α-法尼烯在巴斯德畢赤酵母中的生物合成
2021-08-31 07:49:34劉慧陳勝玲徐建中張偉國(guó)
食品與發(fā)酵工業(yè) 2021年16期
劉慧,陳勝玲,徐建中*,張偉國(guó)*
1(江南大學(xué) 生物工程學(xué)院,江蘇 無(wú)錫,214122)2(工業(yè)生物技術(shù)教育部重點(diǎn)實(shí)驗(yàn)室(江南大學(xué)),江蘇 無(wú)錫,214122)
倍半萜類(lèi)化合物在醫(yī)藥、化妝品、調(diào)味品和生物能源方面具有巨大的經(jīng)濟(jì)價(jià)值[1-2]。法尼烯是植物產(chǎn)生的最簡(jiǎn)單的無(wú)環(huán)倍半萜之一,在蘋(píng)果皮中分布豐富[3]。但是,它的天然合成受到植物生長(zhǎng)的嚴(yán)重限制,無(wú)法滿足市場(chǎng)需求[4]。因此,研究人員已經(jīng)努力設(shè)計(jì)微生物細(xì)胞工廠以生產(chǎn)法尼烯[5-9]。最近,已經(jīng)在大腸桿菌[5]、釀酒酵母[6]和解脂耶氏酵母[7-9]中通過(guò)設(shè)計(jì)代謝工程來(lái)生產(chǎn)法尼烯。與這些宿主相比,巴斯德畢赤酵母表達(dá)系統(tǒng)成本低、周期短、表達(dá)量高且可高密度培養(yǎng),具有大規(guī)模生產(chǎn)α-法尼烯的潛力。一些研究已經(jīng)證明,巴斯德畢赤酵母是生產(chǎn)類(lèi)異戊二烯的合適宿主[10]。然而,目前沒(méi)有報(bào)道描述巴斯德畢赤酵母中α-法尼烯的異源生產(chǎn)。
在本項(xiàng)研究中,首先成功構(gòu)建了產(chǎn)α-法尼烯的巴斯德畢赤酵母重組菌株。然后揭示了巴斯德畢赤酵母甲羥戊酸途徑和α-法尼烯合成途徑中限速步驟。之后對(duì)限速酶組合過(guò)表達(dá)并優(yōu)化基因拷貝數(shù)以平衡代謝途徑逐步提高α-法尼烯產(chǎn)量。最后通過(guò)外源添加不飽和脂肪酸促進(jìn)α-法尼烯分泌到細(xì)胞外,在搖瓶中最終獲得α-法尼烯產(chǎn)量約1.40 g/L[0.32 g/g 細(xì)胞干重(dry cell weight,DCW)]。這些策略提供了增強(qiáng)巴斯德畢赤酵母中α-法尼烯生產(chǎn)的有效方法,并為其他增值化學(xué)品的生物生產(chǎn)提供新的思路。
1 材料與方法
1.1 菌株與質(zhì)粒
釀酒酵母S288C、大腸桿菌DH5α、野生型巴斯德畢赤酵母X33、大腸桿菌-酵母穿梭質(zhì)粒PGAPZA,購(gòu)自Invitrogen。
