竹葉提取物和敲除轉(zhuǎn)錄因子DAL80對釀酒酵母精氨酸代謝的調(diào)控互作影響
2021-08-31 02:34:30劉小杰婁行行陳啟和
食品科學(xué) 2021年16期
劉小杰,婁行行,陳啟和,*
(1.上海城建職業(yè)學(xué)院,上海 201415;2.浙江大學(xué)生物系統(tǒng)工程與食品科學(xué)學(xué)院,浙江 杭州 310058)
黃酒為中國著名的發(fā)酵酒,含有氨基酸、多肽、低聚糖、多糖以及酚類化合物[1-2],但黃酒在發(fā)酵過程中會產(chǎn)生風(fēng)險化合物,如氨基甲酸乙酯(ethyl carbamate,EC),其對人體具有潛在的遺傳毒性和致癌性[3-4],黃酒中EC的含量明顯高于其他酒類[5],因此降低黃酒中EC含量對黃酒產(chǎn)業(yè)的可持續(xù)發(fā)展具有重要意義。EC通常是由乙醇與尿素自發(fā)反應(yīng)形成的,因此通過控制尿素以減少黃酒中EC是控制其含量的主要思路之一[6]。黃酒中的尿素一部分來源于食品原料,另一部分來源于釀酒酵母對精氨酸(arginine,Arg)的降解作用[7-8]。多年來,許多研究者均致力于探究發(fā)酵食品中EC的控制方法,目前主要從優(yōu)化生產(chǎn)工藝[9-10]、外源添加脲酶分解尿素[11-12]、基因工程手段修飾發(fā)酵菌株[13-15]、外源添加天然產(chǎn)物調(diào)控Arg降解[16]等方面展開研究。
在釀酒酵母中氮代謝阻遏效應(yīng)相關(guān)基因的轉(zhuǎn)錄水平受4 個GATA家族轉(zhuǎn)錄因子GLN3、GZF3、GAT1、DAL80和1 個全局調(diào)控蛋白Ure2p的調(diào)控。其中,DAL80不僅能調(diào)控氮代謝阻遏效應(yīng)相關(guān)基因的表達(dá),也能與其他因子調(diào)控彼此的表達(dá)[17]。焦志華[18]的研究表明敲除釀酒酵母BY4741中的DAL80可以降低黃酒中38.5%的EC含量。竹葉提取物(bamboo leaves extract,BLE)作為食品添加劑,具有抗氧化、抑制有害物質(zhì)的生成等作用[19-20]。BLE加入黃酒會抑制EC的形成,其具有多種作用機(jī)制[16,21-22]。目前,控制黃酒釀造EC的方法集中于使用一種手段,較少將2 種及以上的方法聯(lián)合使用。因此,本實驗添加BLE,并結(jié)合GATA家族轉(zhuǎn)錄因子中氮阻遏代謝轉(zhuǎn)錄調(diào)控因子DAL80的敲除手段,闡明二者對釀酒酵母細(xì)胞Arg代謝的互作調(diào)控機(jī)制。
1 材料與方法
1.1 材料與試劑
釀酒酵母BY4741(MATa、his3Δ1、leu2Δ0、met15Δ、ura3Δ0),釀酒酵母DAL80敲除菌(DAL80Δ),實驗室-80 ℃保藏。
BLE 浙江圣氏生物科技有限公司;蛋白胨、酵母提取物、L-精氨酸鹽酸鹽、L-亮氨酸、酵母氮源(yeast nitrogen base,YNB)(含硫酸銨) 生工生物工程(上海)股份有限公司;L-組氨酸、L-甲硫氨酸、尿嘧啶阿拉丁試劑(上海)有限公司。
總蛋白濃度測定試劑盒 生工生物工程(上海)股份有限公司;精氨酸酶活力測定試劑盒 美國博世生物技術(shù)有限公司;酵母RNA提取試劑盒 美國Omega公司;PrimeScriptTMRT reagent Kit with gDNA Eraser、TB GreenTMPremix ExTaqTMII 日本Takara生物工程株式會社。
