鹽堿脅迫下香椿幼苗光合及抗逆生理特性
2021-09-04 03:16:24鞏志勇辛建華商小雨郭曉明王延平
西北植物學報 2021年7期
關鍵詞:水平
鞏志勇,辛建華,商小雨,郭曉明,王延平*
(1 黃河下游森林培育國家林業局重點實驗室,山東泰安 271018;2 山東農業大學林學院,山東泰安 271018;3 山東省菏澤市林業局,山東菏澤 274011)
土壤鹽堿化對農業生產構成重大威脅,培育和引種能適應鹽堿環境的優良植物材料是鹽堿地綠色開發的迫切需要[1-2]。中國鹽堿地面積約3600萬hm2,鹽堿地的開發利用已上升為國家戰略[3-4]。近年來,研究者對鹽堿條件下植物的形態結構[5-7]、生理特征[8-9]以及生物量分配[10-11]等方面進行了研究,篩選出一些鹽堿地適生的植物種類[12]。但是,鹽堿地植物選擇不但應注重良好生態適應性,還應注重經濟效益發揮。采用經濟型樹種資源更有助于提升鹽堿地開發綜合效益。
香椿(Toonasinensis)是楝科(Meliaceae)香椿屬(Toona)落葉喬木,其嫩芽可食,是中國著名的“樹上蔬菜”[13];此外,其木材材性好,又有“中國桃花心木”之稱,也是中國特有的珍貴樹種[14]。香椿在中國已有兩千多年的栽培歷史,隨著市場經濟的發展,高效益多功能的香椿被認為是一種極具發展潛力的生態經濟型樹種[15]。香椿耐干旱瘠薄[16],但對其耐受鹽堿能力的研究鮮見報道。本研究模擬黃泛沙地鹽漬化土壤環境[17],通過設置中性鹽和堿性鹽混合的鹽堿脅迫梯度處理,探究鹽堿脅迫對香椿幼苗光合及抗逆生理指標的影響,揭示鹽堿脅迫下香椿幼苗生理響應機制,以期定量評估香椿對鹽堿脅迫的耐受能力,從而為今后開展鹽堿地香椿栽培技術研究提供科學依據。
1 材料和方法
1.1 試驗材料
供試香椿種源來自于山東省濟南市章丘區盆崖村。經前期對比,該種源香椿椿芽產量高,香味濃郁,種子發芽率高,是較理想的菜用香椿資源。課題組于2019年10月中旬在該地區采集香椿母樹種子,翌年4月經催芽處理后播于山東農業大學林學試驗站溫室中,常規養護管理,以當年香椿幼苗進行盆栽試驗。
1.2 混合鹽堿脅迫處理
鹽堿土由鹽化土壤和堿化土壤組成,鹽化主要由 NaCl 和 Na2SO4引起,堿化主要由 Na2CO3和 NaHCO3引起[17]。因此,本試驗選用2種中性鹽NaCl、Na2SO4和2種堿性鹽NaHCO3、Na2CO3按一定比例混合制成處理液,通過基質澆灌模擬鹽堿土環境。