柱花草磷饑餓響應基因SgPHR1和SgPHR2的克隆與表達分析
2021-09-14 02:37:23黃杰宋劍靈安娜鄒曉燕李季膚劉國道陳志堅
熱帶作物學報 2021年8期
黃杰 宋劍靈 安娜 鄒曉燕 李季膚 劉國道 陳志堅


摘 ?要:低磷脅迫是限制作物生長和產量的重要因素之一。磷饑餓響應因子PHR(phosphate starvation response)是植物磷信號調控網絡中的關鍵因子,具有調控植物磷平衡的生物學功能。本研究在柱花草(Stylosanthes guianensis)中克隆到轉錄因子SgPHR1和SgPHR2基因。SgPHR1和SgPHR2基因cDNA全長分別為1413?bp和849?bp,編碼470和282個氨基酸殘基,蛋白分子量分別為51.4?kD和30.9?kD。SgPHR1和SgPHR2均為SANT家族成員,包含MYB蛋白結構域和CC蛋白結構域,并具有多個潛在的磷酸化位點。亞細胞定位預測表明,SgPHR1和SgPHR2均定位于細胞核中。實時定量PCR結果表明,SgPHR1基因在柱花草根和葉中的表達量高于莖中的表達量,而SgPHR2基因在葉中表達量顯著高于根和莖。缺磷(-P)和缺氮(-N)處理均顯著增強了SgPHR1和SgPHR2在柱花草根中的表達。不同缺磷時間處理結果進一步表明了SgPHR1和SgPHR2在轉錄水平上響應低磷脅迫,暗示SgPHR1和SgPHR2可能參與了柱花草對低磷脅迫的應答。本研究結果為解析柱花草響應低磷脅迫的分子機制提供了候選基因。
關鍵詞:低磷脅迫;柱花草;PHR;基因表達;轉錄因子
Abstract: Low phosphorus (P) stress is one of the constraints limiting crop growth and yield. PHR (phosphate starvation response) is the key factor in P signaling network, regulating plant phosphate (Pi) homeostasis. In this study, two transcription factors, SgPHR1 and SgPHR2, were cloned in stylo (Stylosanthes guianensis). The full length of SgPHR1 and SgPHR2 was 1413?bp and 849?bp, encoding 470 and 282 amino acid residues, respectively. The protein molecular weight of SgPHR1 and SgPHR2 was 51.4?kD and 30.9?kD, respectively. Both SgPHR1 and SgPHR2 belonged to the SANT family and contained MYB protein domain and CC protein domain. Both SgPHR1 and SgPHR2 included a set of potential phosphorylation sites. Subcellular localization prediction indicated that both SgPHR1 and SgPHR2 localized to the nucleus. Real-time quantitative PCR results showed that the transcript of SgPHR1 in leaves and roots was higher than that in stems, while SgPHR2 exhibited the highest expression in leaves. Furthermore, the expression of SgPHR1 and SgPHR2 was up-regulated by both -P and nitrogen (-N) deficient treatments in stylo roots. The transcription level of SgPHR1 and SgPHR2 was found to be increased during -P treatments, suggesting that SgPHR1 and SgPHR2 involved in stylo response to Pi starvation. This study would provide candidate genes for investigating the mechanism of stylo response to P deficiency.