酵母膏胨葡萄糖瓊脂(yeast extract peptone dextrose,YPD)培養(yǎng)基:酵母提取物、蛋白胨、葡萄糖質(zhì)量分?jǐn)?shù)分別為1%、2%、2%;Arg培養(yǎng)基:質(zhì)量分?jǐn)?shù)1.7%YNB(含硫酸銨)、質(zhì)量分?jǐn)?shù)2%葡萄糖、20 mmol/LL-精氨酸鹽酸鹽、10×氨基酸混合物(包含200 mg/LL-組氨酸、1 000 mg/LL-亮氨酸、200 mg/LL-甲硫氨酸、200 mg/L尿嘧啶);配制不同質(zhì)量濃度的BLE(0、150、300、500 mg/L)標(biāo)準(zhǔn)溶液,過濾除菌,4 ℃保存。
1.2 儀器與設(shè)備
1510酶標(biāo)儀 美國Thermo Fisher公司;ZQLY-300振蕩培養(yǎng)箱 上海知楚儀器有限公司;JY92-IIDN超聲波細(xì)胞破碎儀 寧波新芝生物科技股份有限公司;JP-040S超聲清洗儀 深圳市潔盟清洗設(shè)備有限公司;Agilent 1200色譜儀、VORTEX-5渦旋儀 廣州永程設(shè)備有限公司;DD-5M大型離心機(jī) 湖南湘儀實驗室儀器開發(fā)有限公司;5417R桌面離心機(jī) 德國艾本德股份公司;SIM-100超微量可見分光光度計 杭州新景生物試劑開發(fā)有限公司。
1.3 方法
1.3.1 菌株生長性能評價
通過生長曲線、濕質(zhì)量測定、點板實驗和細(xì)胞活力測定,綜合評價BY4741和DAL80敲除菌在BLE存在時的生長規(guī)律,具體操作參見張偉平等[17]研究。
1.3.2 發(fā)酵液和胞內(nèi)尿素含量的測定
Arg培養(yǎng)基中培養(yǎng)48 h的釀酒酵母BY4741和DAL80敲除菌培養(yǎng)液于室溫、4 000 r/min離心10 min,取上清液檢測發(fā)酵液尿素含量。收集細(xì)胞用去離子水重懸,再以相同條件離心后重懸。最后棄上清液,加入10 mL去離子水重懸細(xì)胞,將離心管置于冰上,用超聲波細(xì)胞破碎儀破碎細(xì)胞10 min,離心得上清液檢測胞內(nèi)尿素。將試劑I(硫酸120 mL、磷酸50 mL、FeCl30.05 g、蒸餾水330 mL)和試劑II(0.5 mg/mL的二乙酰一肟、0.1 mg/mL的硫銨脲)按體積比2∶1混合,制成顯色劑。將2.5 mL顯色劑加入0.5 mL樣品,沸水浴反應(yīng)15 min后迅速冷卻至室溫,在526 nm波長處檢測吸光度變化[23]。
1.3.3 Arg含量的測定
采用高效液相色譜法在線衍生法測定發(fā)酵液或破碎后細(xì)胞液中的Arg含量[24]。
1.3.4 精氨酸酶活力的測定
胞內(nèi)蛋白提取:收集培養(yǎng)的酵母細(xì)胞2 mL,于4 ℃、4 000 r/min離心10 min,棄上清液,用磷酸鹽緩沖液重懸,相同條件離心2 次并棄上清液,用400 μL的Tris-HCl緩沖液(pH 7.4)重懸細(xì)胞,加入4 μL 10 mmol/L苯甲基磺酰氟(phenylmethanesulfonyl fluoride,PMSF)溶液和4 μL的150 mmol/L二硫蘇糖醇(dithiothreitol,DTT)溶液,超聲破碎細(xì)胞15 min,吸取上清液,用總蛋白濃度試劑盒測定總蛋白濃度。用精氨酸酶活力試劑盒測定精氨酸酶活力。1 個單位酶活力(U)定義為:pH 7.4、30 ℃時,每分鐘將1 μmol/L Arg轉(zhuǎn)化為尿素和鳥氨酸所需的酶量。
