大豆鮮莢籽粒上位性QTL及其互作效應分析
2021-09-11 08:14:40陳士亮孫亞倩邵振啟李文龍孔佑賓杜匯李喜煥張彩英
太平洋學報 2021年8期
陳士亮, 孫亞倩, 邵振啟, 李文龍, 孔佑賓,杜匯, 李喜煥*, 張彩英
(1.河北農業大學農學院, 華北作物改良與調控國家重點實驗室, 河北 保定 071001;2.河北農業大學生命科學學院, 河北 保定 071001)
大豆[Glycinemax(L.) Merr.]是重要的糧食與油料作物,籽粒富含蛋白、脂肪、異黃酮、氨基酸等多種營養物質,在人們膳食結構中占有不可替代的重要地位[1-2]。同時,大豆還可進行鮮莢采摘,作為鮮食毛豆豐富人們餐桌[3-4]。據統計,鮮食毛豆單位面積經濟效益約為普通粒用大豆的2倍以上,因而對于提高大豆經濟效益具有重要意義。然而,鮮食大豆產量與鮮莢、鮮粒性狀關系密切。大豆籽粒產量與其莢長性狀呈現極顯著正相關[4];改良大豆莢長和莢寬性狀不僅有利于增大籽粒,并且有利于提高其單位面積產量[5]。并且,鮮莢和籽粒性狀是鮮食大豆品種審定和出口的重要參考依據。因此,改良大豆鮮莢和籽粒相關性狀,對于鮮食大豆品種審定、產品出口和經濟效益提升均具有重要意義[6]。
目前,已有少數學者針對大豆莢粒相關性狀開展研究。王珍[7]利用大豆重組自交系(recombinant inbred line, RIL)獲得多個控制二粒莢長、莢寬、粒長和粒寬加性數量性狀位點(quantitative trait loci,QTL),分別位于9條染色體(4、5、6、7、8、10號染色體等)上。趙晉銘等[8]分析大豆RIL群體百粒鮮重,獲得1個位于18號染色體,可解釋7.64%~12.7%表型變異加性QTLs。牛遠[9]鑒定了257份大豆資源籽粒相關性狀,獲得多個控制粒長、粒寬、粒厚加性QTLs。Kenichiro等[10]鑒定大豆RIL群體單株莢數、每莢粒數和單粒重等,獲得4個穩定QTLs。由此可見,盡管目前已有少數關于大豆莢粒相關性狀遺傳位點研究報道[11-16],但多是利用成熟籽粒針對單一加性QTLs進行研究,很少有利用鮮莢和鮮粒、針對上位性QTLs及其互作效應開展研究的報道。
然而,上位性QTLs及其互作效應作為數量性狀遺傳結構重要組成部分,其對復雜數量性狀的影響與簡單性狀相比更為重要,因而在育種工作中的作用不容忽視[17-18]。由于自花授粉作物較異花授粉作物具有較強的非等位基因之間的互作,因而若檢測到加性×加性上位性互作效應,則可以通過人工選擇實現其穩定遺傳,進而產生額外的遺傳收益[19];并且,在上位性效應存在時,僅考慮加性效應將會嚴重影響高世代材料的選擇效率[19]。有學者在研究水稻種子活力相關性狀時發現,對于水稻種子發芽率、苗長和干物重而言,上位性效應的作用高于加性效應[20]。另外,對于異花授粉作物而言,上位性效應還是其雜種優勢形成的重要遺傳基礎。可見,在研究數量性狀單個加性QTL的同時,挖掘控制其上位性QTL及其互作效應對于今后實現數量性狀精準遺傳改良具有重要意義。