其中,堿性鹽設置AⅠ(pH=7.16)、AⅡ(pH=8.47)、AⅢ(pH=9.91) 3個水平,同時各堿水平內設置S50(50 mmol/L)、S100(100 mmol/L)、S150(150 mmol/L)、S200(200 mmol/L) 4個鹽濃度,總計12個混合鹽堿處理組合;以無鹽無堿土壤作為對照(CK)。將上述配制的處理液澆灌香椿幼苗并計算土壤含鹽量 (表1)。由于組內pH值變化不大,采用各處理組的pH平均值表示各組的堿水平。
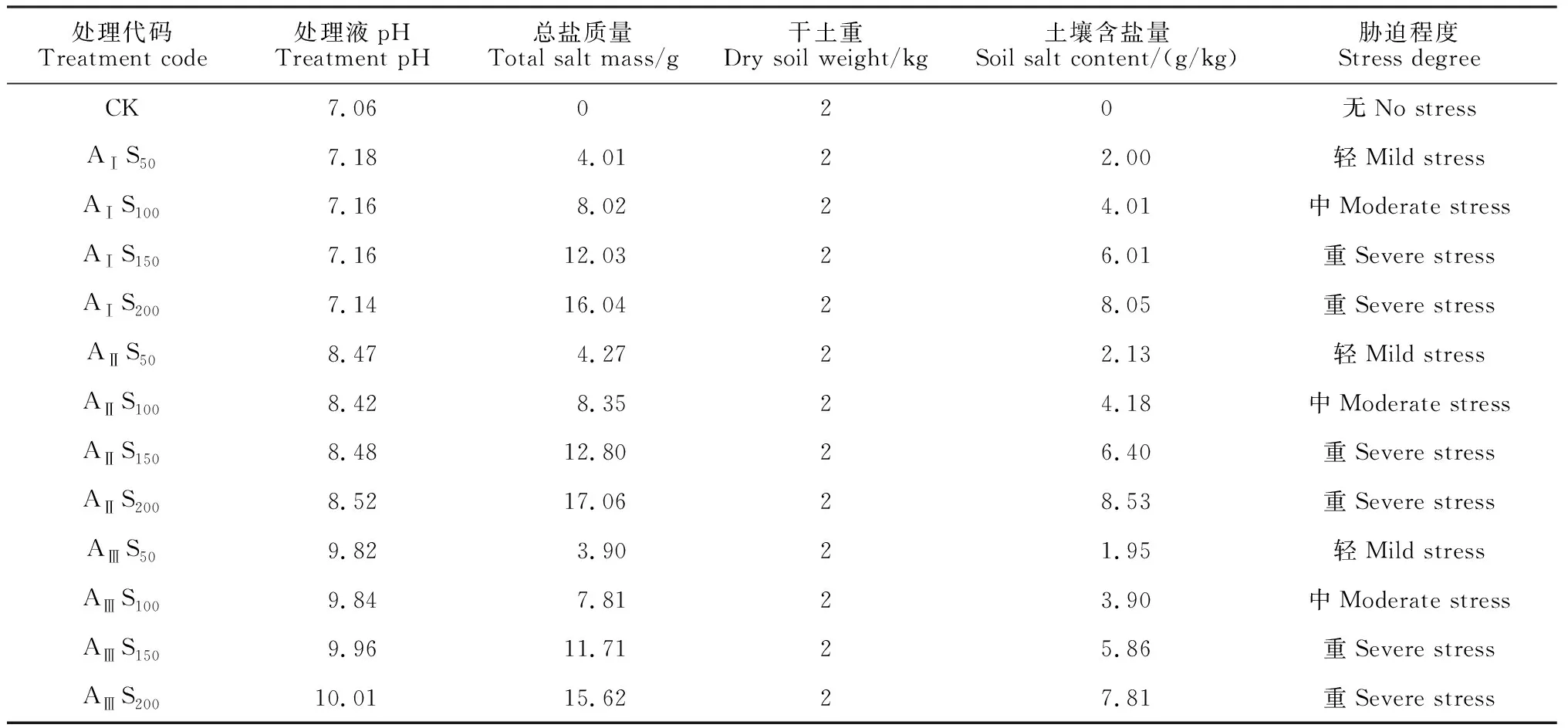
表1 鹽堿混合各處理組盆栽土壤鹽堿概況
稱取2.0 kg自然晾干栽培基質(壤土、細沙、蛭石按照體積比2∶1∶1混合土壤,容重1.307 g/cm3,pH 7.09,肥力中等)放入花盆中(盆直徑 25 cm,高 25 cm,盆底套上塑料盆防止鹽分流失)。量取上述各處理溶液800 mL,平均分為2份,連續2 d 加入基質中(為防止一次性加入鹽溶液質量分數過高導致植物出現燒苗現象)。其后,將各處理盆栽稱重,后期每天18:00前進行稱重補水,盡力保證盆栽土壤鹽堿水平穩定,脅迫持續35 d后進行相關指標的測定。
1.3 測定指標及方法
1.3.1 光合指標使用LI-6800 便攜式光合儀(LI-Cor Inc 美國),于上午 9:00~11:00取幼苗中上部相同部位3片葉,在飽和光強下(1 200 μmol·m-2·s-1,流速 500 mL·min-1,CO2濃度為400 μmol·mol-1,葉室溫度為28 ℃)測定凈光合速率(Pn)、氣孔導度(Gs)、胞間 CO2濃度(Ci) 、蒸騰速率(Tr) 等指標。每處理重復測3次,并計算得到瞬時水分利用效率(WUEi=Pn/Tr)。
1.3.2 光響應曲線選取幼苗中部生長良好的葉片為測定對象,采用光合儀自動模式進行測定,光照強度設置為 1 400、1 200、1 000、800、600、400、200、150、100、50、0 μmol·m-2·s-1。每次測定前進行一次匹配。凈光合速率(Pn)對光強的響應進程用葉子飄[18]報道的雙曲線修正模型進行擬合。根據擬合光響應曲線,計算最大凈光合速率(Pnmax)、光飽和點(LSP)、光補償點(LCP)、暗呼吸速率(Rd)。
1.3.3 生理生化指標每盆采集香椿幼苗復葉3片,將同處理的小葉混合成待測樣品。參照李合生的方法[19],95%乙醇浸提法測定葉綠素含量,氮藍四唑比色法測定超氧化物歧化酶(SOD)活性,愈創木酚比色法測定過氧化物酶(POD)活性,酸性茚三酮比色法測定脯氨酸(Pro)含量,考馬斯亮藍 G-250 比色法測定可溶性蛋白含量,蒽酮比色法測定可溶性糖含量,硫代巴比妥酸法測定丙二醛(MDA) 含量。