Keywords: low phosphorus stress; Stylosanthes; phosphate starvation response; gene expression; transcription factor
磷(phosphorus,P)是植物生長發育所必需的大量營養元素之一,參與植物多種生理生化過程,如光合作用、能量和脂類代謝等[1-3]。然而,土壤磷的有效性低,特別是在酸性土壤中,磷容易被鋁(Al)和鐵(Fe)等金屬元素結合形成不能被植物直接吸收和利用的難溶性磷[4-5]。因此,低磷脅迫是酸性土壤上限制作物生長和產量的重要因素之一[6]。在低磷環境下,植物表現出植株矮小,生長緩慢,葉色暗綠和根系發育受阻等脅迫癥狀[7]。為應對低磷脅迫,植物形成了一系列的適應性機制,如植物可以通過改變根系形態和構型增加對土壤磷的吸收[8-9];通過分泌有機酸和酸性磷酸酶來活化利用土壤難溶性磷和有機磷[10-12];通過誘導磷轉運基因表達增強對磷的吸收和轉運[13]。另外,磷信號網絡在維持植物細胞磷平衡中具有重要調控作用[14-15]。
磷饑餓響應因子(phosphate starvation response,PHR)屬于MYB-CC型轉錄因子,在磷信號網絡中發揮關鍵作用[16]。PHR具有Myb DNA結合結構域,C端具有卷曲螺旋(CC)結構域。PHR一般定位于細胞核,具有轉錄調控的作用,能調控下游磷饑餓響應基因的表達,如PT、PHT和MIR399等[17]。在低磷條件下,PHR會形成二聚體,通過CC結構域結合在磷饑餓響應基因啟動子P1BS(GNATATNC)順式作用元件上,從而調控靶基因表達[16]。綠色衣藻(Algachamycary)CrPSR1是最早發現的MYB-CC型轉錄因子,其在調節綠色衣藻響應低磷脅迫中起作用[17]。AtPHR1是擬南芥(Arabidopsis thaliana)磷信號網絡中的重要調節因子,其可通過調控PT等下游基因表達響應低磷環境[18-19]。近年來,在水稻(Oryza sativa)[20]、玉米(Zea mays)[21]、大豆(Glycine max)[22]和菜豆(Phaseolus vulgaris)[23]等植物中相繼克隆并鑒定了PHR同源基因,暗示PHR在植物適應低磷脅迫中起重要作用。
柱花草(Stylosanthes spp.)原產于美洲和非洲熱帶和亞熱帶地區,是重要的熱帶豆科牧草[24]。柱花草廣泛用于牧草飼料和綠肥覆蓋等方面。柱花草具有良好的適應低磷酸性土壤的能力,其被認為是研究植物適應酸性土壤低磷脅迫的重要熱帶豆科植物。迄今為止,已從柱花草中克隆了一系列低磷響應基因,如SgPT1、SgPAP7/10/23/26和SgSPX1[25-27],但是,對柱花草磷信號網絡調控因子基因的克隆和研究較少。因此,本研究克隆了柱花草2個磷饑餓響應轉錄因子基因SgPHR1和SgPHR2,并對這兩個基因進行了生物信息學和基因表達分析,可為進一步解析柱花草適應低磷脅迫的分子調控網絡提供基因資源。
1 ?材料與方法
1.1 ?材料
本研究所用的實驗材料為‘熱研2號圭亞那柱花草(Stylosanthes guianensis)。柱花草種子由中國熱帶農業科學院熱帶作物品種資源研究所草業研究中心提供。
1.2 ?方法
1.2.1 ?材料與處理 ?柱花草種子去種皮后于80?℃水浴3?min,在濕潤的濾紙上萌發2?d。將萌發后的柱花草幼苗移到1/2 Hoagland營養液中進行正常培養。柱花草培養14?d后,進行不同磷濃度處理,分別為0.5?μmol/L KH2PO4(低磷,LP)和250?μmol/L KH2PO4(正常磷,CK)。處理14?d后收獲樣品,測定生物量和磷含量。每個處理設置3個生物學重復。