鑒于此,本研究利用大豆重組自交系群體,2年4種不同環境條件下分別鑒定其鮮莢、鮮粒共8個相關性狀,在前期已明確其在鮮莢、鮮粒等相關性狀存在廣泛遺傳變異,并已揭示其在不同環境條件下存在許多一致性加性主效QTLs基礎上,繼續挖掘控制其鮮莢籽粒性狀上位性QTLs,并分析其互作效應,對今后結合利用加性和上位性QTLs,實現大豆鮮莢籽粒性狀分子遺傳改良以及進一步解析大豆莢粒性狀分子遺傳機制具有重要意義。
1 材料與方法
1.1 供試材料
以大豆重組自交系群體(C813×墾農7號,193個家系)為材料,由河北農業大學大豆遺傳育種課題組構建并提供。
1.2 供試大豆RIL群體田間種植與管理
供試大豆重組自交系群體分別于2019、2020年種植于河北省4種環境條件下(E1~E4),以鑒定供試群體在不同年度、不同地點的遺傳變異,其中E1為2019年保定環境(河北農業大學作物育種中心)、E2為2020年保定環境(河北農業大學作物育種中心)、E3為2020年文安環境(廊坊文安)、E4為2020年鹿泉環境(石家莊鹿泉)。從地理位置看,鹿泉試點較保定試點偏西偏南、文安試點較保定試點偏東,4個試點在溫度、水分和土壤地力等方面存在一定差異。4個試點均采用完全隨機區組試驗設計,行長3 m,行距0.5 m,3次重復,田間管理同一般大田管理方法。
1.3 供試大豆RIL群體鮮莢籽粒相關性狀測定
供試大豆RIL群體生長至R6~R7期,參照國家鮮食大豆區域試驗調查方法并結合前人研究進展[7, 11],分別檢測供試群體鮮莢與籽粒相關性狀,具體包括標準二粒莢長(fresh pod length, FPL)、二粒莢寬(fresh pod width, FPW)、鮮莢重量(fresh pod weight, FPWT)、鮮粒重量(fresh seed weight, FSWT)、鮮粒長度(fresh seed length, FSL)、鮮粒寬度(fresh seed width, FSW)、長寬比(fresh seed length/width, FSLW)、鮮粒面積(fresh seed area, FSA)共8個性狀。
1.4 供試RIL群體上位性QTLs及其互作效應分析
利用供試大豆RIL群體上述8個鮮莢籽粒相關性狀,結合前期已構建的群體單核苷酸多態性(single nucleotide polymorphisms, SNP)標記遺傳連鎖圖譜(含2 234個SNP標記),采用IciMapping V4.2軟件MET模塊[21],將不同環境設為因子,通過完備區間作圖法,對8個性狀進行上位性QTLs及其互作效應分析,其中檢測步長設為5.0 cM,若臨近QTL間的遺傳距離小于5.0 cM,則將其認定為相同QTL;隨后,參照McCouch等[22]提出的QTL命名方法進行命名。
