鄉土物種補播對青藏高原高寒草甸群落穩定性的影響
2021-09-07 08:28:42段麗輝劉曉麗位曉婷才仁措邵新慶
草地學報 2021年8期
關鍵詞:物種
段麗輝, 劉曉麗, 韓 冰, 位曉婷, 才仁措, 邵新慶,3*
(1.中國農業大學草業科學與技術學院,北京 100193; 2.玉樹州農牧業綜合執法局, 青海 玉樹 815000;3.青海省寒區恢復生態學重點實驗室, 青海 西寧 810008)
青藏高原位于我國西北部[1]。其隆升對中國乃至亞洲的地勢地貌格局、大江大河發育、氣候條件形成均產生了深遠影響,逐漸發育形成了獨特的自然生態系統。由于該區域生態環境脆弱[2],原生植被敏感,因此植物群落受影響后自我修復和調節能力弱,極易受到干擾[3]。青藏高原植被類型豐富,以高寒草甸植被為主[4]。近年來,該區域草地出現了不同程度的退化,毒雜草叢生、植被退化、鼠害猖獗等問題頻發[5],對當地牧民生產生活造成了極為惡劣的影響。
隨著人們科學眼光的發展以及生活物質資料的日益豐富,草地保護、合理利用及科學管理得到了越來越多的重視。草地補播作為改良退化草地的有效途徑[6],因為其成效顯著、效率高而備受關注:吳宛萍[7]在寧夏荒漠草原選用鄉土物種建植人工混播草地,使得草地群落地上生物量增加,群落穩定性顯著提高(P<0.05);周冀瓊[8]在呼倫貝爾退化羊草(Leymuschinensis(Trin.) Tzvel.)草地利用紫花苜蓿(Medicagosativa)及黃花苜蓿(MedicagofalcataL.)進行免耕補播,使草地生態系統的多樣性和物種豐富度維持在較高水平,同時使草地生產力大幅提升;馬和等[9]在甘肅省肅南縣對因去除毒雜草后重度貧瘠的草地實施淺翻耕后補播,使得草地植被密度迅速升高,地上生物量顯著增加(P<0.05)。關于補播鄉土物種對于群落穩定性及種間聯結性的研究較少。
為了篩選出適應青藏高原生境的生態恢復物種組合,本試驗采用種間聯結性及群落穩定性相結合的分析方法,選取垂穗披堿草(Elymusnutans)、異針茅(Stipaaliena)、中華羊茅(Festucasinensis)、溚草(Koeleriacristata)、星星草(Puccinelliatenuiflora)、扁蓿豆(Melissitusrutenica)、冷地早熟禾(Poacrymophila)共7種鄉土物種,以5種混播組合在青海省海南藏族自治州高寒草甸天然草地進行補播。本試驗為多年生草地的建植、經營與管理體系奠定了理論基礎,為高寒草甸天然草地的生態保護和資源合理利用提供了理論支持。
1 材料與方法
1.1 研究區概況
本試驗在青海省海南藏族自治州貴南縣草原站(簡稱貴南站)(100°13′~101°33′E,35°09′~36°08′N)[10]的野外試驗樣地進行。該樣地于2016年圍封,長140 m,寬120 m,面積約1.9 ha。該地區屬于高原大陸性氣候,冬寒夏涼,年平均氣溫為2.3℃,雨熱同期,干濕季分明而四季不分明,年降水量為403.8 mm,年平均蒸發量為1 378.5 mm[11]。植被類型為高寒草甸,土壤類型為高山草甸土[12]。該區域主要優勢種包括克氏針茅(Stipakrylovii)、垂穗披堿草、矮嵩草(Kobresiahunilis)、溚草和紫花針茅(Stipacapillata)[13]。