1.4 數據處理
采用 Excel 和 SPSS軟件進行數據分析,用Origin軟件繪制統計圖。
2 結果與分析
2.1 鹽堿脅迫對香椿幼苗地上部形態特征的影響
由圖1可看出,在S50鹽濃度下,各堿水平處理的香椿幼葉葉片舒展、濃綠,植株整體長勢良好,與對照差異不明顯;在S100鹽濃度下,隨堿水平的逐漸增加,植株葉尖逐漸發黃、干枯,但總體長勢尚可;而在S150、S200兩個鹽濃度下,各堿水平處理的香椿葉片大部分發黃、枯萎甚至開始脫落,植株長勢較差,尤其在AⅡ和AⅢ堿水平處理下,葉片大量脫落。上述地上部形態特征說明,鹽堿脅迫對香椿幼苗生長具有顯著影響,其中,香椿幼苗地上部生長對堿水平的響應強于鹽脅迫,且隨脅迫程度增加鹽堿交互效應越明顯。

圖1 鹽堿脅迫下香椿幼苗地上部形態特征Fig.1 Aboveground morphological characteristics of T. sinensis seedlings under saline-alkali stress
2.2 鹽堿脅迫對香椿幼苗葉片光合氣體交換參數的影響
隨著鹽堿濃度的增加,香椿幼苗葉片凈光合速率、蒸騰速率、氣孔導度和瞬時水分利用效率總體呈現出下降趨勢,而胞間二氧化碳濃度有上升趨勢(圖 2)。其中,AⅠS50處理下,香椿幼苗葉片凈光合速率、蒸騰速率、氣孔導度、水分利用效率與對照相比無顯著差異(P> 0.05),而胞間二氧化碳濃度顯著低于對照(P< 0.05);但隨堿水平增加,AⅡS50和AⅢS50處理香椿凈光合速率比對照分別顯著下降36.5%和24.6% (P< 0.05),而它們的蒸騰速率、胞間二氧化碳濃度、瞬時水分利用效率與對照相比仍沒有顯著差異(P> 0.05)。在AⅠS100處理中,除胞間二氧化碳濃度顯著增加外,香椿幼苗葉片各光合指標與對照均沒有顯著差異(P> 0.05);但隨堿水平增加,AⅡS100和AⅢS100處理香椿葉片凈光合速率、氣孔導度、瞬時水分利用效率均顯著低于對照(P< 0.05);而蒸騰速率和胞間二氧化碳濃度在AⅡ堿水平下與對照無顯著差異(P> 0.05),而在AⅢ堿水平下差異達顯著水平(P<0.05)。在S150和S200兩個鹽濃度下,各堿水平下的香椿葉片光合參數與對照相比均顯著下降(P<0.05)。可見,堿水平對香椿幼苗葉片光合參數影響更大,其次是鹽濃度水平。
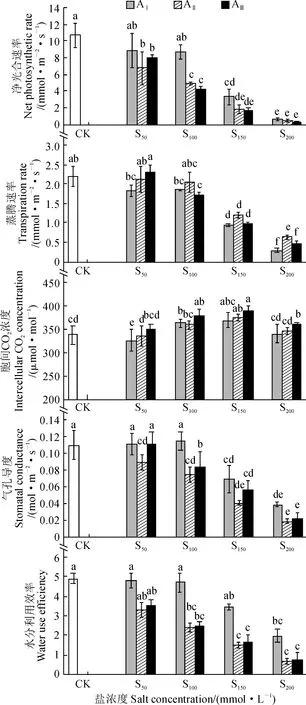