2 結果與分析
2.1 大豆鮮莢籽粒上位性QTLs及其染色體分布
通過分析供試大豆RIL群體8個鮮莢籽粒相關性狀上位性QTLs(表1)發現,共獲得321對鮮莢籽粒“加性×加性”上位性QTLs,涉及大豆所有染色體,其中以13號染色體分布的上位性QTLs數量最多,其次為6號染色體,而以1號和16號染色體分布數量最少。由此可見,控制大豆鮮莢籽粒的上位性QTLs數量較多,且在染色體上的分布范圍較廣,除以往報道的單個加性QTLs外,“加性×加性”上位性QTLs在大豆鮮莢籽粒形成和發育中也發揮著重要作用。進一步分析上述測試性狀檢測到的上位性QTLs還發現,以鮮粒長寬比定位到的上位性QTLs最多,其次為鮮粒寬度,而以鮮莢寬度定位到的上位性QTLs最少。
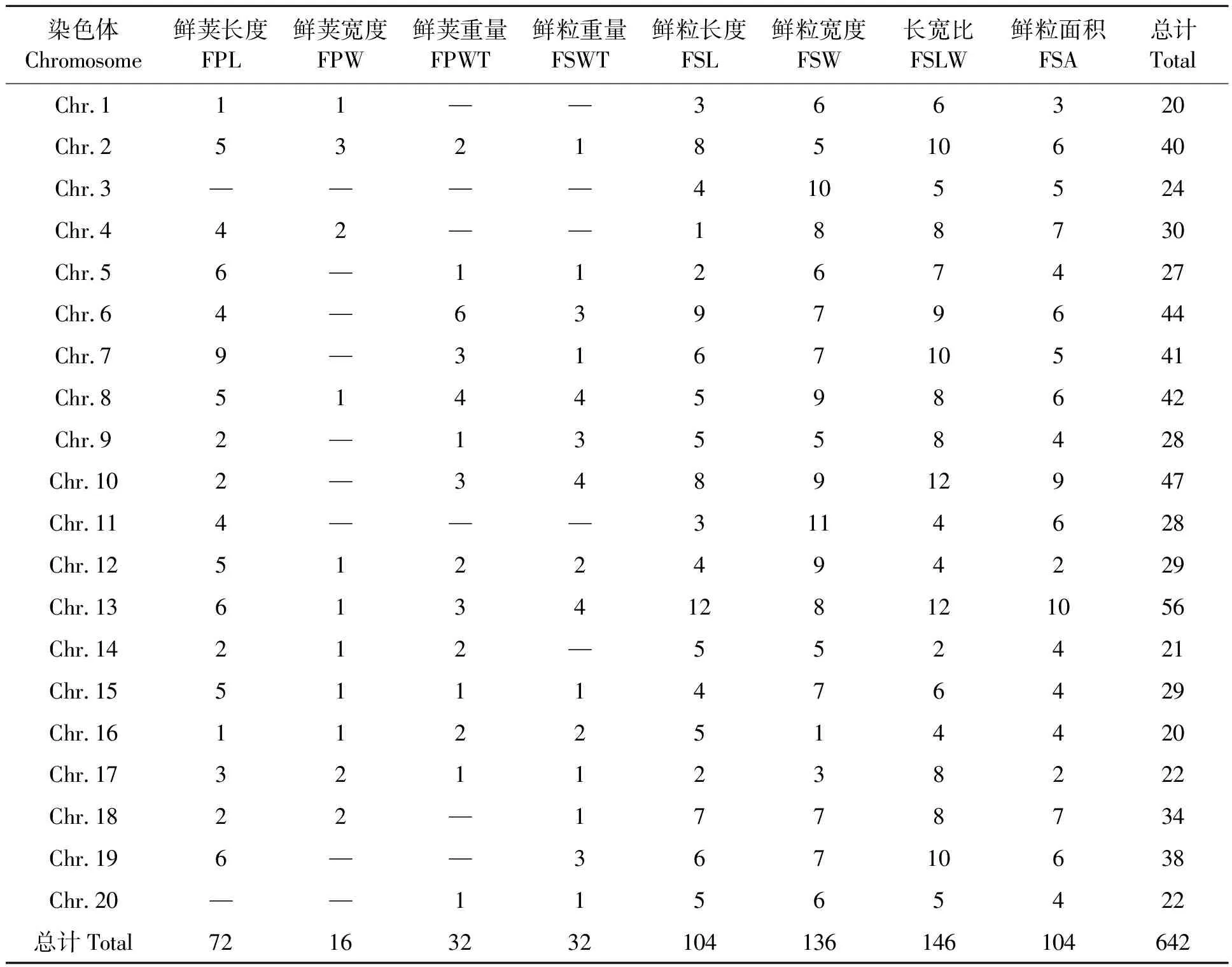
表1 大豆鮮莢籽粒上位性QTLs染色體分布Table 1 Distribution of epistatic QTLs of fresh pod and seed related traits on soybean chromosomes
另外,上述上位性QTLs互作關系如圖1所示,可以看出,大豆鮮莢籽粒性狀存在復雜的QTLs互作網絡,并以7號、13號染色體與其他染色體間的互作關系較多,其中7號染色體與其余18條染色體間存在互作關系(除3號和17號染色體外),而13號染色體則與除1號和19號染色體外的18條染色體存在互作關系。由此推測,7號和13號染色體可能在大豆鮮莢和籽粒形成與發育過程中處于較為重要的“核心”位置,其通過與較多的染色體間的互作來影響鮮莢和籽粒形成。

圖1 大豆鮮莢籽粒性狀上位性QTLs染色體互作網絡Fig.1 Interaction network of epistatic QTLs associated with soybean fresh pod and seed related traits
2.2 大豆鮮莢籽粒上位性QTLs效應分析
通過分析供試大豆群體鮮莢籽粒性狀上位性QTLs遺傳效應(表2)發現,在定位到的321對上位性QTLs中,有144對上位性QTLs為正向效應(加性效應為正值),表明兩基因座位間的互作基因型與具有正向效應的加性基因型作用方向相同;有177對上位性QTLs為負向效應(加性效應為負值),說明兩基因座位間的互作基因型與具有正向效應的加性基因型作用方向相反。在144對正向效應QTLs中,有37對QTLs效應為“++”型,70對效應為“+-”型,另有37對效應表現為“--”型;而在177對負向效應QTLs中,有45對效應為“++”型,99對效應為“+-”型,33對效應為“--”型。