1.2.2 ?磷含量測定 ?柱花草樣品置于105?℃殺青30?min,75?℃烘干至恒重。將樣品粉碎后,稱取0.07?g樣品,在馬弗爐中600?℃灰化10?h,然后采用鉬銻抗顯色法測定植株磷含量。
1.2.3 ?RNA提取和cDNA合成 ?總RNA提取參考TRIzol Universal提取試劑(Tiangen,中國)說明書。稱取0.1?g根系樣品,加入1?mL TRIzol提取液充分研磨并在室溫放置5?min。4?℃,12 000?r/min離心3?min,吸取上清液至1.5?mL離心管中,加入預冷的5?mol/L氯化鈉和氯仿,充分混勻。4?℃,12 000?r/min離心10?min,取上清液,加入等體積的異丙醇,充分混勻,室溫放置10?min。4?℃,12 000?r/min離心10?min,棄上清液,加入1?mL 75%乙醇。4?℃,7500?r/min離心5?min,棄上清液,風干沉淀,加入無RNase ddH2O充分溶解RNA。
參考RevertAid First Strand cDNA Synthesis試劑盒(Thermo Fisher,美國)方法合成cDNA第一鏈。在PCR管中加入2?μg RNA、1?μL Oligo (dT)18,并加入無RNase ddH2O至12?μL。于65?℃反應5?min后,加入4?μL 5×Reaction Buffer、2?μL dNTP Mix、1?μL RNase Inhibitor和1?μL RevertAid M-MuL V Reverse transcriptase。反應混合物于42?℃反應60?min,70?℃反應5?min。反應結束后,cDNA樣品于–20?℃保存備用。
1.2.4 ?SgPHR1和SgPHR2全長克隆 ?基于本課題組柱花草根系低磷響應轉錄組結果,篩選獲得SgPHR1和SgPHR2基因全長序列,根據序列設計SgPHR1-ORF-F/R和SgPHR2-ORF-F/R引物(表1)。以根系cDNA為模板,通過PCR擴增SgPHR1和SgPHR2基因。回收PCR產物并連接到克隆載體pMD18-T(TaKaRa,日本),進一步轉化大腸桿菌DH5α并進行測序分析,獲得SgPHR1和SgPHR2全長序列。
1.2.5 ?生物信息學分析 ?利用KinasePhos(http://kinasephos.mbc.nctu.edu.tw/)進行磷酸化位點預測;利用WoLF PSORT(https://www.genscript. com/ wolf-psort.html)進行亞細胞定位預測分析;運用ClustalX1.8進行多序列比對分析;采用MAGA-X構建系統進化樹;利用NCBI(https://www.ncbi.nlm.nih.gov/)網站進行保守結構域分析。
1.2.6 ?實時定量PCR分析 ?使用QuantStudioTM Real-Time PCR(Thermo Fisher,美國)系統和SYBR Green(Vazyme,中國)定量試劑盒,進行實時定量PCR分析。定量PCR反應體系為10?μL 2×SYBR Green PCR master mix、1?μL引物(10?μmol/L)、2?μL稀釋30倍的cDNA模板,并加入ddH2O至20?μL。反應程序為:95?℃ 1?min,95?℃ 15?s,58?℃ 15?s,72?℃ 30?s,40個循環。基因相對表達量=目的基因(SgPHRs)表達量/內參基因(SgEF1a)表達量。定量PCR引物見表1。
1.3 ?數據處理