1.2 試劑與儀器
限制性?xún)?nèi)切酶,Thermo Scientific;TaqDNA聚合酶、DNA Marker、DNA切膠回收試劑盒、T4DNA連接酶,TaKaRa;PCR引物、質(zhì)粒快速提取試劑盒,上海生工;酵母基因組提取試劑盒,康為世紀(jì);博來(lái)霉素、遺傳霉素,Invitrogen;標(biāo)準(zhǔn)品β-法尼烯、玻璃珠,Sigma;正十二烷,羅恩試劑;油酸、硬脂酸、棕櫚酸、亞麻酸、亞油酸、棕櫚油酸均為國(guó)藥分析純。
臺(tái)式高速冷凍離心機(jī),德國(guó)Sigma公司;真空冷凍干燥機(jī),日本Toshiba公司;高效氣相色譜儀,日本Shimadzu公司;電脈沖基因轉(zhuǎn)移儀,美國(guó)BIO-RAD公司。
1.3 培養(yǎng)基
LB培養(yǎng)基(g/L):蛋白胨10,酵母膏5,氯化鈉5;YPD培養(yǎng)基(g/L):葡萄糖20,蛋白胨20,酵母膏10;BMDY培養(yǎng)基(g/L):葡萄糖20,蛋白胨20,酵母膏10,YNB 13.4,生物素0.5,磷酸鉀緩沖液100 mmol/L (pH 6.0)。
1.4 重組質(zhì)粒以及菌株的構(gòu)建
表1和表2列出了該研究中使用的巴斯德畢赤酵母重組菌株和質(zhì)粒。利用GENEWIZ對(duì)來(lái)自蘋(píng)果(Malusdomestica)的α-法尼烯合成酶(α-farnesene synzyme,AFS)基因(Gene ID:103446592)進(jìn)行了密碼子優(yōu)化和合成。所有酶表達(dá)載體均基于PGAPZA載體插入了相應(yīng)表達(dá)基因。ERG10,ERG13,ERG12,ERG8,ERG19,IDI1,ERG20分別從巴斯德畢赤酵母X33基因組中擴(kuò)增相應(yīng)基因。tHmg1從釀酒酵母S288C中擴(kuò)增被截短的HMG1(Gene ID:854900)。AFSLERG20是AFS和ERG20的融合基因,兩個(gè)基因之間用GSG 3個(gè)氨基酸連接。重組質(zhì)粒通過(guò)使用AvrII酶切線性化然后將其整合到巴斯德畢赤酵母X33基因組上。
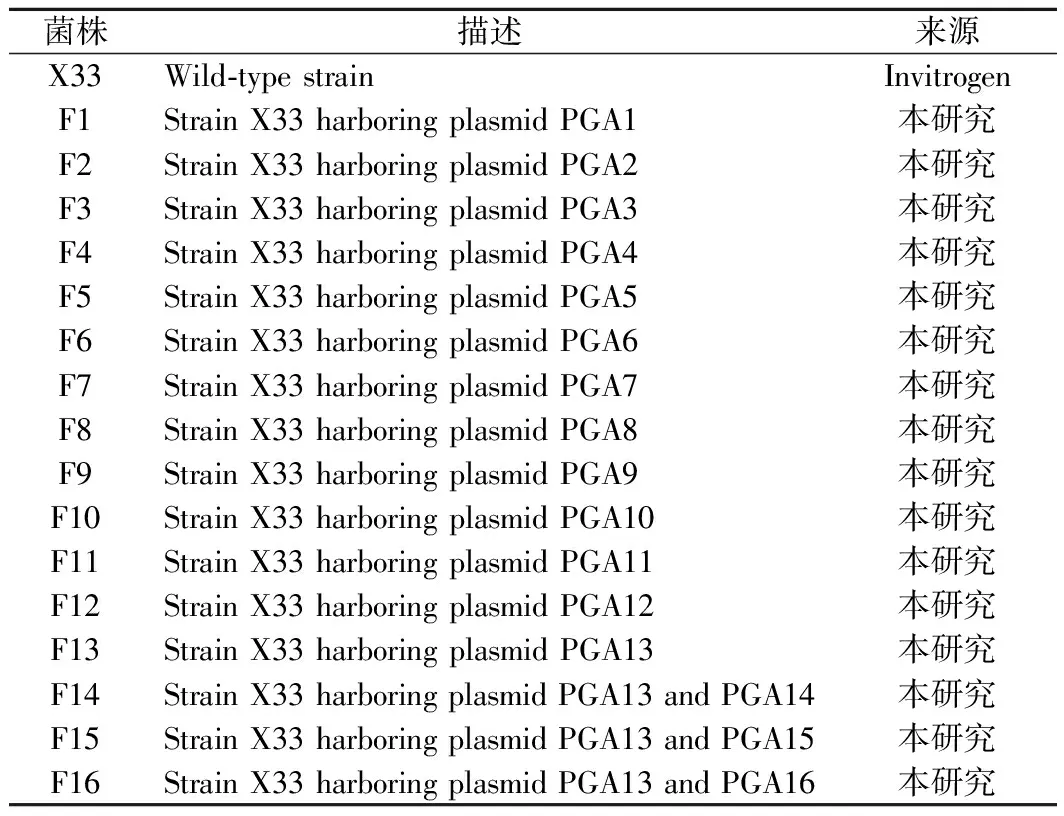
表1 本研究使用的菌株Table 1 The strains used in this study

表2 本研究中使用的質(zhì)粒Table 2 The plasmids used in this study
1.5 巴斯德畢赤酵母的轉(zhuǎn)化和篩選