1.3.5 氮代謝相關(guān)基因轉(zhuǎn)錄水平的實時聚合酶鏈?zhǔn)椒磻?yīng)(real-time polymerase chain reaction,real-time PCR)檢測
將菌株培養(yǎng)48 h后,用酵母RNA提取試劑盒提取酵母總RNA,用超微量可見分光光度計測定提取RNA的濃度及質(zhì)量,基因組DNA去除反應(yīng)體系如表1所示,42 ℃保溫2 min后置于冰上。逆轉(zhuǎn)錄PCR體系(20 μL)如表2所示,37 ℃保溫15 min,85 ℃熱激5 s后保存于4 ℃。長期保存于-20 ℃或更低溫度。引物由杭州有康生物科技有限公司設(shè)計并合成,引物序列見表3,保存于-20 ℃。real-time PCR體系(20 μL)如表4所示。real-time PCR程序如表5所示,以ACT1基因作為內(nèi)標(biāo)基因,用real-time PCR熒光定量數(shù)據(jù)分析(2-ΔΔCt法),對擴(kuò)增基因進(jìn)行相對定量分析。

表1 基因組DNA去除反應(yīng)體系Table 1 Reaction system used for genomic DNA removal

表2 逆轉(zhuǎn)錄反應(yīng)體系Table 2 Reverse transcription reaction system

表3 引物序列Table 3 Primer sequences used in this study

表4real-time PCR體系Table 4 Real-time PCR reaction system

表5real-time PCR程序Table 5 Real-time PCR reaction procedures
2 結(jié)果與分析
2.1 BLE對釀酒酵母BY4741細(xì)胞生長及代謝的影響
2.1.1 BLE對BY4741生長的影響
通過生長曲線檢測、濕質(zhì)量測定、點板實驗和細(xì)胞活力測定,對BY4741在不含(對照組)和含不同質(zhì)量濃度BLE的Arg培養(yǎng)基中生長性能進(jìn)行評價。如圖1A、B所示,BY4741在不同質(zhì)量濃度BLE處理下的生長曲線差異不大,發(fā)酵48 h后,各組間的濕質(zhì)量差異也不大,僅BLE 500 mg/L質(zhì)量濃度組略有降低。點板實驗表明,各組間菌落形態(tài)及數(shù)量差異均不大(圖1C)。細(xì)胞活力實驗(圖1D)結(jié)果表明,培養(yǎng)24 h后各組間細(xì)胞活力無顯著差異,其中BLE 500 mg/L質(zhì)量濃度組略有降低,故BLE對BY4741的生長無顯著影響。通過如圖1所示的4 個生長性能評價實驗的結(jié)果可知,在質(zhì)量濃度0~500 mg/L范圍內(nèi),BLE對BY4741菌的生長影響不大,這意味著BLE添加可以進(jìn)行后續(xù)氮代謝水平相關(guān)研究,也為BLE在實際發(fā)酵過程的應(yīng)用提供前提條件。