數據通過Microsoft Excel 2010軟件進行平均數、標準誤計算和作圖,采用SPSS數據分析軟件(V23.0,SPSS Institute,美國)進行方差分析。
2 ?結果與分析
2.1 ?不同磷濃度處理對柱花草干重和磷含量的影響
從圖1A可以看出,相對正常磷(250 μmol/L KH2PO4,CK)處理,低磷(0.5 μmol/L KH2PO4,LP)處理顯著抑制了柱花草地上部干重,但對根部干重沒有顯著影響。低磷處理下的地上部干重僅為正常磷條件下的46%(圖1A)。相對正常磷(CK)處理,低磷(LP)處理顯著降低了柱花草地上部和根部磷含量,低磷處理下的地上部和根部磷含量分別僅為正常磷條件下的17%和15%(圖1B)。
2.2 ?柱花草SgPHR1和SgPHR2的克隆
研究基于前期柱花草轉錄組測序結果獲得的SgPHR1和SgPHR2基因序列,設計PCR引物,以低磷處理下柱花草根系cDNA為模板,擴增出SgPHR1和SgPHR2基因cDNA全長。如圖2所示,SgPHR1和SgPHR2基因全長分別為1413?bp和849?bp。利用ExPASyProt軟件分析發現SgPHR1基因編碼470個氨基酸殘基,蛋白分子量為51.4?kD,理論等電點為5.4,為親水性蛋白。SgPHR2基因編碼282個氨基酸殘基,蛋白分子量為30.9?kD,理論等電點為7.7,也為親水性蛋白。
2.3 ?柱花草SgPHR1和SgPHR2蛋白理化性質分析
利用WoLF PSORT進行亞細胞定位預測,結果表明SgPHR1和SgPHR2蛋白均定位于細胞核上,暗示其為核蛋白。利用KinasePhos網站對SgPHR1和SgPHR2蛋白的磷酸化位點進行預測,發現SgPHR1蛋白具有14個絲氨酸、4個蘇氨酸和4個酪氨酸等潛在的磷酸化位點,而SgPHR2蛋白具有11個絲氨酸、6個蘇氨酸和1個酪氨酸等潛在的磷酸化位點(表2)。此外,SgPHR1蛋白的磷酸化位點主要分布在N端,而SgPHR2蛋白的磷酸化位點主要分布在C端(圖3)。
2.4 ?柱花草SgPHR1和SgPHR2蛋白保守結構域和多序列比對分析
CDD Tools預測表明,SgPHR1和SgPHR2都屬于SANT家族成員,均含有PHR蛋白的保守序列,即myb_SHAQKYF和Myb_CC_LHEQLE特征序列(圖4)。SgPHR1蛋白在248~300和331~378氨基酸范圍內,分別包含myb_SHAQKYF結構域和Myb_CC_LHEQLE結構域;SgPHR2蛋白在20~76和106~152氨基酸范圍內,分別包含myb_ SHAQKYF結構域和Myb_CC_LHEQLE結構域。
多序列比對分析發現,SgPHR1和SgPHR2蛋白與已報道的不同物種PHR蛋白均包含myb_SHAQKYF和Myb_CC_LHEQLE保守結構域。此外,不同物種PHRs蛋白N端和C端同源性較低(圖5)。
2.5 ?系統進化樹分析
系統進化樹分析表明,PHRs蛋白可以分為3大組(圖6)。第Ⅰ組包括擬南芥、截形苜蓿(Medicago truncatula)、草莓(Fragaria vesca)和甘藍型油菜(Brassica naus)等植物PHR蛋白,如擬南芥AtPHR1和甘藍型油菜BnPHR1。柱花草SgPHR1被分在第Ⅰ組中,與截形苜蓿MtPHR1屬于同一分枝,同源性最高。第II組包括水稻、玉米和小麥(Triticum aestivum)等植物PHR蛋白,如水稻OsPHR1、OsPHR2、小麥TaPHR1和玉米ZmPHR1。第III組包括水稻、大豆、菜豆和柱花草等植物PHR蛋白。柱花草SgPHR2與菜豆PvPHR1、大豆GmPHR1和GmPHR25同源性最高(圖6)。