按照J(rèn)OAN等[11]的方法進(jìn)行巴斯德畢赤酵母感受態(tài)細(xì)胞的制備和電穿孔。30 ℃、90 r/min復(fù)蘇1 h,涂在含有100 mg/L博來(lái)霉素或者400 mg/L遺傳霉素G418的YPD板上以篩選重組菌落。使用酵母基因組提取試劑盒提取酵母基因組,并通過(guò)PCR驗(yàn)證基因整合到基因組中。
1.6 細(xì)胞干重測(cè)定
取1 mL發(fā)酵液12 000 r/min離心棄上清液,用清水洗滌2次后于烘箱中90 ℃烘干至恒重,得到菌體干重。
1.7 α-法尼烯含量的測(cè)定
按照LIU等[9]的方法進(jìn)行α-法尼烯含量的測(cè)定。收集兩相培養(yǎng)物的上層有機(jī)相,并以12 000 r/min離心10 min以去除細(xì)胞碎片,然后使用日本Shimadzu GC-2010 Plus配備有火焰離子化檢測(cè)器的氣相色譜儀,以1∶20的分流比進(jìn)樣,并在db-17色譜柱(30.0 m×0.32 mm,0.25 μm)上分離出1 μL樣品。烤箱溫度最初在80 ℃下保持1 min,然后以15 ℃/min的速度升至245 ℃保持1.5 min,以相同的速度升溫至280 ℃后運(yùn)行時(shí)間為5 min。氦氣用作載氣,入口壓力為96 kPa。探測(cè)器溫度保持在250 ℃。通過(guò)比較樣品和β-法尼烯標(biāo)準(zhǔn)品的峰面積進(jìn)行定量分析。
1.8 重組畢赤酵母搖瓶培養(yǎng)
將重組菌接種于10 mL YPD培養(yǎng)基中,30 ℃、200 r/min 下培養(yǎng)17 h作為種子液。以10%接種量接種于50 mL BMDY培養(yǎng)基的500 mL搖瓶進(jìn)行搖瓶培養(yǎng)72 h。培養(yǎng)基中添加體積分?jǐn)?shù)為10%正十二烷來(lái)收集α-法尼烯。不飽和脂肪酸包括油酸、亞油酸、亞麻酸、棕櫚油酸(均為液態(tài)),直接加入培養(yǎng)基中。飽和脂肪酸包括硬脂酸和棕櫚酸(均為固體顆粒),為保證其在培養(yǎng)基中能夠均勻地分散,將其溶于200 μL無(wú)水乙醇后進(jìn)行添加,因此對(duì)于飽和脂肪酸,對(duì)照組應(yīng)加入200 μL的乙醇。每組試驗(yàn)設(shè)置3個(gè)平行,分別測(cè)定α-法尼烯產(chǎn)量和菌體干重。
2 結(jié)果與分析
2.1 巴斯德畢赤酵母中α-法尼烯生物合成途徑的構(gòu)建