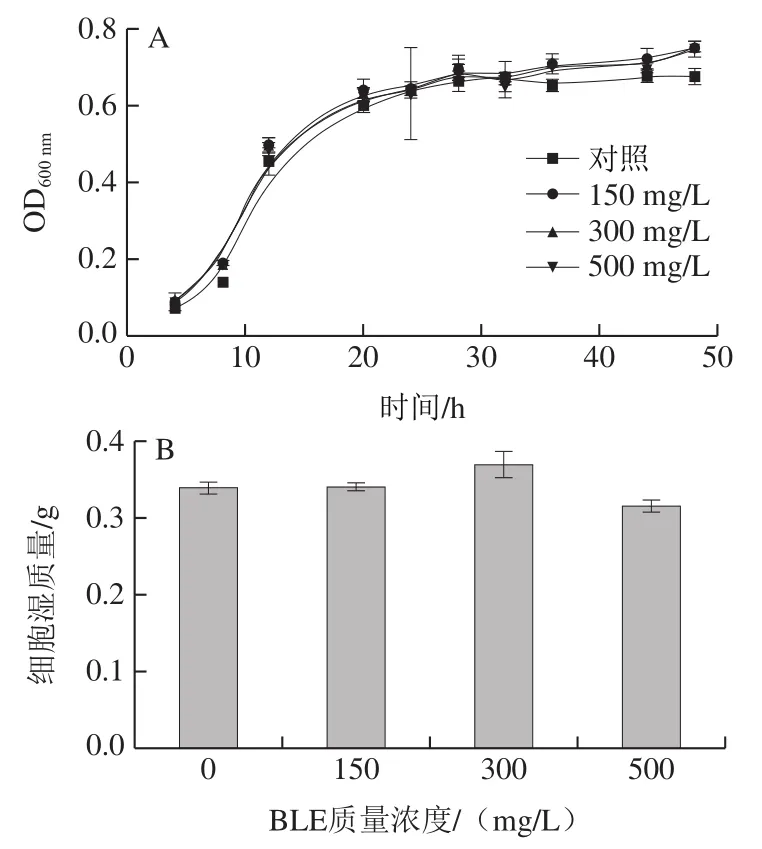

圖1 BLE質(zhì)量濃度對含有Arg培養(yǎng)體系中的BY4741生長的影響Fig. 1 Effects of different BLE concentrations on the growth of BY4741 in culture medium containing Arg
2.1.2 BLE對BY4741細(xì)胞氮代謝的影響
尿素和Arg是釀酒酵母氮代謝過程的重要物質(zhì),其中尿素可以與乙醇自發(fā)反應(yīng)生成EC,而Arg在精氨酸酶的作用下,可以降解產(chǎn)生尿素。因此,為探究BLE在黃酒模擬發(fā)酵體系中可能的氮代謝調(diào)控作用,利用Arg培養(yǎng)基培養(yǎng)BY4741酵母,通過比較BY4741發(fā)酵過程中尿素和Arg代謝水平,探究BLE對BY4741細(xì)胞氮代謝的調(diào)控作用,結(jié)果如圖2所示。

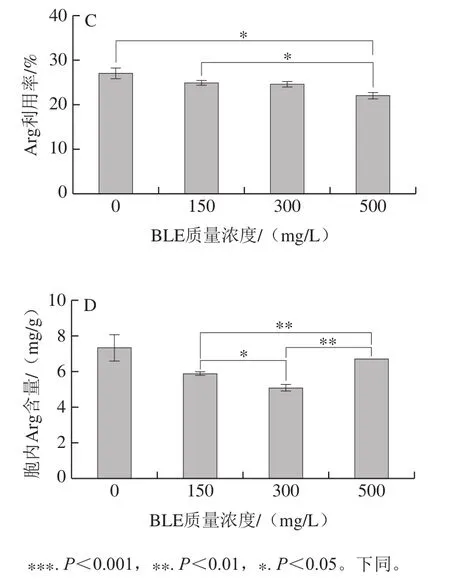
圖2 BY4741在不同BLE質(zhì)量濃度結(jié)束發(fā)酵時發(fā)酵液尿素含量(A)、胞內(nèi)尿素含量(B)、發(fā)酵液Arg利用率(C)、胞內(nèi)Arg含量(D)Fig. 2 Effects of different BLE concentrations on urea concentration in fermentation broth (A), intracellular urea concentration (B), Arg utilization rate in fermentation broth (C), and intracellular Arg concentration (D) of BY4741
添加BLE會增加BY4741發(fā)酵液中尿素積累,且添加質(zhì)量濃度越高,效果越明顯;當(dāng)添加質(zhì)量濃度為500 mg/L時,發(fā)酵液中尿素積累明顯增多。胞內(nèi)尿素處理組的積累較對照組更低,但隨著BLE質(zhì)量濃度的增加,BLE對尿素積累的影響減弱。胞外的尿素質(zhì)量濃度越高,胞內(nèi)的尿素含量隨之降低,可能是BLE中的某些物質(zhì)影響尿素的轉(zhuǎn)運或者尿素的重吸收,也可能是BLE影響尿素代謝酶活力,如尿素羧化酶和脲基甲酸鹽水解酶活力[25]。BLE還降低了胞內(nèi)Arg的積累,但與對照組相比,BLE對其影響不顯著。值得注意的是,添加BLE降低了BY4741細(xì)胞對Arg的利用率,說明BLE可能抑制Arg的分解代謝,添加質(zhì)量濃度為500 mg/L的影響最為突出。這說明添加BLE后,發(fā)酵液中積累的尿素并不能僅歸因于培養(yǎng)基中Arg的降解,還可能是BLE影響尿素的進(jìn)一步代謝,其他物質(zhì)合成尿素等因素導(dǎo)致尿素的積累,但這需要深入研究。因為BLE能降低Arg的利用率,且影響程度與BLE質(zhì)量濃度呈正相關(guān),所以選擇500 mg/L BLE作為后續(xù)研究的添加質(zhì)量濃度。
2.2 BLE與轉(zhuǎn)錄因子DAL80對釀酒酵母氮代謝途徑的影響
2.2.1 BLE對DAL80敲除菌株和野生型BY4741生長的影響