圖中數據為平均值 ± 標準差,不同小寫字母表示在0.05水平差異顯著(P<0. 05),下同圖2 鹽堿脅迫下香椿葉片光合參數的變化The data are mean ± standard deviation. The different lowercase letters on the error bar mean significant difference between treatments and CK at 0.05 level (P<0. 05). The same as belowFig.2 The changes of photosynthetic parameters in leaves of T. sinensis seedling under saline-alkali stress
2.3 鹽堿脅迫下香椿幼苗的光響應特征
利用直角雙曲線模型分別對不同鹽堿脅迫下香椿幼苗的光合-光響應曲線進行擬合,各方程的擬合系數均在 0.97以上(表2),能較好地反映不同鹽堿脅迫條件下香椿葉片的光響應過程。各鹽堿脅迫處理香椿幼苗葉片Pn隨光合有效輻射 (PAR)的增加均逐漸上升,當PAR增加到一定程度(達到光飽和點)后開始下降(光抑制)并逐漸趨于平穩(圖3)。其中,在AⅠ-ⅢS50、AⅠS100處理下,隨PAR的增加,香椿幼苗葉片Pn上升速率較快且最大凈光合速率高,其光合能力未受顯著影響;而在AⅡ-ⅢS150、AⅠ-ⅢS200處理下,隨著PAR的增加,葉片Pn上升速率變慢且最大凈光合速率趨近于0,幼苗的光合效率極低。
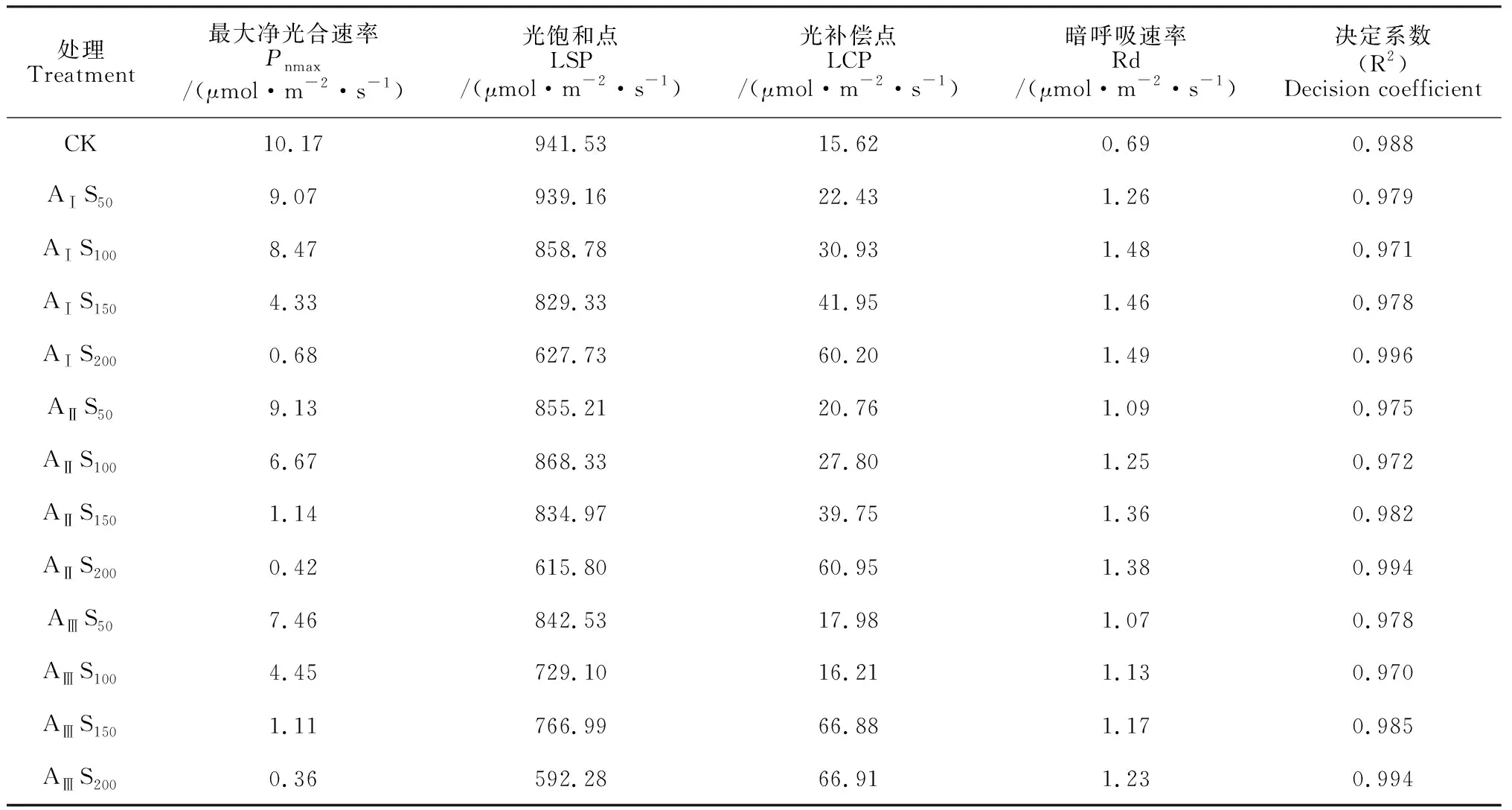
表2 不同鹽堿脅迫下光響應模型擬合的特征參數
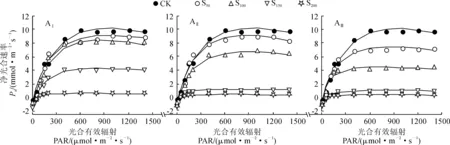
圖3 鹽堿脅迫下香椿凈光合速率隨光合有效輻射的變化Fig.3 Changes of net photosynthetic rate with photosynthetically active radiation under saline-alkali stress
同時,從表2可知,隨著鹽堿脅迫程度的逐漸增加,香椿幼苗的最大凈光合速率(Pnmax)、光飽和點(LSP)均呈現出不同程度的下降,而其光補償點(LCP)、暗呼吸速率(Rd)總體呈上升趨勢。其中,在AⅠ堿水平下,AⅠS50和AⅠS100處理葉片Pnmax分別低于對照10.84%、16.68%,LSP分別低于對照0.25%、8.79%,總體上保持著較高的光能利用率;在AⅡ堿水平下,僅AⅡS50處理葉片Pnmax低于對照10.23%,LSP未見明顯下降;而在AⅢ堿水平時,只有AⅢS50處理香椿幼苗葉片Pnmax和LSP比對照分別降低26.65%和10.51%,還維持較高的光能利用率。以上結果說明香椿幼苗光能利用效率隨鹽堿脅迫的加重而逐漸降低。