通過(guò)GENEWIZ對(duì)來(lái)源于蘋(píng)果的法尼烯合成酶基因(AFS)進(jìn)行了密碼子優(yōu)化和合成。構(gòu)建了組成型表達(dá)質(zhì)粒PGAPZA-AFS,通過(guò)AvrII將其線性化并整合到巴斯德畢赤酵母的基因組中從而獲得重組菌株F1。僅使用少量線性化質(zhì)粒(500 ng/80 mL 細(xì)胞)來(lái)確保單拷貝整合的通用性[12]。將重組菌株在BMDY培養(yǎng)基中培養(yǎng)72 h,其中分別覆蓋體積分?jǐn)?shù)為10%的十二烷以收獲產(chǎn)物。GC/GC-MS分析表明(圖1),在菌株F1中觀察到法尼烯產(chǎn)量為(451.13±18.92) mg/L。先前的研究表明,來(lái)源于蘋(píng)果的α-法尼烯合成酶在大腸桿菌中α-法尼烯的產(chǎn)量為1.56 mg/L[3],在釀酒酵母中為4 mg/L[20],在解脂耶氏酵母中為0.13 g/L[9]。研究結(jié)果表明,重組畢赤酵母中α-法尼烯的產(chǎn)量是遠(yuǎn)高于其他宿主。這可能是與巴斯德畢赤酵母外源蛋白表達(dá)水平高,高密度培養(yǎng)特性有關(guān)[21]。此外,菌株之間的生產(chǎn)能力差異很大也突出了巴斯德畢赤酵母合成α-法尼烯的優(yōu)越性。
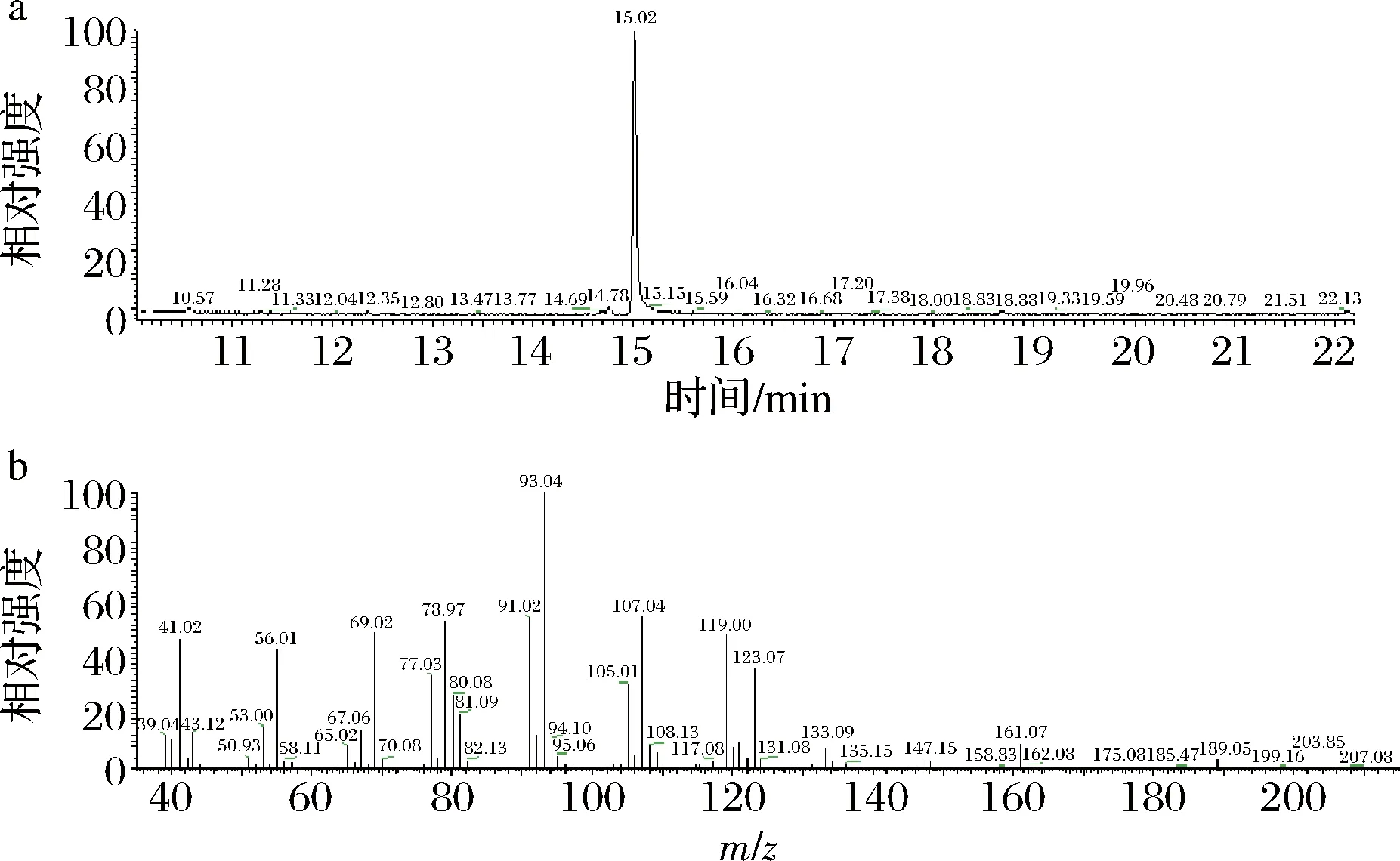
a-氣相色譜圖;b-質(zhì)譜圖圖1 重組菌株F1合成的α-法尼烯氣相色譜以及質(zhì)譜圖Fig.1 GC/GC-MS of α-farnesene synthesized by recombinant strain F1
2.2 揭示甲羥戊酸途徑和α-法尼烯合成途徑中的限速步驟
截短的HMG1(tHmg1)基因可以有效解除膽固醇合成途徑中麥角固醇等的反饋抑制[13],過(guò)表達(dá)tHmg1通常認(rèn)為是有益于類(lèi)異戊二烯在酵母中生產(chǎn)[14]。為了改善α-法尼烯在畢赤酵母中的生物合成,在菌株F1基礎(chǔ)上成功地過(guò)表達(dá)tHmg1得到菌株F2。菌株F2顯示α-法尼烯的產(chǎn)量達(dá)到(0.53±0.01) g/L,比F1中觀察到的產(chǎn)物水平提高約18%。
研究表明,tHmg1與甲羥戊酸(mevalonate ,MVA)途徑以及α-法尼烯合成途徑其他基因的共過(guò)量表達(dá)可能會(huì)使α-法尼烯的產(chǎn)量進(jìn)一步提高[15]。在此,將tHmg1分別與MVA途徑與α-法尼烯途徑(圖2)中涉及到的8個(gè)基因ERG10、ERG13、ERG12、ERG8、ERG19、IDI1、ERG20、AFSLERG20進(jìn)行組合過(guò)表達(dá)(圖3),并探究對(duì)α-法尼烯合成的影響,從而確定了限速步驟。