蛋白序列編號包括:柱花草SgPHR1、擬南芥(AtPHR1,NP_194590)、玉米(ZmPHR1,JF831533.1)、水稻(OsPHR1,AK063486.1)、草莓(FvPHR1,XP_004289982.1)、甘藍型(BnPHR1,JN806156.1)和大豆(GmPHR1,HQ007311);紅色方框內為MYB結構域,藍色方框內為CC結構域。
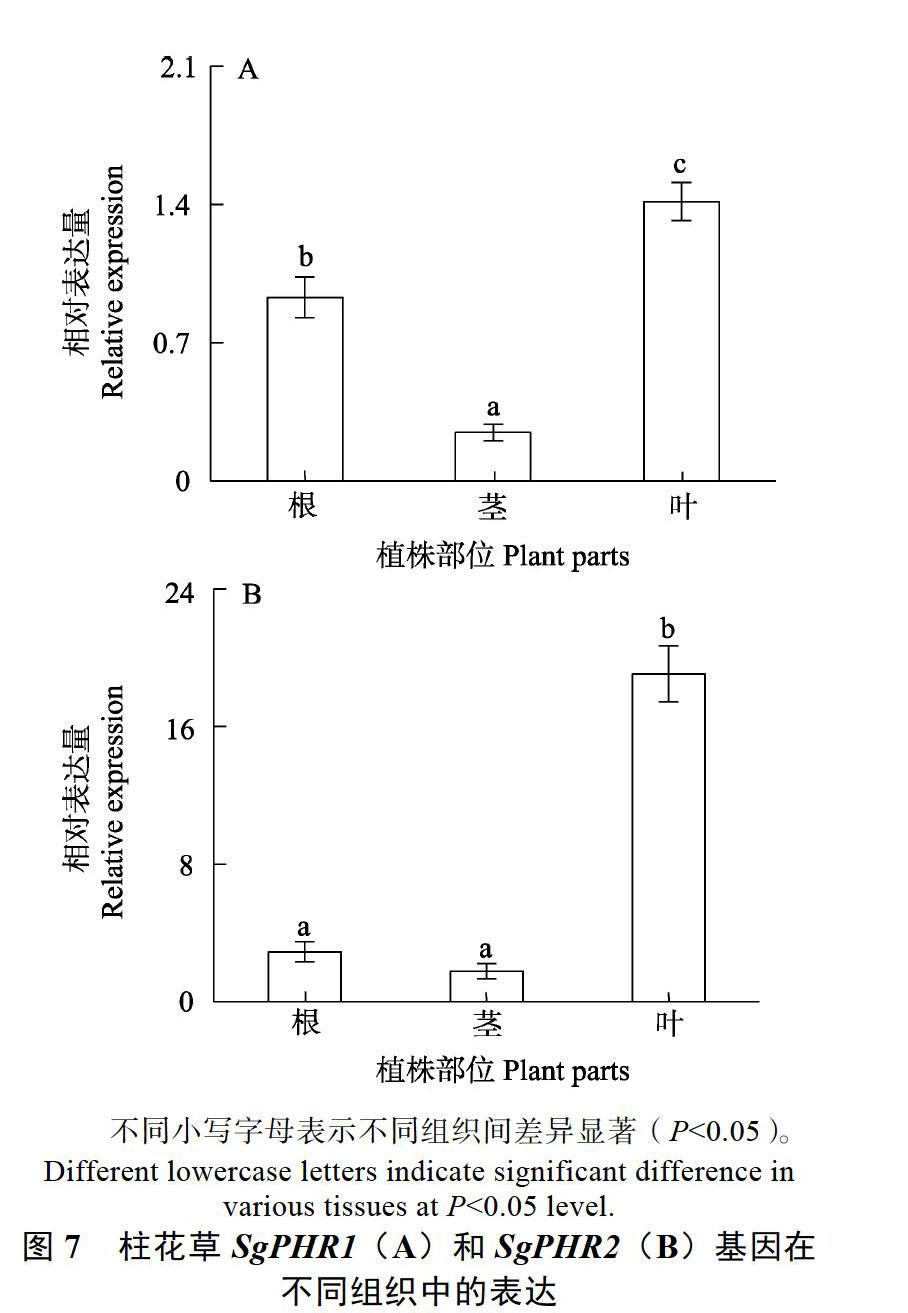
2.6 ?柱花草SgPHR1和SgPHR2基因表達分析
本研究對SgPHR1和SgPHR2基因表達進行分析。結果表明,SgPHR1和SgPHR2基因在根、莖和葉中均有表達,其中,SgPHR1在葉中表達量最高,根次之,莖中表達量最低,SgPHR2基因在葉中表達量顯著高于莖和根(圖7)。由于PHR基因被報道參與了植物對養分脅迫的應答,本研究進一步分析了不同缺素處理對柱花草SgPHR1和SgPHR2基因表達的影響。結果發現,與對照(CK)相比,缺氮(–N)、缺磷(–P)和缺鉀(–K)處理均顯著增強了SgPHR1基因在柱花草根系中的表達(圖8A);與對照相比,缺氮和缺磷處理顯著增強了SgPHR2基因在柱花草根系中的表達,而缺鉀處理對SgPHR2基因的表達無顯著影響(圖8A)。其中,缺磷處理下SgPHR1和SgPHR2基因表達量分別是對照條件下的1.3和2.6倍(圖8A)。
從圖8B可見,隨著低磷處理時間的延長,SgPHR1和SgPHR2基因表達量逐漸增加,并在低磷處理第7天基因表達量最高,SgPHR1和SgPHR2Genebank序列號包括:擬南芥(AtPHR1,NP_194590)、玉米(ZmPHR1,JF831533.1)、大豆(sGmPHR25,NP_001350613.1)、菜豆(PvPHR1,ACD13206.1)、水稻(OsPHR1,AK063486.1;OsPHR2,AK100065.1;OsPHR3,A2X0Q0.1;OsPHR4,XP_015644151.1)、小麥(TaPHR1,AGH13375.1)、草莓(FvPHR1,XP_004289982.1)、甘藍型油菜(BnPHR1,JN806156.1)、梅花(PmPHR1,XP_016650539.1)、桃李(PpPHR1,XP_020425868.1)、甜櫻桃(PaPHR1,XP_021831311.1)、海棠(MdPHR1,XP_008372382.1)、梨(PbPHR1,XP_018506029.1)、截形苜蓿(MtPHR1,XP_003625354.1)、可可樹(HcPHR1,XP_007050189.2)、巴西橡膠樹(HbPHR1,XP_021670552.1)、葡萄(VvPHR1,XP_002270511.1)、蘿卜(RsPHR1,XP_018469653.1)和大豆(GmPHR1,HQ007311)等。代表柱花草PHRs蛋白。
3 ?討論
磷是維持植物正常生長發育所必需的大量營養元素之一,低磷脅迫抑制作物的生長和產量。在水稻、大豆和玉米等植物中的研究發現,低磷脅迫顯著抑制了這些植物的植株生物量和磷含量[20-22]。類似的,本研究結果發現,低磷處理顯著抑制了柱花草地上部干重,并降低了柱花草地上部和根部磷含量(圖1)。在應對低磷脅迫過程中,植物形成了如改變根系形態構建型等適應性機制,進而增強對土壤磷的吸收和利用[8]。低磷脅迫下植物的適應性變化受到磷信號網絡的精細調控,其中,轉錄因子PHR基因在植物磷信號網絡中發揮關鍵作用[15]。因此,為探索柱花草響應低磷脅迫的磷信號網絡,本研究克隆了柱花草MYB-CC型轉錄因子SgPHR1和SgPHR2(圖2)。