同時分析不同莢粒相關性狀連鎖QTLs(表2)發現,控制鮮莢長度的36對上位性QTLs中,有19對表現為正向效應,17對表現為負向效應,其中8對為“++”型,22對為“+-”型,6對為“--”型;控制鮮莢寬度的8對上位性QTLs中,2對表現為正向效應,均為“--”型,6對表現為負向效應。控制鮮莢重量的16對QTLs中有6對表現為正向效應,10對表現為負向效應;控制鮮粒長度的52對QTL中有21對正向效應,31對負向效應;鮮粒寬度的68對QTLs中,29對表現為正向效應,39對表現為負向效應。上述上位性QTLs及其遺傳效應分析可為今后改良大豆莢粒相關性狀提供一定參考。

表2 大豆鮮莢籽粒性狀上位性QTLs遺傳效應分析Table 2 Genetic effect analysis of epistatic QTLs controlling soybean fresh pod and seed related traits
2.3 大豆鮮莢籽粒性狀“一因多效”上位性QTLs分析
2.3.1“一因多效”上位性QTLs染色體分布與效應分析 通過分析不同鮮莢籽粒性狀定位到的“加性×加性”上位性QTLs(表3)發現,檢測到34對“一因多效”上位性QTLs,其貢獻率分布在1.98%~6.58%之間,且“加性×加性”上位性QTLs間互作貢獻率為1.89%~4.85%(PVE-AA),明顯高于其與環境互作間的貢獻率(PVE-AAE),說明這些上位性QTLs主要由遺傳因素決定,而環境因素影響較小。
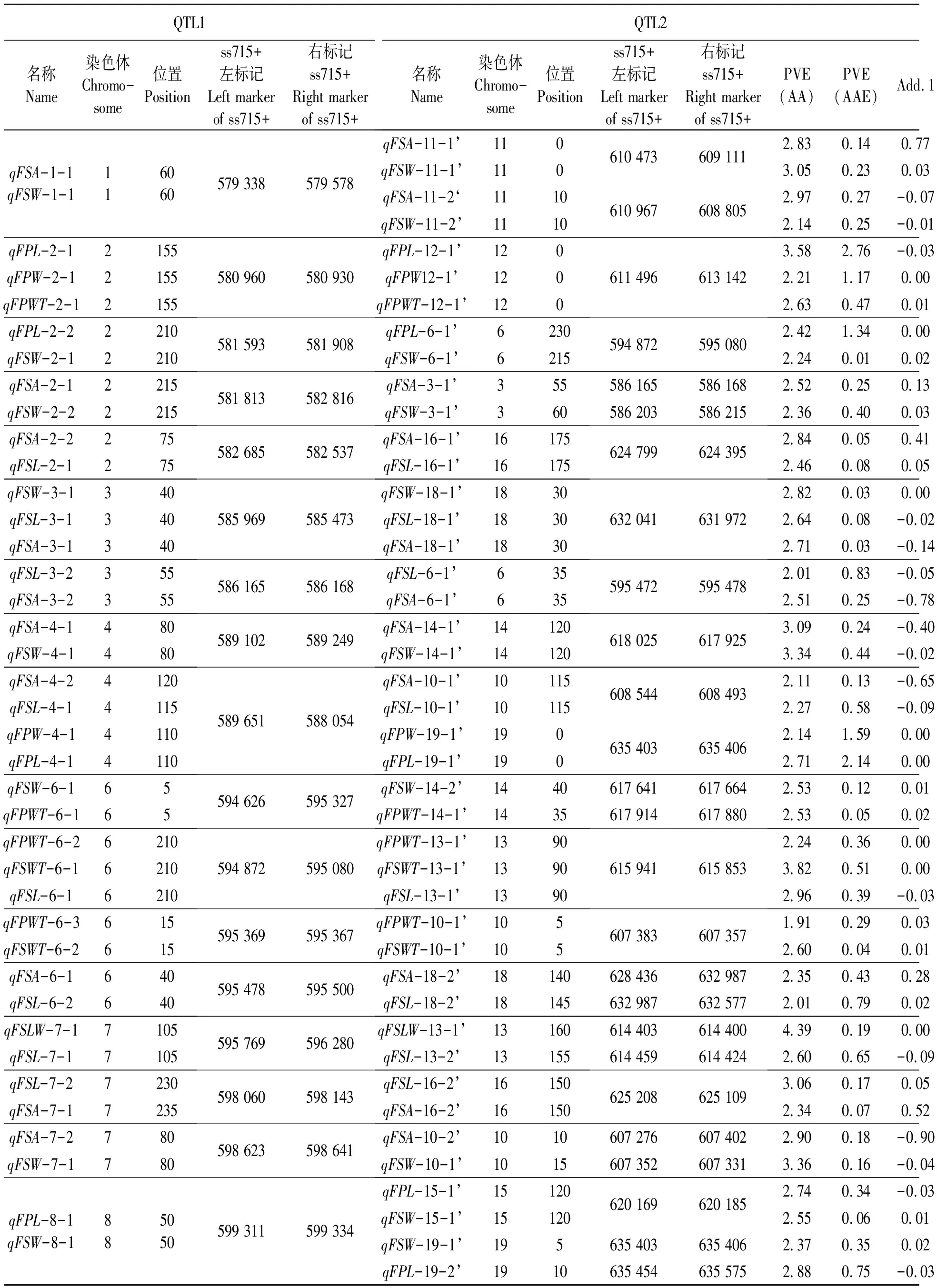
表3 大豆鮮莢籽粒性狀一因多效上位性QTLsTable 3 Pleiotropism epistatic QTLs associated with soybean fresh pod and seed related traits
同時發現,在上述34對“一因多效”QTLs中,有23對QTLs定位區間完全一致,11對QTLs定位區間距離較近(< 5 cM),涉及18條染色體,并以13號染色體數量最多,該染色體分別與6號、7號、8號、11號、12號、13號、14號、15號和16號染色體發生互作;6號染色體存在6對“一因多效”QTLs,其分別與2號、3號、10號、13號、14號和18號染色體發生互作。可見,由于大豆鮮莢籽粒相關性狀間存在一定相關性,這些相關性不僅與單個加性QTLs的一因多效有關,而且與上位性QTLs的一因多效密切相關。上述一因多效位點的發現可為今后進一步揭示大豆莢粒性狀間的相關關系提供分子依據。
另外還發現,上述34對“一因多效”上位性QTLs中,有12對QTLs效應值為正值,另有22對QTLs效應值為負值;并且,上述“一因多效”上位性QTLs在不同性狀間的效應值同時為正值或負值,一方面揭示了上述性狀間的正相關關系,同時為今后實現上述性狀同步遺傳改良提供了依據。