所有的基因都整合到畢赤酵母的基因組中,并以強(qiáng)組成型啟動(dòng)子PGAP表達(dá)。與菌株F2相比,菌株F7、F8、F10、F11的α-法尼烯濃度增加了約37.13%~55.24%(圖3)。在F10/F11中,tHmg1和AFSLERG20組合過(guò)表達(dá)顯示最高的α-法尼烯產(chǎn)量為(0.70±0.03) g/L,其次是tHmg1和IDI1/ERG19組合過(guò)表達(dá)。然而,F(xiàn)11是在F10中額外過(guò)表達(dá)AFS,幾乎沒(méi)有增加α-法尼烯的產(chǎn)生,說(shuō)明此時(shí)AFS可能不是α-法尼烯合成途徑中的限速基因。同時(shí)tHmg1與ERG8/ERG10/ERG12/ERG13/ERG20基因的過(guò)表達(dá)導(dǎo)致α-法尼烯產(chǎn)量?jī)H產(chǎn)生細(xì)微的變化(圖3),可能是由于過(guò)表達(dá)導(dǎo)致中間體的積累和途徑不平衡[7,16]。結(jié)果表明,tHmg1,IDI1,ERG19,AFSLERG20是MVA途徑和α-法尼烯合成途徑的限速酶基因,過(guò)表達(dá)后可改善巴斯德畢赤酵母中α-法尼烯的合成。
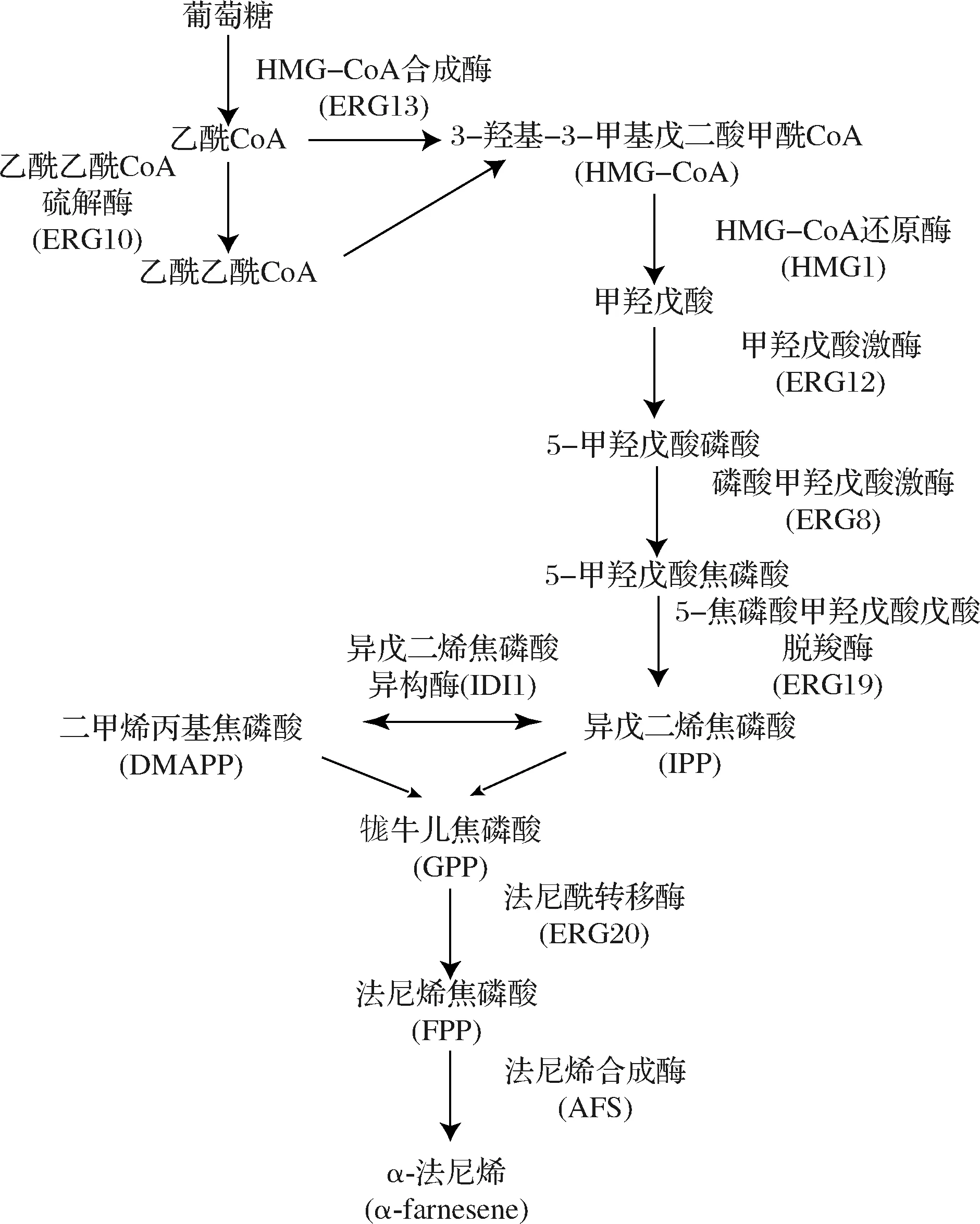
圖2 α-法尼烯的生物合成途徑Fig.2 The biosynthetic pathway of α-farnesene注:二甲基烯丙基焦磷酸(dimethylallyl pyrophosphate,DMAPP),異戊二烯焦磷酸(isopentenyl pyrophosphate,IPP),牻牛兒焦磷酸(geranyl pyrophosphate,GPP),法尼基焦磷酸(farnesyl pyrophosphate,F(xiàn)PP)
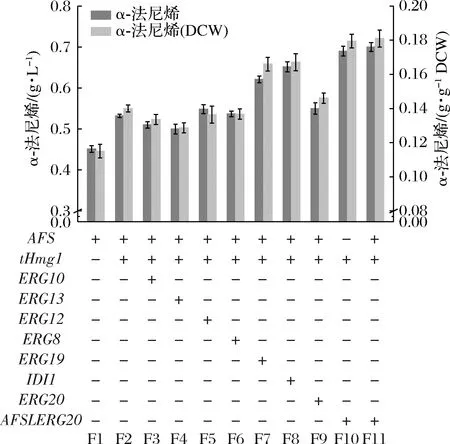
圖3 tHmg1與MVA途徑和α-法尼烯合成途徑基因分別組合過(guò)表達(dá)對(duì)巴斯德畢赤酵母中α-法尼烯產(chǎn)生的影響Fig.3 Effect of overexpression of tHmg1 in combination with genes of the mevalonate pathway and α-farnesene synthesis pathway on the production of α-farnesene in P.pastoris注:“+”代表包含對(duì)應(yīng)的基因;“-”代表不包含對(duì)應(yīng)的基因(下同)