通過生長曲線、濕質(zhì)量測定、點板實驗和細(xì)胞活力測定實驗評價BLE對DAL80敲除菌和野生型BY4741的生長影響,BLE添加質(zhì)量濃度為500 mg/L,結(jié)果如圖3所示。首先CK組2 株菌的生長曲線差異不大,僅在20~25 hDAL80敲除菌株的OD600nm比野生型BY4741略高;質(zhì)量濃度500 mg/L BLE組中2 株菌的生長曲線也差異不大,幾乎重合。對照組和質(zhì)量濃度500 mg/L BLE組的生長曲線差異不大(圖3A)。在發(fā)酵48 h后,所有菌株的對照組細(xì)胞濕質(zhì)量略高于500 mg/L BLE組;而在添加相同劑量BLE時,敲除DAL80后細(xì)胞濕質(zhì)量有所提高(圖3B)。從圖3C可知,2 組間菌落形態(tài)及數(shù)量差異均不大。細(xì)胞活力實驗顯示培養(yǎng)24 h后,500 mg/L BLE組細(xì)胞的活力高于對照組(圖3D)。上述實驗說明,敲除DAL80和添加500 mg/L BLE對酵母菌的生長幾乎沒有影響。

圖3 500 mg/L BLE對DAL80敲除菌和BY4741生長的影響Fig. 3 Effects of 500 mg/L BLE on cell growth of BY4741 and DAL80Δ
2.2.2 BLE對DAL80敲除菌和野生型BY4741的Arg及尿素代謝水平的影響
如圖4所示,相較于野生型酵母菌株BY4741,敲除DAL80提高釀酒酵母胞外尿素含量、Arg利用率及精氨酸酶活力,分別提高54.8%、27.3%、71.1%。BLE處理后,DAL80敲除提高精氨酸酶活力,BLE的干預(yù)使得提高幅度降低4.2%,其原因可能為DAL80抑制精氨酸酶編碼基因(如CAR1)的轉(zhuǎn)錄,而BLE也有相似的作用。DAL80敲除菌的Arg利用率和精氨酸酶活力均呈現(xiàn)相同的變化趨勢,這說明DAL80敲除菌中BLE略微影響Arg利用率和精氨酸酶活力,但敲除DAL80對二者的影響更突出,因此DAL80的敲除對釀酒酵母氮代謝的影響占主導(dǎo)作用。
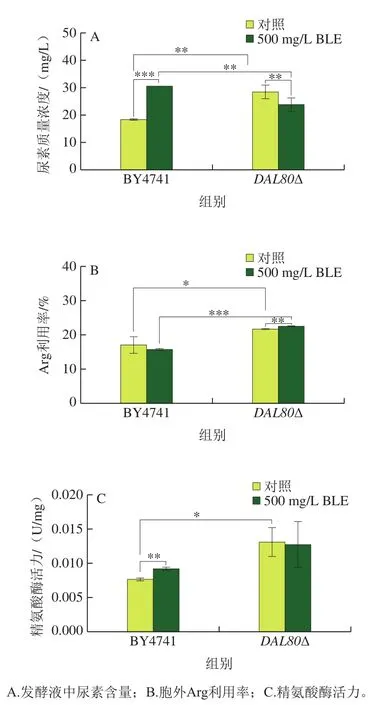
圖4 DAL80敲除菌和BY4741在BLE處理下的代謝水平Fig. 4 Metabolic levels of DAL80Δ and BY4741 in culture medium containing bamboo leaf extract
2.2.3DAL80敲除菌和BY4741中Arg及尿素代謝相關(guān)基因的表達(dá)水平
采用real-time PCR繼續(xù)檢測氮代謝相關(guān)基因的表達(dá)水平,可以從轉(zhuǎn)錄水平進(jìn)一步解釋DAL80敲除菌和BY4741的氮代謝水平差異。基因表達(dá)倍數(shù)差異表示為2-ΔΔCt,其中ΔCt為目的基因與內(nèi)參ACT1的Ct值差,ΔΔCt為目的基因在不同樣本中的ΔCt值差。BY4741菌相比于不含BLE的處理方式,在BY4741添加500 mg/L BLE與不添加BLE培養(yǎng)基中氮代謝有關(guān)基因轉(zhuǎn)錄水平差異倍數(shù)之比如圖5所示,DUR1,2、ALP1、GAP1的轉(zhuǎn)錄水平均有較顯著提高,GZF3和DAL82的轉(zhuǎn)錄水平顯著降低,DAL81的轉(zhuǎn)錄水平被略微抑制。其中,ALP1和GAP1的轉(zhuǎn)錄水平提高幅度最大,分別提高至2.7 倍和7.6 倍,這意味著BLE主要提高釀酒酵母的氨基酸透性酶編碼基因的轉(zhuǎn)錄表達(dá)水平。