2.3.2“一因多效”上位性QTLs互作方式分析
在上述34對“一因多效”QTLs中,包含6組“一對多”上位性QTLs互作方式(圖2)和16組“一對一”上位性QTLs互作方式。在6組“一對多”上位性QTLs中,包括2組“一對三”上位性QTLs和4組“一對二”上位性QTLs,其中8號染色體(ss715599311~ss715599334)與15號染色體(ss715620169~ss715620185)和19號染色體2個區間(ss715635403~ss715635406、ss715635454~ss715635575)存在互作;11號染色體(ss715610473~ss715609111)與1號染色體(ss715579338~ss715579578)、15號染色體2個區間(ss715620881~ss715620795、ss715621359~ss715621034)存在互作。
同時,1號染色體(ss715579338~ss715579578)與11號染色體2個區間(ss715610473~ss715609111、ss715610967~ss715608805)存在互作;4號染色體(ss715589651~ss715588054)與10號(ss715608544~ss715608493)、19號染色體(ss715635403~ss715635406)存在互作;6號染色體(ss715594872~ss715595080)與2號(ss715581593~ss715581908)、13號染色體(ss715615941~ss715615853)存在互作;19號染色體(ss715635403~ss715635406)與4號(ss715589651~ss715588054)、18號染色體(ss715632987~ss715632577)存在互作關系。

注:*為“一對多”類型上位性QTL。Note: * indicates the “single to multiple” QTL interaction mode.圖2 大豆鮮莢籽粒性狀“一因多效”上位性QTLs互作分析Fig.2 Interaction analysis of pleiotropism epistatic QTLs associated with soybean fresh pod and seed related traits
3 討論
莢粒性狀對于鮮食大豆品種審定、產品出口以及產量和品質形成等至關重要,而且是普通粒用大豆產量性狀重要構成因素,因而已成為大豆育種重要目標性狀。大豆莢長、莢寬、莢重、籽粒長度、寬度等性狀與產量關系密切[23];并且,莢長、莢寬、莢重和鮮百粒重等性狀間存在顯著相關[24]。然而,大豆莢粒性狀屬于多基因控制數量性狀,因而發掘控制其遺傳位點成為實現性狀分子遺傳改良重要前提。