2.3 過(guò)表達(dá)甲羥戊酸途徑以及α-法尼烯合成途徑限速酶促進(jìn)α-法尼烯的合成
結(jié)果表明,tHmg1與AFSLERG20組合過(guò)表達(dá)對(duì)系統(tǒng)的貢獻(xiàn)最大,其次是tHmg1與IDI1/ERG19。基于這些結(jié)果,選擇了tHmg1,IDI1,ERG19和AFSLERG20這4個(gè)關(guān)鍵酶基因,通過(guò)3個(gè)或者3個(gè)以上組合過(guò)表達(dá)嘗試進(jìn)一步提高菌株中α-法尼烯的產(chǎn)量。在菌株F12中,tHmg1,IDI1和AFSLERG20的組合過(guò)表達(dá),α-法尼烯的產(chǎn)量與菌株F10相比分別提高約25.96%(圖4)。有趣的是,菌株F13中tHmg1,ERG19和AFSLERG20組合過(guò)表達(dá)與對(duì)照菌株F10相比,α-法尼烯產(chǎn)量反而降低約18.57%(圖4)。菌株F13是在F10的基礎(chǔ)上過(guò)表達(dá)ERG19,推測(cè)可能是由于F13具有更高水平的ERG19酶的表達(dá)以造成有毒中間體IPP的積累,從而導(dǎo)致α-法尼烯產(chǎn)量降低[5]。
異戊烯基二磷酸異構(gòu)酶(由IDI1基因編碼)催化IPP向DMAPP的異構(gòu)化,并在GPP和FPP通量的分布中起關(guān)鍵作用[15]。為了驗(yàn)證猜想,通過(guò)在菌株F13引入1~2個(gè)拷貝IDI1基因,分別產(chǎn)生菌株F14和F15。菌株F14和F15的α-法尼烯產(chǎn)量達(dá)到約1.00 g/L,這暗示著平衡IPP和DMAPP池可能在增強(qiáng)倍半萜生物合成的碳通量中起重要作用。除此之外,構(gòu)建了菌株F16,在F15基礎(chǔ)上額外過(guò)表達(dá)1個(gè)拷貝AFSLERG20,α-法尼烯產(chǎn)量進(jìn)一步提高,如圖4所示,達(dá)到(1.09±0.02) g/L(菌株F1的2.42倍)。上述一系列組合的結(jié)果表明,優(yōu)化MVA途徑限速酶基因的組合表達(dá)可以有效地促進(jìn)畢赤酵母中倍半萜的異源生物合成。同時(shí)也揭示了在微生物宿主中建立高效的萜類(lèi)化合物途徑,必須小心平衡途徑中上下游代謝通量以避免有毒中間代謝物的積累。
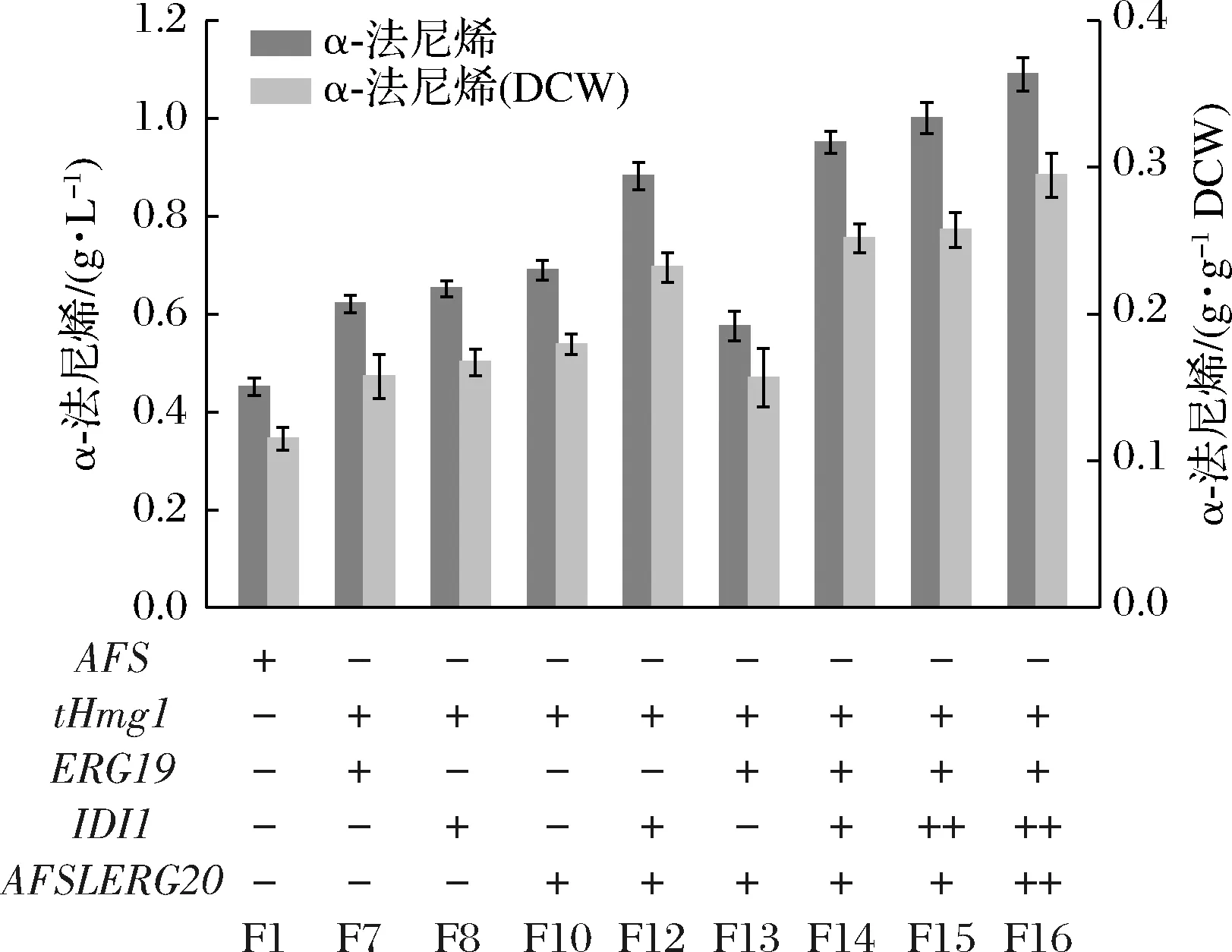