1.2 研究方法
1.2.1試驗設計與方法 本試驗于2017年開展試驗,選取垂穗披堿草、異針茅、中華羊茅、溚草、星星草、扁蓿豆、冷地早熟禾共7個鄉土物種,搭配成5種混播組合,具體物種搭配見表1。

表1 混播組合物種組成Table 1 Species composition in combination
試驗采用隨機區組試驗設計方案,將5種處理設置4組重復,共20個小區,單個小區面積為:5 m×4 m=20 m2,各小區間距為1 m,小區外圍設有寬為1 m的保護行。在各個小區內沿長邊方向將草皮劃破,劃溝深度為0.5~1.5 cm,行距25 cm為宜,以單個小區內分布16~17條劃溝為宜。將7個草種分別依照扁蓿豆0.6 kg·km-2、異針茅0.6 kg·km-2、溚草0.325 kg·km-2、中華羊茅0.6 kg·km-2、星星草0.325 kg·km-2、垂穗披堿草1.5 kg·km-2、冷地早熟禾0.6 kg·km-2的標準,于2017年6月13日播種至相應小區內(組合E為空白對照,故僅需要沿長邊劃破草皮,不做補播處理。)為確保種子順利發芽,播種后要覆土并踩實,無額外水肥投入。
1.2.2植物群落特征調查方法 對試驗區的植物群落進行調查,將植物鑒定到種,根據Padisák等[14]提出的功能群分類方法劃分出5個主要的功能群。此外,每個小區內,在避開單優植物斑塊及邊緣地塊的基礎上,隨機選擇3個面積為0.25 m2(0.5 m×0.5 m)的小樣方,于2017—2018連續2年8月上旬對蓋度、高度、多度和地上生物量進行測定。各指標測定方法及要求如下:蓋度:利用網格法,得到群落總蓋度及每個物種的分蓋度[10];高度:每種植物隨機選擇10株并用卷尺測得高度,不足10株的物種則測定全部株的高度;多度:直接記錄樣方內不同種植物的株數;地上生物量:在測定得到蓋度、高度和多度的基礎上,對樣方內不同植物種進行齊地面刈割,收集刈割得到的地上部分,裝于信封袋中,置于65℃~80℃烘箱烘干,稱重得到的干重即為地上生物量大小[13]。
1.3 測定指標計算方法
1.3.1群落種間聯結性分析 一個植物群落中有多種植物共存,各個種群之間都產生聯系、相互作用,我們將這種關系稱為種間聯結[15]。群落內不同種群間的關聯性(或稱為聯結性)不僅能夠客觀地反映出群落內不同物種之間互作關系的有無及緊密程度[16],還能夠直觀地反映出群落的結構特征[17]并指示其演替方向[18]。本試驗采用方差比率法(Variance ratio,VR)[19]定量測定植物群落的總體關聯性。其公式如下:
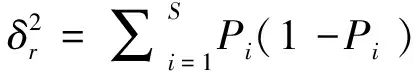
(1)
(2)
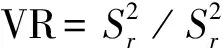
(3)
S:物種總數;Pi:包含物種i的樣方數;N:總樣方數;Tj:樣方j的物種總數;t:樣方包含物種數的均值;VR:群落內物種間的總體關聯指數。
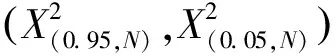