圖5 BLE處理對BY4741氮代謝相關(guān)基因表達(dá)水平的影響Fig. 5 Effect of bamboo leaf extract on nitrogen metabolism-related gene expression levels in BY4741
如圖6所示,DAL80的敲除顯著提高GZF3、DUR1,2、DUR3、ALP1、DAL82、GLN3、CAR1、CAN1和GAP1的轉(zhuǎn)錄表達(dá)水平,說明這9 個基因均受到DAL80的調(diào)控,其中GZF3、DUR1,2、DUR3、ALP1的轉(zhuǎn)錄水平大幅提高,其分別提高至野生型酵母BY4741與四者對應(yīng)基因的29.0、23.9、42.1、25.3 倍。此前已有報道,DUR1,2、DUR3、DAL82、GAP1的轉(zhuǎn)錄水平受到DAL80的負(fù)調(diào)控作用,GZF3在阻遏型培養(yǎng)基上受到DAL80的負(fù)調(diào)控作用[21,26],而GLN3與DAL80間的相互調(diào)控作用值得關(guān)注。DUR1,2編碼尿素羧化酶和脲基甲酸鹽水解酶,釀酒酵母胞內(nèi)尿素的降解主要是在這2 種酶的連續(xù)作用下分解為二氧化碳和胺鹽;DUR3編碼尿素轉(zhuǎn)運酶,將分泌到胞外的尿素重吸收進(jìn)入胞內(nèi)由尿素轉(zhuǎn)運酶參與完成。DUR1,2和DUR3的大幅過表達(dá)利于酵母細(xì)胞內(nèi)尿素的降解,減少尿素從細(xì)胞內(nèi)轉(zhuǎn)移到發(fā)酵液中,有利于降低發(fā)酵液中EC含量[17,27]。CAR1編碼精氨酸酶,用來將Arg水解成為鳥氨酸和尿素[28];CAN1和ALP1分別編碼Arg透性酶和高親和力氨基酸透性酶[29]。CAN1、ALP1和GAP1的過表達(dá)利于Arg的分解和轉(zhuǎn)運[30],可能對細(xì)胞的Arg利用率有提高作用。在DAL80敲除菌中,DAL81的轉(zhuǎn)錄表達(dá)水平被大幅抑制,說明DAL81的表達(dá)可能需要DAL80的誘導(dǎo)。張偉平等[17]報道,Gln3p誘導(dǎo)DAL82的表達(dá),而DAL81p阻遏DAL82的表達(dá),在本研究中,敲除DAL80后,DAL81的表達(dá)水平被抑制,而GLN3和DAL82的表達(dá)水平顯著提高,且DAL82提高的倍數(shù)大于GLN3,其原因可能是GLN3誘導(dǎo)DAL82的表達(dá),DAL81抑制DAL82的表達(dá),當(dāng)誘導(dǎo)作用加強(qiáng),抑制作用減弱,則DAL82的表達(dá)水平提高得更多,與張偉平等[17]的研究結(jié)果一致。

圖6 DAL80敲除菌和BY4741氮代謝相關(guān)基因表達(dá)水平差異Fig. 6 Nitrogen metabolism-related gene expression levels in DAL80Δ and BY4741