圖4 限速酶基因組合過(guò)表達(dá)以及基因拷貝數(shù)優(yōu)化對(duì)巴斯德畢赤酵母中α-法尼烯產(chǎn)生的影響Fig.4 Effects of rate-limiting enzyme gene combination overexpression and gene copy number optimization on the production of α-farnesene in P.pastoris
2.4 添加不飽和脂肪酸促進(jìn)α-法尼烯的分泌
小分子的萜類(lèi)化合物,例如倍半萜和類(lèi)黃酮,通常被分泌到細(xì)胞外[6]。然而,有限的細(xì)胞膜通透性是倍半萜分泌的障礙[6]。各項(xiàng)研究表明,增大不飽和脂肪酸(unsaturated fatty acid,UFA)/飽和脂肪酸(saturated fatty acid,SFA)的比例有助于增加有毒物質(zhì)跨膜的膜通透性[6, 17-19]。同時(shí),酵母細(xì)胞可以很容易地從培養(yǎng)基中吸收外源UFA和SFA,并迅速整合到膜脂質(zhì)中[19]。因此,通過(guò)向菌株F16培養(yǎng)基中外源添加脂肪酸,來(lái)試圖提高巴斯德畢赤酵母中α-法尼烯的產(chǎn)量。最初添加了10 mmol/L不飽和脂肪酸油酸(C18∶1)、亞油酸(C18∶2)、亞麻酸(C18∶3)、棕櫚油酸(C16∶1)、飽和脂肪酸硬脂酸(C18∶0)和棕櫚酸(C16∶0)于菌株F16的培養(yǎng)基中,首先確定了不同脂肪酸對(duì)重組菌株的α-法尼烯產(chǎn)量以及對(duì)細(xì)胞生長(zhǎng)的影響。如圖5所示,除了亞油酸,其他不飽和脂肪酸對(duì)α-法尼烯產(chǎn)量的提高均有積極影響,其中油酸的積極作用尤為顯著。之前的研究[19]和本研究都觀察到添加飽和脂肪酸對(duì)異戊二烯產(chǎn)量的增加并沒(méi)有積極影響。脂肪酸的添加可以促進(jìn)菌體的生長(zhǎng),這種促進(jìn)作用僅存在于3種不飽和脂肪酸中,飽和脂肪酸對(duì)菌體生長(zhǎng)均沒(méi)有促進(jìn)作用(圖5)。此外,亞油酸雖然對(duì)菌體生長(zhǎng)有積極作用,然而對(duì)MVA途徑似乎沒(méi)有促進(jìn)作用,這可能歸因于菌體對(duì)脂肪酸的利用有偏好性和菌株對(duì)不飽和脂肪酸氧化壓力的耐受能力不同[19]。因此選擇油酸來(lái)促進(jìn)α-法尼烯的分泌。為了最大程度地提高畢赤酵母中α-法尼烯的產(chǎn)量,優(yōu)化了油酸添加濃度。如圖6所示,確定向細(xì)胞提供20 mmol/L的油酸獲得最佳的α-法尼烯產(chǎn)量約為1.40 g/L,收率約為0.07 g/g,生產(chǎn)率約為0.019 g/(L·h)。α-法尼烯產(chǎn)量是出發(fā)菌株F1的3.1倍。