據統計,有關大豆不同莢粒相關性狀遺傳位點發掘研究最早可追溯到20世紀初,到目前為止,已報道的10余個莢粒相關性狀加性QTLs約400多個,分布范圍涉及大豆所有染色體。截至2018年,國內外已報道的154個大豆莢粒性狀加性QTLs,經元分析(meta-analysis)等方法最終獲得13個有效加性QTLs,分別位于2號、3號、4號、5號、6號、7號和14號染色體,其中5個QTLs區間縮小到0.5 Mb以內,同時在上述QTLs區間尋找到11個與莢粒性狀相關候選基因[25]。
與上述莢粒相關加性QTLs相比,目前有關莢粒性狀上位性QTLs及其互作效應分析研究報道甚少,發掘到的上位性遺傳位點數量也屈指可數。梁慧珍等[26]對大豆RIL群體6個籽粒性狀進行分析,檢測到3對影響粒寬和寬厚比的上位性QTLs。楊振等[17]利用大豆RIL群體定位到9對控制二粒莢長的加性×加性上位性QTLs,涉及1號、2號、3號、5號、6號、11號、14號、16號和20號染色體,可解釋莢長性狀9.02%表型變異,并提出上位性QTLs在大豆莢長、莢寬性狀遺傳中起著至關重要的作用。由此可見,在已獲得加性QTLs基礎上,進一步分析控制大豆莢粒性狀上位性QTLs才能更充分地揭示莢粒性狀分子遺傳基礎,也才能更準確地實現性狀分子遺傳改良。
鑒于此,本研究利用前期獲得的大豆RIL群體,在2年4種不同環境條件下鑒定其8種鮮莢籽粒相關性狀,獲得321對性狀連鎖上位性QTLs,分布于所有染色體;進一步分析發現,其中包含34對“一因多效”上位性QTLs,涉及18條染色體,且有6組“一對多”類型的上位性QTLs,說明大豆莢粒性狀除與已報道的單個加性QTLs有關外,還受多個上位性QTLs調控,且這些上位性QTLs形成了復雜的互作網絡,進一步揭示這些QTLs互作網絡是實現性狀分子遺傳改良的重要前提。
進一步分析本研究定位到的上位性QTLs可以發現,與前人報道具有一定程度的一致性。本研究在3號染色體ss715585969~ss715585473區間和18號染色體ss715632041~ss715631972區間檢測到大豆粒長、粒寬和籽粒面積“一因多效”上位性QTLs,其中3號染色體ss715585969~ss715585473區間與Cui等[27]獲得的粒長與粒寬加性QTLs以及Li等[28]獲得的莢重加性QTL重疊。同時,本研究在10號染色體和7號染色體檢測到控制籽粒面積的上位性QTLs,其中10號染色體的定位區間與梁慧珍等[26]獲得的籽粒長寬比QTL距離較近。并且,本研究在13號染色體ss715614097~ss715614125區間與ss715614050 ~ss715614188區間定位到粒長和籽粒面積的“一因多效”上位性QTLs,該QTLs與Hina等[29]定位的籽粒長寬比加性QTL重疊。