在BLE處理下,DAL80敲除菌氮代謝相關(guān)基因表達(dá)水平普遍降低,其中較為顯著的是GAT1、DUR3、DAL81、DAL82、CAR1、CAN1、ALP1和GAP1。DAL80敲除菌在添加500 mg/L BLE與不添加BLE培養(yǎng)基中基因表達(dá)水平差異倍數(shù)結(jié)果如圖7所示,BLE的干預(yù)會削弱DAL80敲除的調(diào)控影響。

圖7 BLE處理對DAL80敲除菌氮代謝相關(guān)基因表達(dá)水平的影響Fig. 7 Effect of BLE on nitrogen metabolism-related gene expression levels in DAL80Δ
敲除釀酒酵母BY4741中的DAL80基因,同時在培養(yǎng)基中加入500 mg/L BLE,DAL80敲除菌在500 mg/L BLE培養(yǎng)基與BY4741在不含BLE培養(yǎng)基中基因表達(dá)水平差異倍數(shù)結(jié)果如圖8所示。盡管BLE降低部分基因的轉(zhuǎn)錄表達(dá)水平,GLN3、DUR1,2、DUR3、DAL82和ALP1的轉(zhuǎn)錄表達(dá)水平仍有明顯提高,說明與BLE的調(diào)控影響相比,轉(zhuǎn)錄因子DAL80在釀酒酵母細(xì)胞氮代謝中的抑制調(diào)控作用更為突出。
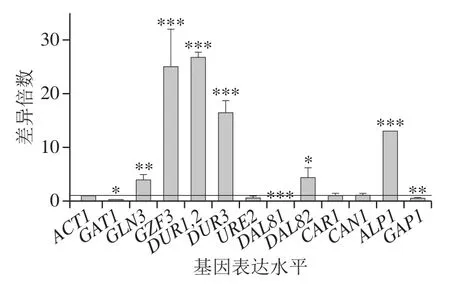
圖8 DAL80敲除和BLE處理對BY4741氮代謝相關(guān)基因表達(dá)水平的影響Fig. 8 Effect of DAL80 knockout and BLE on the expression of nitrogen metabolism-related genes in BY4741
3 結(jié) 論
本實驗研究BLE與釀酒酵母細(xì)胞GATA家族轉(zhuǎn)錄因子DAL80對黃酒釀造中釀酒酵母細(xì)胞Arg代謝網(wǎng)絡(luò)的調(diào)控及其互作影響。在培養(yǎng)釀酒酵母BY4741時,添加BLE可以降低Arg利用率,敲除DAL80會提高氮代謝有關(guān)基因(DUR1,2、DUR3、ALP1、DAL82、GLN3、CAR1、CAN1和GAP1)的轉(zhuǎn)錄表達(dá)水平。當(dāng)兩者同時作用時,BLE的干預(yù)會削弱DAL80敲除帶來的影響,但DAL80敲除對釀酒酵母氮代謝的影響仍占主導(dǎo)地位。
猜你喜歡
美與時代·美術(shù)學(xué)刊(2022年3期)2022-04-27 01:18:15
中學(xué)生數(shù)理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學(xué)生數(shù)理化·中考版(2020年10期)2020-11-27 01:59:48
火花(2019年12期)2019-12-26 01:00:28
中國生殖健康(2019年2期)2019-08-23 08:12:08
人大建設(shè)(2019年12期)2019-05-21 02:55:32
產(chǎn)品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
學(xué)苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
中國質(zhì)量與標(biāo)準(zhǔn)導(dǎo)報(2014年1期)2014-02-28 22:21:28