2.4 鹽堿脅迫對香椿葉片葉綠素含量的影響
表3顯示,在AⅠ堿水平下,香椿幼苗葉片各葉綠素含量總體上隨著鹽脅迫程度的增加而呈現出先升高后降低的趨勢,并均在S100時達到最大值;葉綠素a、葉綠素b及葉綠素總含量在鹽濃度為S100時分別比對照顯著增加4.46%、133.80%和44.73%,葉綠素a和葉綠素總含量在S200時分別比對照顯著降低38.22%和24.56%,各葉綠素含量在其余鹽濃度脅迫下均與相應對照無顯著差異。在AⅡ堿水平下,各葉綠素含量均隨著鹽濃度的增加而逐漸降低,在鹽濃度為S50和S100時均不同程度地高于對照,且S100時葉綠素總含量增幅達到顯著水平;在鹽濃度達到S150和S200時均不同程度地低于對照,并在S200時降幅均達到顯著水平,分別為39.49%、54.93%和48.68%。在AⅢ堿水平下,各葉綠素含量均隨著鹽濃度的增加而基本呈逐漸降低趨勢,在鹽濃度為S50時均不同程度地高于對照,且葉綠素a和葉綠素總含量均達到顯著水平;在其余鹽濃度處理下,各葉綠素含量均顯著低于對照(P< 0.05)。可見,香椿幼苗葉片葉綠素含量總體呈現對堿脅迫較為敏感,葉綠素a含量的變化幅度又大于葉綠素b。
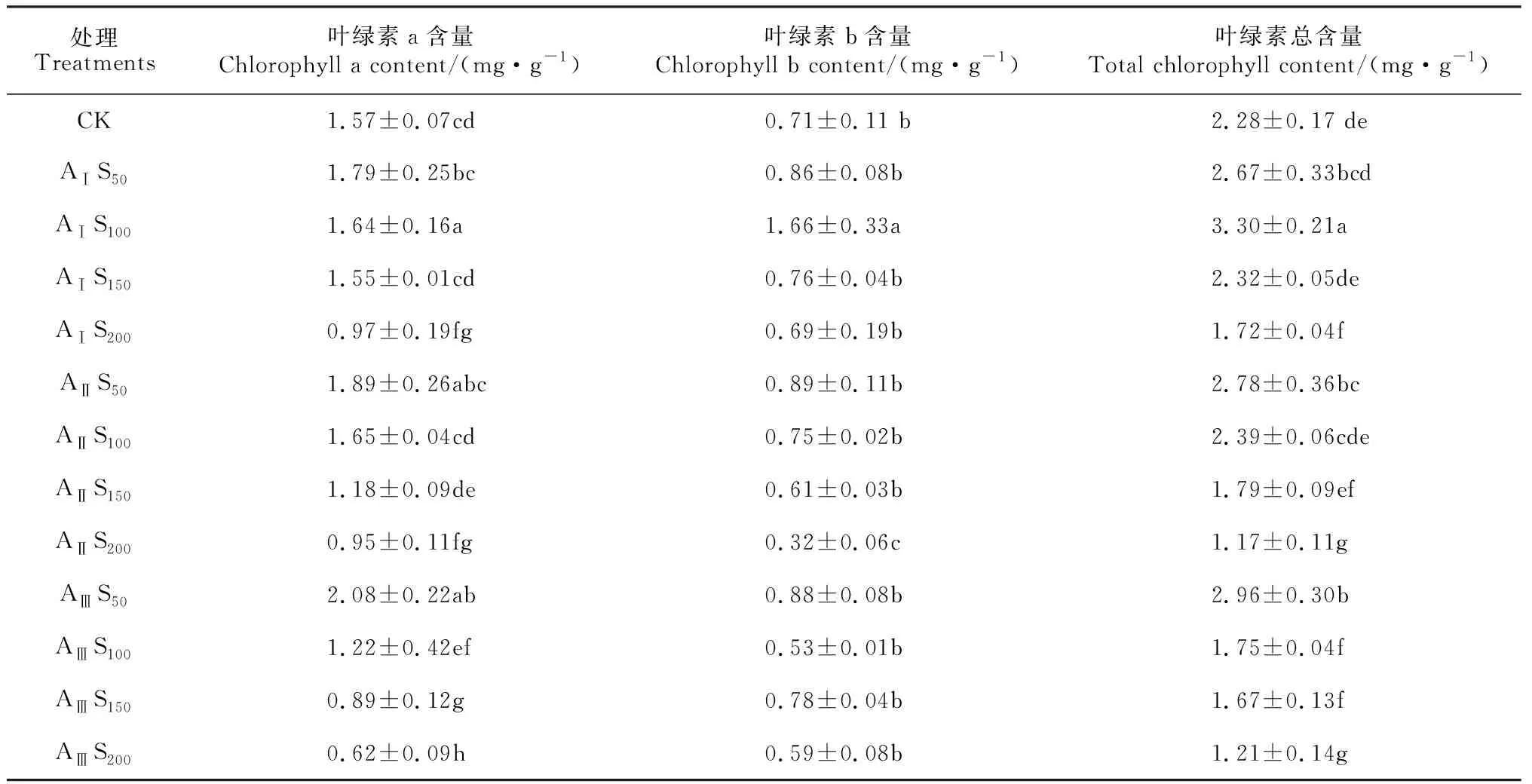
表3 鹽堿脅迫下香椿葉片葉綠素含量的變化
2.5 鹽堿脅迫對香椿幼苗細胞膜透性、丙二醛含量及抗氧化酶活性的影響
從圖4可以看出,各鹽堿處理香椿幼苗葉片相對電導率均顯著高于對照;在相同堿水平下,隨著鹽濃度的增加,香椿葉片相對電導率逐漸增大;各堿水平下的葉片相對電導率在S50、S100鹽濃度處理時均低于40%,而在S150、S200鹽濃度處理時均高于60%。香椿幼苗葉片丙二醛(MDA)含量在S50鹽濃度處理時隨堿水平的增加而逐漸降低,但在AⅠ堿水平下顯著高于對照;在S100、 S150、S200三個鹽濃度處理下,葉片MDA含量隨堿水平增加逐漸升高并顯著高于對照(P< 0.05)。上述結果說明,鹽堿脅迫容易導致香椿葉片細胞膜受損進而可能影響正常細胞代謝功能。