近年來,對MYB型轉錄因子PHR功能研究較為廣泛,PHR在磷信號網路中起重要作用[14]。
據報道,擬南芥包括14個AtPHRs、大豆包括35個GmPHRs、水稻包括4個OsPHRs[19,22,28]。PHR轉錄因子具有高度保守的與DNA結合的MYB結構域(myb_SHAQKYF)和與二聚體形成相關的螺旋CC結構域(Myb_CC_LHEQLE)。PHR通過與下游磷饑餓誘導基因的啟動子結合以調控細胞內的磷平衡。研究表明,MYB結構域和卷曲螺旋CC結構域都是PHR結合下游靶基因所必需的結構域[29]。柱花草SgPHR1和SgPHR2均屬于SANT蛋白家族成員,包含myb_SHAQKYF和Myb_ CC_LHEQLE特征序列(圖4),表明柱花草SgPHR1和SgPHR2具備典型的PHR蛋白特性和結構特征。
系統進化樹分析表明,柱花草SgPHR1被分在第Ⅰ組中(圖6),這一組包括擬南芥AtPHR1、油菜BnPHR1和草莓FvPHR1等蛋白,這些蛋白被認為是植物響應低磷脅迫的重要因子[28,30]。如,AtPHR1定位在細胞核內,是擬南芥關鍵的磷信號調控因子[18]。BnPHR1是甘藍型油菜響應低磷的重要調控因子,具有調控下游磷饑餓響應基因表達,從而促進植物對磷的吸收和維持磷平衡的生物學功能[31]。草莓FvPHR1定位于細胞核內,FvPHR1被報道可以直接激活下游磷饑餓響應基因的表達[30]。柱花草SgPHR2與大豆GmPHR1和GmPHR25以及菜豆PvPHR1等被分在第III組中。大豆GmPHR25定位在細胞核中,具有調控大豆磷穩態的生物學功能[22]。PvPHR1被報道是調控菜豆磷轉運和穩態相關基因的正調節因子[23]。因此,SgPHR1和SgPHR2可能具有植物PHR蛋白保守的生物學功能。
氮、磷和鉀是植物正常生長和發育所必需的大量營養元素。研究表明,MYB轉錄因子家族基因的表達受到氮、磷和鉀等元素缺乏的影響[14,32,33]。如,在水稻中,缺磷處理能夠增強水稻根中MYB家族基因OsPHR4的表達[31]。在大豆中,缺氮處理增強了7個GmPHR家族基因在根中的表達[22]。另外,轉錄組分析結果表明,缺鉀處理也能調控水稻和番茄(Lycopersicon esculentum)多個MYB家族基因的表達[34-35]。本研究結果發現,缺磷和缺氮處理均顯著增強了SgPHR1和SgPHR2在柱花草根系中的表達,表明SgPHR1和SgPHR2可能受到氮磷協同調控,進而參與了柱花草對氮和磷脅迫的應答。另外,SgPHR1和SgPHR2表達量隨著磷饑餓處理時間的延長而增加(圖8)。類似的,缺磷處理增加了OsPHR3和OsPHR4在水稻根部中的表達量[31],而玉米ZmPHR1在根和葉中的表達也會隨著磷饑餓程度增加而增加[21]。然而,研究發現,擬南芥AtPHR1、水稻OsPHR1和OsPHR2等PHR基因在轉錄水平上不受外源磷的影響,這些PHRs基因可能通過翻譯后水平調控植物響應低磷脅迫[19,32]。雖然低磷處理對擬南芥AtPHR1基因表達的影響不明顯,但是,與野生型擬南芥相比,phr1突變體缺失對磷饑餓響應基因的調控,表明AtPHR1在擬南芥磷信號網路中起重要的轉錄調控作用[18-19]。因此,SgPHR1和SgPHR2可能作為重要的轉錄調控因子參與了柱花草對低磷脅迫的響應。
參考文獻
Chen S S, Luo Y, Ding G D, et al. Comparative analysis of Brassica napus plasma membrane proteins under phosphorus deficiency using label--free and MaxQuant-based proteomics approaches[J]. Journal of Proteomics, 2016, 133: 144-152.
