是無法進(jìn)入細(xì)胞有絲分裂的M期,或進(jìn)入M期后會引起有絲分裂畸形,從而減少新生細(xì)胞數(shù)量,導(dǎo)致植物根系無法正常生長。本研究結(jié)果表明,臭椿酮表現(xiàn)出較強(qiáng)抑制種子萌發(fā)和幼苗生長活性,究其原因可能為活性氧積累,或引起了部分植物根尖分生區(qū)細(xì)胞死亡使其無法正常有絲分裂。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