圖4 鹽堿脅迫下香椿幼苗葉片相對電導率及丙二醛含量的變化Fig.4 Relative electrical conductivity and MDA content in leaves of T. sinensis seedlings under saline-alkali stress
另外,隨著鹽濃度增加,各堿水平處理香椿幼苗葉片的超氧化物歧化酶(SOD)活性呈先上升后下降的趨勢(圖5),并分別在S100(AⅠ)和S50(AⅡ、AⅢ)達到最大值;除AⅢS150處理葉片SOD活性與對照相近,以及AⅠ-ⅢS200處理葉片SOD活性顯著低于對照外,其余鹽堿處理SOD活性均顯著高于對照(P< 0.05);AⅠS100處理葉片SOD活性在各鹽堿處理中最高,顯著高于對照71.7%(P< 0.05)。同時,隔各鹽堿處理香椿幼苗葉片過氧化物酶(POD)活性隨著鹽(堿)濃度的增加均呈現逐漸上升的趨勢,并均顯著高于對照(P< 0.05)。說明香椿葉片SOD活性在中低鹽脅迫水平受到顯著誘導,而在高鹽濃度下(S200)受到抑制,而葉片POD活性各鹽堿脅迫下都得到顯著增強,即香椿幼苗可以通過調控抗氧化酶活性來適應鹽堿脅迫環境。
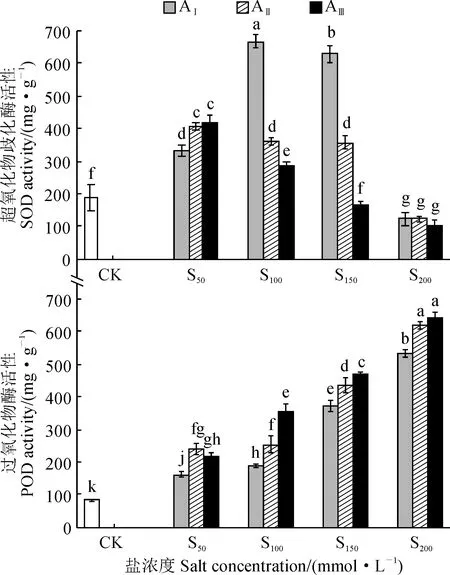
圖5 鹽堿脅迫下香椿幼苗抗氧化酶活性的變化Fig.5 The antioxidant enzyme activity in leaves of T. sinensis seedlings under saline-alkali stress
2.6 鹽堿脅迫對香椿幼苗滲透調節物質含量的影響
圖6顯示,隨鹽堿脅迫濃度的增加,香椿幼苗葉內脯氨酸含量逐漸增加,而葉片可溶性蛋白和可溶性糖含量均先升高后下降,并均在鹽濃度為S100或者S100時達到最大值。其中,香椿幼苗葉內脯氨酸的含量在各鹽堿處理下均顯著高于對照(P< 0.05);各堿水平下的葉片可溶性蛋白含量均在鹽濃度S100處理時達到最大值且顯著高于對照(P< 0.05),AⅠ、AⅡ、AⅢ堿水平增幅分別為63.18%、55.93%、58.04%;各堿水平下葉片可溶性糖含量在S50、S100、S150鹽濃度處理時也均顯著高于對照(P< 0.05)。以上結果說明香椿幼苗在鹽堿脅迫時仍可以通過調控滲透調節物質含量來適應鹽堿脅迫環境。
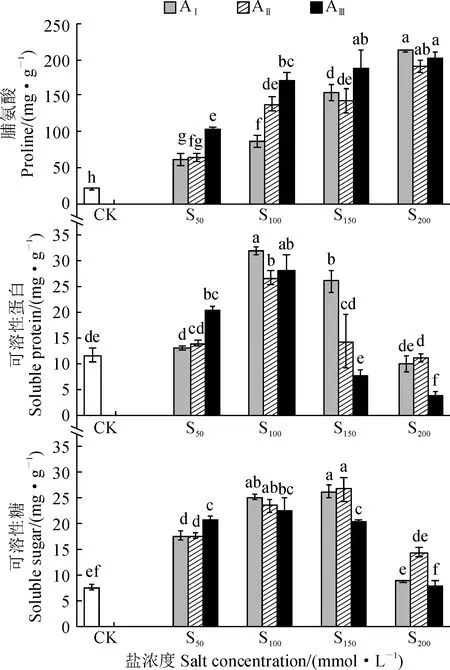
圖6 鹽堿脅迫下香椿幼苗滲透調節物質含量的變化Fig.6 The content of osmotic regulation substances in leaves of T. sinensis seedlings under saline-alkali stress