Liu Y, Xie Y R, Wang H, et al. Light and ethylene coordinately regulate the phosphate starvation response through transcriptional regulation of phosphate starvation response1[J]. The Plant Cell, 2017, 29(9): 2269-2284.
Murakawa M, Ohta H, Shimojima M, et al. Lipid remodeling under acidic conditions and its interplay with low Pi stress in Arabidopsis[J]. Plant Molecular Biology Reporter, 2019, 101(1/2): 81-93.
Yang Z B, Rao I M, Horst W J, et al. Interaction of aluminium and drought stress on root growth and crop yield on acid soils[J]. Plant and Soil, 2013, 372(1-2): 3-25.
Qu X J, Zhou J Q, Masabni J, et al. Phosphorus relieves aluminum toxicity in oil tea seedlings by regulating the metabolic profiling in the roots[J]. Plant Physiology and Biochemistry, 2020, 152: 12-22.
Koopmans G F, Chardon W J, Willigen P, et al. Phosphorus desorption dynamics in soil and the link to a dynamic concept of bioavailability[J]. Journal of Environmental Quality, 2004, 33(4): 1393-1402.
Tania?Galindo‐Castaeda, Kathleen?M Brown, Jonathan?P Lynch, et al. Reduced root cortical burden improves growth and grain yield under low phosphorus availability in maize[J]. Plant Cell and Environment, 2018, 41(7): 1579-1592.
Güsewell?Sabine.?Regulation of dauciform root formation and root phosphatase activities of sedges (Carex) by nitrogen and phosphorus[J].?Plant & Soil,?2017,?415(1-2):?57-72.
馬若囡, 劉 ?慶, 李 ?歡, 等. 缺磷脅迫對甘薯前期根系發育及養分吸收的影響[J]. 華北農學報, 2017, 32(5): 171-176.