1.3.2群落穩定性分析 試驗采用M.Godron貢獻定率法[21](Contribution law)分析群落穩定性的變化情況。首先計算群落中所有物種的頻度,并將物種按照頻度從大到小的順序排列,換算成相對頻度后逐步累積,得到累積相對頻度,記作y;隨后將樣地包含植物種類總數取倒數,依照物種排列順序逐步對應累積,得到植物種類累積倒數百分率,記作x;最后利用成對數據擬合出一條平滑曲線,并與給定直線y=100-x相交,交點比值即可表征群落穩定性的大小[22]。一般來說,交點比值為20/80時群落穩定性最高,因此計算得到的交點坐標越接近20/80,群落就越穩定[22]。
平滑曲線模擬模型:y=ax2+bx+c
(4)
直線方程:y=100-x
(5)
將(5)代入(4)得:ax2+(b+1)x+c-100=0
(6)
交點坐標為:
(7)
y=100-x
(8)
y:物種的累積相對頻度,即,將植物種類按頻度按從大到小順序排列,將頻度轉化為相對頻度并進行累積;x為物種總數倒數的累積,即將植物種類總數取倒數并依次累積;a,b,c均為參數,算法及取值參考黃祥童[22]方法。
1.3.3生物量穩定性分析 群落生物量是群落的生長發育情況的直觀表征[23],也是衡量群落穩定性大小的重要指標[24]。本試驗根據Tilman[25]提出的時間穩定性計算方法計算群落的生物量穩定性。具體公式如下:
S=μ/σ
(9)
μ:群落中各個種群生物量的均值之和;σ:群落中各個種群生物量的標準差之和。
生態學界對于群落生物量的時間穩定性叫法各異,如時間序列穩定性[26],生物量穩定性[27]等。為了避免引起歧義,以下在文中一致稱為生物量穩定性,記作S。但不論采取哪一種叫法,其本質都與生物統計學中的變異系數一樣,能夠用于不同單位、不同尺度下一組觀測值變異程度的比較[27]。本試驗中還把這一指標的計算方法延伸到了功能群生物量穩定性的計算中,用于探究群落生物量穩定性與功能群生物量穩定性的關系。
1.4 數據處理
采用Excel 2016進行數據處理、計算及繪圖,利用SPSS 21.0軟件進行統計分析,利用單因素方差分析(One-way ANOVA)方法及LSD法在顯著性水平a=0.05下進行顯著性檢驗,不同小寫字母表示結果間存在顯著差異(P<0.05),利用Sigmaplot 12.5進行回歸分析并繪制圖像。
2 結果與分析
2.1 植物群落物種組成
植物群落調查結果表明,試驗區共有草本植物34種,分屬于11個科(表2)。其中常見科包括禾本科(Grass)、菊科(Composite)、豆科(Legume)、莎草科(Sedge)占比超過80%。

表2 群落物種組成Table 2 Species Structure of community
2.2 種間聯結性
對5種混播組合的植物群落總體關聯性進行分析,結果表明(表3):2017年僅組合A的群落總體表現為顯著正聯結;其余4個組合植物群落總體均表現為不顯著正聯結。2018年,組合B、組合C和組合E群落總體表現為不顯著負聯結;組合A、組合D群落物種總體表現為不顯著正聯結。
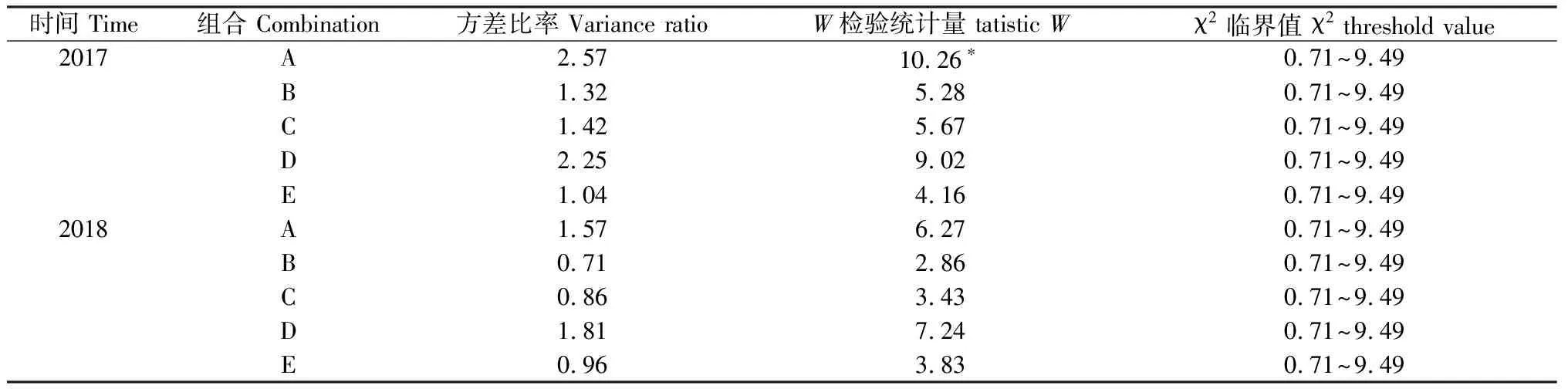