3 討 論
3.1 香椿對鹽堿脅迫的光合響應
光合作用對植物體生長發育和生態適應均具有重要意義[20-21],葉片光合參數是衡量植物光合效率的重要指標[22]。通常情況下,鹽堿逆境通過滲透脅迫、離子毒害等過程影響植物光合能力[23]。研究表明,高鹽堿脅迫下植物葉片凈光合速率、氣孔導度、蒸騰速率等光合參數都會下降[24]。其中,氣孔導度的下降意味著植物從環境中吸收CO2的能力降低,從而引起光合碳同化能力減弱[25];而蒸騰速率下降可以減緩植物對鹽分離子的吸收。本研究發現,香椿幼苗光合參數凈光合速率、蒸騰速率、水分利用效率、最大凈光合速率等在鹽濃度為S50處理下受堿水平的脅迫影響較小,其光合效率較高;而當鹽濃度為S100時,AⅠ水平下葉片光合效率仍然較高,而AⅡ和AⅢ水平下光合效率明顯下降,這說明隨著鹽濃度逐漸增加,香椿幼苗葉片光合參數受堿脅迫的影響加大,鹽堿交互脅迫開始逐漸增強,香椿幼苗的光合能力受到明顯抑制。這與劉建新等[26]在燕麥幼苗上的研究結果一致。此外,有研究表明,影響植株Pn降低的因素分為氣孔因素(凈光合速率、氣孔導度及胞間二氧化碳濃度同時下降)和非氣孔因素(凈光合速率、氣孔導度下降,胞間二氧化碳濃度增加)[27-28],從本研究結果來看,在鹽堿脅迫條件下,非氣孔因素可能是影響香椿幼苗光合能力下降的主導因素。
葉綠素是植物進行光合作用的重要光合色素,參與光合作用中光能的吸收、傳遞和轉化功能,其含量的多少能夠直接反映葉片光合能力的強弱[29]。鹽堿脅迫會破壞葉片葉綠體結構,打破葉綠素合成與降解的動態平衡,從而導致光合作用受抑制[30-31]。本研究發現,在堿水平為AⅠ時,香椿幼苗葉片內葉綠素含量在鹽濃度為S150處理中仍然保持著較高水平,而當堿水平繼續逐漸升高時,葉綠素含量明顯下降。這可能是由于較高的pH值破壞了土壤中離子的穩態,使與葉綠素合成有關離子如 Fe2+、Cu2+、Mg2+等沉淀,進而打破了葉綠素合成與分解的動態平衡,從而導致葉綠素含量降低。此外,當堿度較低時,葉綠素 b 含量開始下降的鹽濃度值要低于葉綠素 a,說明葉綠素 a對鹽堿混合脅迫的敏感性高于葉綠素 b。
3.2 香椿幼苗在鹽堿脅迫下的抗逆生理生化特征