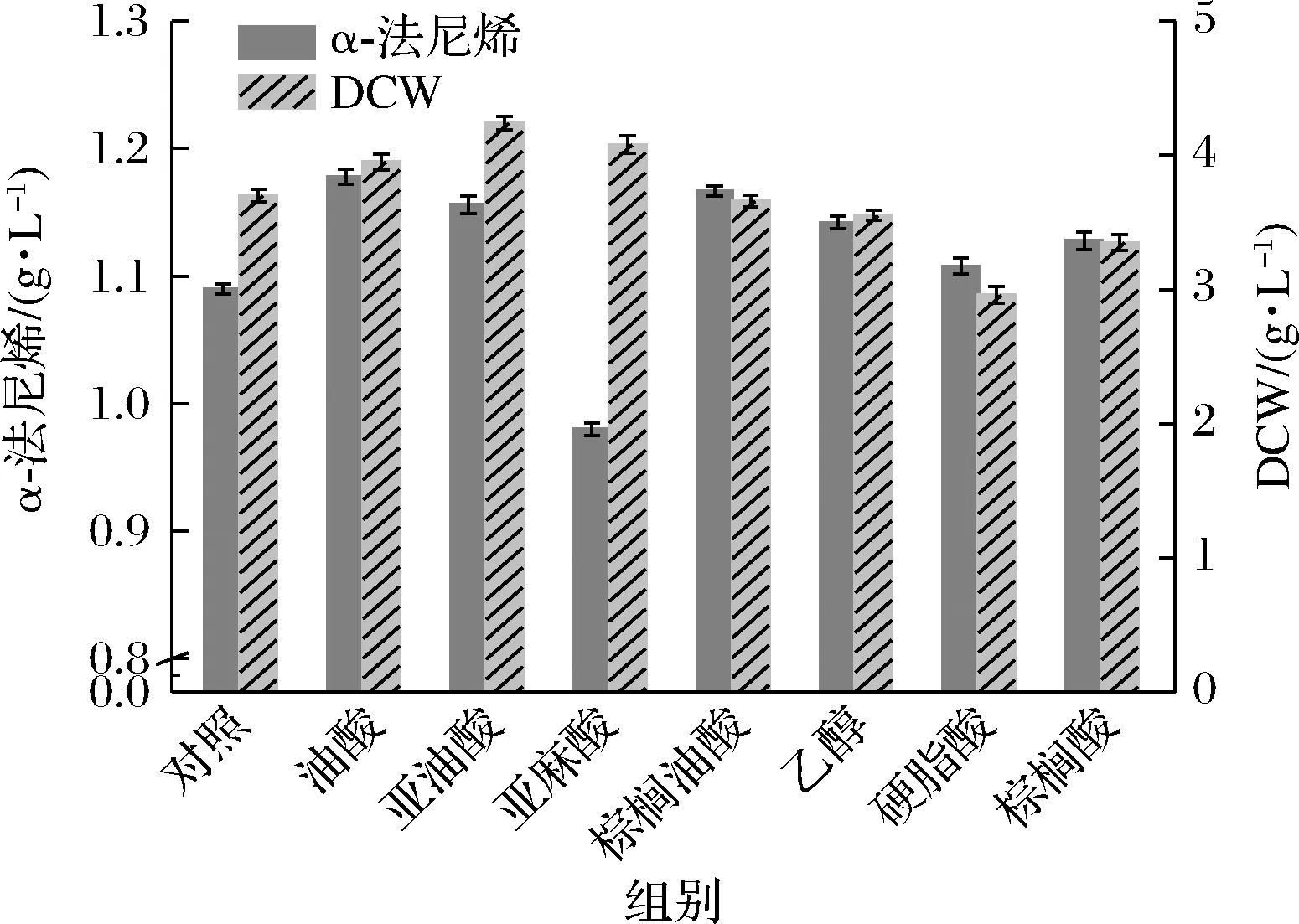
圖5 外源添加不同脂肪酸對(duì)巴斯德畢赤酵母中α-法尼烯合成的影響Fig.5 The effect of different fatty acids on the synthesis of α-farnesene in P.pastoris
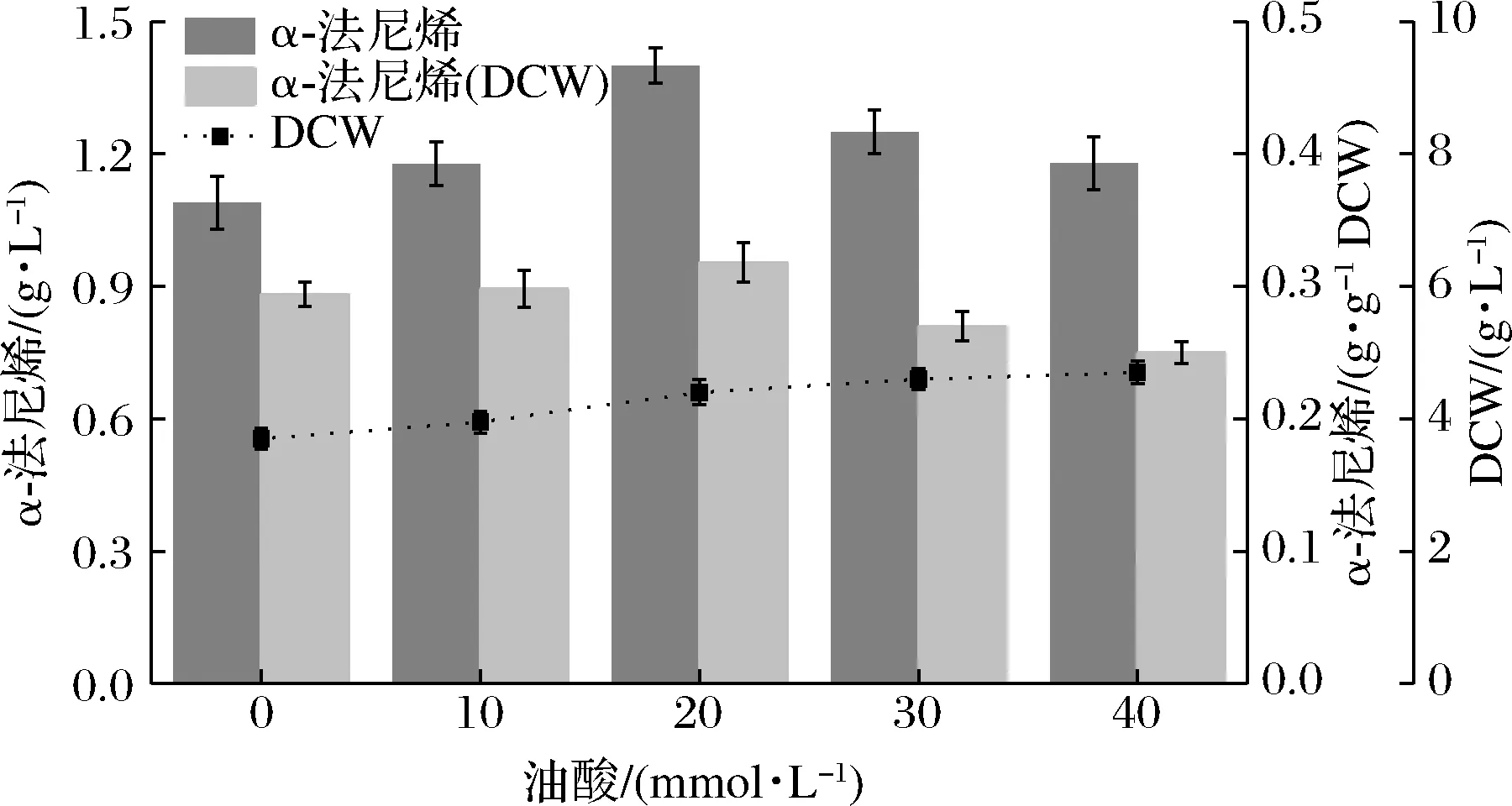
圖6 外源添加油酸的最適濃度Fig.6 Optimal concentration of oleic acid
3 結(jié)論與討論
本研究是首次以發(fā)酵周期短,易高密度培養(yǎng)特性的巴斯德畢赤酵母作為底盤(pán)微生物細(xì)胞來(lái)生產(chǎn)α-法尼烯,揭示了MVA途徑和α-法尼烯合成途徑中限速步驟,對(duì)限速酶組合過(guò)表達(dá)并優(yōu)化基因拷貝數(shù)以平衡代謝途徑逐步提高α-法尼烯產(chǎn)量,最后通過(guò)外源添加不飽和脂肪酸促進(jìn)α-法尼烯分泌到細(xì)胞外,在搖瓶中菌株F16最終獲得α-法尼烯產(chǎn)量約1.40 g/L(0.32 g/g DCW),生產(chǎn)率約為0.019 g/(L·h)。在搖瓶水平上,菌株F16的α-法尼烯生產(chǎn)率是解脂耶氏酵母[9]的1.38倍,大腸桿菌[3]的2.4倍,釀酒酵母[22]的3.8倍。盡管巴斯德畢赤酵母工程菌中α-法尼烯產(chǎn)量顯著提高,但是競(jìng)爭(zhēng)途徑[13]以及代謝串?dāng)_[23]仍然是阻止目標(biāo)化合物有效合成的障礙。因此,減弱競(jìng)爭(zhēng)途徑和代謝串?dāng)_將是未來(lái)努力的方向。總之,以巴斯德畢赤酵母作為底盤(pán)微生物生產(chǎn)α-法尼烯是非常有前景的,為大規(guī)模發(fā)酵生產(chǎn)α-法尼烯提供新的希望。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年10期)2019-11-25 07:33:58
中國(guó)果業(yè)信息(2019年10期)2019-11-13 01:21:34
中學(xué)生數(shù)理化·高一版(2018年9期)2018-10-09 06:46:50
湖南教育·C版(2018年3期)2018-06-05 16:54:36
聚氯乙烯(2018年9期)2018-02-18 01:11:34
中國(guó)衛(wèi)生(2016年3期)2016-11-12 13:23:26