表3 群落物種聯結性計算結果Table 3 Results to community species association
2.3 植物群落穩定性
由圖1可知,2017年及2018年5種混播組合整體群落的種類累積倒數百分率x與累積相對頻度y的比值均遠離20/80。但每個組合先后兩年的補播結果相比之下,2018年的圖像交點更接近20/80(圖1)。
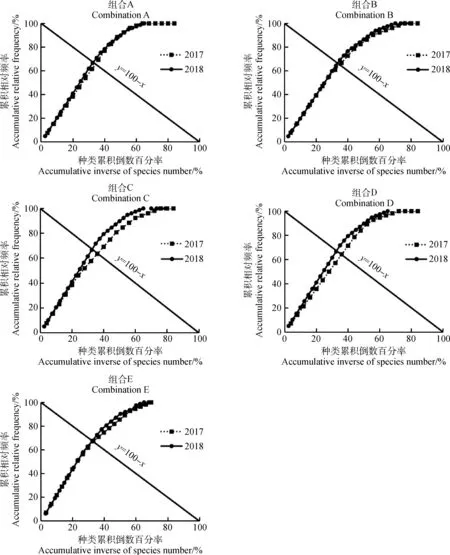
圖1 不同組合群落穩定性的分析Fig.1 Analysis of community stability of different combinations
2.4 群落生物量穩定性
試驗結果表明,5種混播組合的群落生物量穩定性排序為D>A>C>E>B。其中組合B與組合D的差異顯著(P<0.05),其余組合兩兩之間不存在顯著差異(圖2a)。
本試驗根據Padisák等[14]提出的功能群分類方法,將觀察到的物種劃分為禾本科、豆科、菊科、莎草科、雜類草(Forb)5個功能群。采用Tilman的時間穩定性計算方法[25]對功能群的生物量穩定性和群落整體的生物量穩定性進行計算,并建立直線回歸方程進行相關性分析。結果表明(圖2b~2f):群落生物量穩定性與禾本科生物量穩定性之間存在顯著的相關關系(R2=0.633,P<0.01),而與莎草科、菊科、豆科、雜類草的生物量穩定性之間均不存在顯著的相關關系(豆科:R2=0.19,P=0.053;菊科:R2=0.044,P=0.376;雜類草:R2=0.81,P=0.223;莎草科:R2=0.002,P=0.947)。
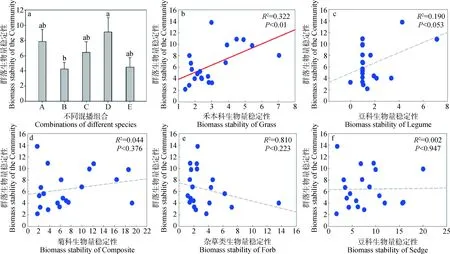
圖2 群落生物量穩定性與功能群生物量穩定性的關系Fig.2 Relationship between biomass stability of the community and biomass stability of the functional group
3 討論
3.1 種間聯結性與群落穩定性
種間聯結性是群落穩定性的長期背景,群落穩定性是種間聯結性的階段性表達[22]。通常情況下,種間正關聯程度越大,群落越穩定,種間關系越緊密,相互促進越多;相反,種間負關聯程度越大,種間競爭越激烈,群落穩定性越小[28]。有關種間聯結性與群落穩定性間相互關系的研究有很多:龐曉攀等[29]對高原鼠兔有效洞口數密度,同高山嵩草(KobresiapygmaeaC. B. Clarke)草甸植物群落種間聯結性之間的關系進行了一系列研究,發現植物群落主要物種總體聯結性由正聯結轉變為負聯結時對應的有效洞口數密度下,嵩草草甸植物群落穩定性最高;張鮮花等[30]對天山北坡不同海拔地段鴨茅(DactylisglomerataL.)群落主要物種進行了一系列研究,發現隨海拔升高,鴨茅群落主要物種種間聯結性由正關聯逐漸過渡到負關聯,同時群落穩定性上升到一定程度后減弱。本研究觀察到,5種混播組合所得群落整體種間正關聯程度降低而負關聯程度升高的同時,群落穩定性升高。這與上述研究[29-30]結論一致,推測群落接下來將會向著頂級群落方向發生演替,其穩定性也將會進一步上升,并在群落總體種間聯結性由正關聯轉變為負關聯時達到最大。