同時,本研究還發現,11號染色體ss715610795~ss715610804區間與13號染色體ss715614663~ss715614511區間存在一個粒長、粒寬和籽粒面積“一因多效”上位性QTLs,該QTL的11號染色體區間與楊振等[17]定位的莢長上位性QTLs距離較近,且與Soybase中公布的大豆粒重遺傳位點Seedweight49-2[30]一致。同時,6號染色體ss715594872~ss715595080區間與13號染色體ss715615941~ss715615853區間檢測到的莢重、粒重和粒長“一因多效”上位性QTLs,與楊振等[17]定位的莢長上位性QTL重疊,且與Soybase中公布的莢寬遺傳位點Podwallwidth1-2[31]、Podwallwidth1-3[31],粒重位點Seedweight36-2[32]、Seedweight49-4[30]和Seedweight49-5[30]重疊。這些一致性QTLs一方面驗證了本研究結果可靠性,另一方面也說明大豆莢粒加性QTLs除自身主效應外,還可通過與其他位點的互作來調控莢粒形成與發育過程。
除上述與前人報道的一致性加性或上位性位點外,本研究還發現了一些未報道的上位性遺傳位點;同時,通過對比本課題組前期已發現的大豆莢粒相關性狀加性QTLs可以發現,有很多上位性QTLs并未在加性QTLs中檢測到,比如本研究在大豆3號、4號、12號、17號和18號染色體檢測到多對上位性QTLs,而在上述染色體中并未檢測到有加性QTLs存在。由此可見,并未引起顯著加性效應的單個QTL有時可以通過與其他位點的互作來形成顯著的上位性效應,進而影響大豆莢粒相關性狀。因此,在大豆莢粒性狀分子遺傳改良中,一方面要考慮加性QTLs,另一方面還應充分考慮上位性QTLs作用。
另外,上位性QTL應用價值還體現在能夠揭示已有功能基因分子作用機制方面。艾麗娟等[33]在大豆2號染色體(qHS-2-1)和6號染色體(qHS-6-1)之間檢測到一對控制籽粒硬實現象的上位性QTL,通過查閱文獻得知,2號染色體存在2個控制種皮滲透性的相關基因,而關于這2個基因的分子作用機制尚不清楚,因而認為可以將這2個功能基因作為出發點,通過尋找與其相關的調控因子或酶類,進而確定6號染色體qHS-6-1區間與其互作的基因,實現解析上述功能基因分子作用機制的目的。因此,上位性QTL的挖掘還可為揭示已有功能基因分子作用機制提供參考。
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
中學物理·高中(2016年12期)2017-04-22 11:53:03
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