細胞膜承擔著調節和控制細胞內外正常的物質運輸和交換的任務,主要由磷脂和蛋白質組成,具有選擇透過性[32]。鹽堿脅迫導致細胞膜結構受損,膜透性增大,細胞內的電解質外滲,繼而導致電導率升高。因此,相對電導率可表征植物受鹽堿脅迫傷害的程度,即相對電導率越大,則細胞膜透性越大[33]。此外,植物細胞膜系統在遭受鹽堿脅迫后還會引起膜脂過氧化反應[34],其中丙二醛(MDA)作為膜脂過氧化反應的主要產物之一,其含量的多少在一定程度上反映出膜損傷程度的大小[35]。本研究中,隨著鹽堿脅迫增加,香椿葉片相對電導率、MDA含量逐漸上升,但當鹽濃度為S50和S100處理時,各堿水平下的相對電導率、MDA與對照相比增加較少且含量較低;而當鹽濃度增加到S150和S200處理時,二者的含量明顯增加。這說明當鹽濃度較低時,香椿葉片細胞膜的損傷程度較小,隨著鹽濃度繼續增加,膜結構嚴重受損。
同時,逆境脅迫下植物體內抗氧化酶活性會增強或維持較高的水平,這對于植物適應逆境和減少傷害起著重要作用[36]。其中,SOD有助于將氧自由基團轉變為H2O2和O2,減輕自由基對植物體細胞的危害;而POD 則具有去除H2O2和酚類等毒性物質的重要功能[37-38]。本研究中,隨著鹽濃度增加,各堿水平下香椿葉片SOD活性呈先上升后下降的趨勢,POD活性亦呈上升的趨勢。這說明香椿可通過自身調節,誘導增強SOD和POD活性來保護細胞膜免受傷害。
另外,植物在代謝過程中通過脯氨酸、可溶性蛋白、可溶性糖等滲透調節物質的合成和積累來緩解脅迫對植物造成的傷害[39-40]。其中,脯氨酸可以作為細胞質的滲透調節物質和防脫水劑,參與維持細胞的含水量和膨壓,增強細胞結構的穩定性[41-42]。可溶性糖和可溶性蛋白是植物在逆境下重要的滲透調節物質[43],當植物受到逆境脅迫時,進行滲透調節從而減輕植物的受傷害程度。通常情況下,因植物種類不同,參與滲透調節的物質以及調節強度會產生差異[44]。本研究表明,可溶性糖、可溶性蛋白隨著鹽堿脅迫增大,其含量先增加后降低。其中,在鹽濃度為S100處理時,各堿水平下的可溶性蛋白含量均達到最大值,可溶性糖含量在S50、S100、S150鹽濃度處理時,各堿水平下的含量也均顯著高于對照。這說明說明香椿幼苗可在一定程度上通過積累可溶性蛋白,可溶性糖等大分子物質來抵御鹽堿脅迫。
綜上所述,在本試驗條件下,香椿幼苗對堿脅迫表現出比鹽脅迫更強的敏感性;當土壤pH <8.4,鹽含量<4.0 g/kg時,香椿幼苗地上部整體長勢良好,葉片組織內的抗氧化酶活性、滲透調節物質仍能高效的發揮作用,光合效率也維持在較高水平。因此,香椿能夠耐受一定程度的鹽堿,具備在鹽堿地上栽培的潛能。
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