對于群落種間正關聯程度降低這一現象,可能是因為試驗選用的混播組合較簡單,且可利用資源與營養在試驗初期較充沛,因此物種間競爭較弱,從而表現出較好的種間相容性,繼而呈現出較好的種間聯結性。而鄉土物種的加入對原生植物群落產生擾動,盡管它們具有優良的植物學特性、對環境適應能力強[31-33],仍然會對原生群落的種間聯結性產生負效應。而對于群落穩定性提高這一現象,推測是由于補播處理使土壤有效養分呈現出表聚性[34],使根系可利用資源量增加;以及隨著植株生長,葉層高度及覆蓋度出現差異[34],這兩者綜合作用導致的。這有利于生物成層及資源利用效率的升高,最終使得群落穩定性增大。
綜上所述,在進行補播對植物群落穩定性的影響評價時,可將種間聯結性分析方法與群落穩定性分析方法相結合,二者所得結論能夠相互印證,并在一定程度上對群落演替方向做出指示,具有很大實踐意義。
3.2 生物量穩定性
群落生物量是群落的生長發育情況的直觀表征[23],生物量穩定性則是衡量群落穩定性大小的重要指標[24]。試驗結果表明,5種組合的群落生物量穩定性存在差異,其中組合D(扁蓿豆+星星草+垂穗披堿草+異針茅+溚草)的生物量穩定性最高,這可能是因為該組合兼具禾草及豆草,其中豆科植物根系入土深,能夠與根瘤菌共生并完成氮素固持[35];而禾本科植物根系入土淺,比表面積大,利于養分吸收[35]。這種根系分布及作用特點的差異使得組合D植物群落得以維持較高的資源分配效率及生物量穩定性,進而獲得較高的群落穩定性。
此外,研究結果還表明,群落生物量穩定性與禾本科功能群生物量穩定性之間存在極顯著的相關關系(R2=0.633,P<0.01),而與其它功能群的生物量穩定性不存在顯著的相關關系。推測可能是以下幾點原因引起的:首先,選用的7個鄉土物種中有6個為禾本科植物,試驗樣地自身包含的34種植物中禾本科便占據了10種,故試驗設計本身便體現了禾本科對于整體的重要作用;其次,基于1971—2010年的氣候監測數據[36],青藏高原在近40年里整體氣溫升高了約0.5℃,而禾本科植物對增溫十分敏感,增溫有助于其縱向生長、截取更多光照資源[37],并促進根系向下生長尋求水分[38];最后,禾本科植物自身較高[37],而高度又是地上生物量的決定因素之一。綜上所述,諸多因素共同作用,確定了禾本科生物量穩定性同群落總體生物量穩定性間極顯著的相關關系(P<0.01)。
4 結論
研究表明,在高寒草甸天然草地選用5種鄉土物種混播組合進行補播降低了群落種間聯結性、提高了群落穩定性及生物量穩定性。5種組合中,組合D(扁蓿豆+星星草+垂穗披堿草+異針茅+溚草)對群落生物量穩定性的提高最多,是生態恢復的最優組合。這可能與該混播組合為豆、禾混播,對資源的利用率高、配給合理有關。本試驗跨度僅為2年,僅能得到群落穩定性的初步變化趨勢,要想得到天然草地選用鄉土物種混播組合進行補播對草地植物群落穩定性的長期影響,還需要更長時間來探索。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