Li C C, Li C F, Zhang H Y, et al. The purple acid phosphatase GmPAP21 enhances internal phosphorus utilization and possibly plays a role in symbiosis with rhizobia in soybean[J]. Physiologia Plantarum, 2017, 159(2): 215-227.
Mehdi Y H, Ali I D, Ali M M, et al. Agrobacterium rhizogenes transformed soybeans with AtPAP18 gene show enhanced phosphorus uptake and biomass production[J]. Biotechnology and Biotechnological Equipment, 2018, 32(4): 865-873.
Chen Z C, Liao H. Organic acid anions: An effective defensive weapon for plants against aluminum toxicity and phosphorus deficiency in acidic soils[J]. Journal of Genetics and Genomics, 2016, 43(11): 631-638.
Chen G H, Yan W, Yang S P, et al. Overexpression of rice phosphate transporter gene OsPT2 enhances tolerance to low phosphorus stress in soybean[J]. Journal of Agricultural Science and Technology, 2015, 17(2): 469-482.
Amit Sharma, Alice Mühlroth, Juliette Jouhet, et al. The Myb-like transcription factor Phosphorus Starvation Response (PtPSR) controls conditional P acquisition and remodeling in marine microalgae[J]. New Phytologist, 2020, 225(6): 2380-2395.
Xuan L T H, Du N H N, Nguyen B A T, et al. Transcription factors and their roles in signal transduction in plants under abiotic stresses[J]. Current Genomics, 2017, 18(6): 483-497.
Stanislas Thiriet-Rupert, Gregory Carrier, Beno?t Chénais, et al. Transcription factors in microalgae: genome-wide prediction and comparative analysis[J]. BMC Genomics, 2016, 17(1): 1-16.
Wykoff D D, Grossman A R, Weeks D P, et al. Psr1, a nuclear localized protein that regulates phosphorus metabolism in Chlamydomonas[J]. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(26): 15336-15341.
Rubio V, Linhares F, Solano R, et al. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae[J]. Genes and Development, 2001, 15(16): 2122-2133.
Nilsson L, Müller R, Nielsen T H, et al. Increased expression of the MYB-related transcription factor, PHR1, leads to enhanced phosphate uptake in Arabidopsis thaliana[J]. Plant Cell and Environment, 2007, 30(12): 1499-1512.
Guo M, Ruan W Y, Li C Y, et al. Integrative comparison of the role of the Phosphate Response1 subfamily in phosphate signaling and homeostasis in rice[J]. Plant Physiology, 2015, 168(4): 1762-1776.
Wang X H, Bai J R, Liu H M, et al. Overexpression of a maize transcription factor ZmPHR1 improves shoot inorganic phosphate content and growth of Arabidopsis under low-phosphate conditions[J]. Plant Molecular Biology Reporter, 2013, 31(3): 665-677.
Xue Y B, Xiao B X, Zhu S N, et al. GmPHR25, a GmPHR member up-regulated by phosphate starvation, controls phosphate homeostasis in soybean[J]. Journal of Experimental Botany, 2017, 68(17): 4951-4967.
Valdés L O, Arenas H C, Ramírez M, et al. Essential role of MYB transcription factor: PvPHR1 and microRNA: PvmiR399 in phosphorus-deficiency signalling in common bean roots[J]. Plant Cell and Environment, 2008, 31(12): 1834-1843.
嚴琳玲, 張 ?瑜, 白昌軍. 20份柱花草營養成分分析與評價[J]. 湖北農業科學, 2016(1): 128-133.
孫麗莉, 陳志堅, 劉攀道, 等. 柱花草磷轉運蛋白SgPT1的克隆和表達分析[J]. 草業學報2013, 22(4): 187-196.
劉攀道, 董榮書, 丁西朋, 等. 不同磷效率柱花草基因型對外源DNA活化利用能力的比較分析[J]. 分子植物育種, 2018, 16(4): 1085-1091.
孫麗莉, 田 ?江, 陳志堅, 等. 柱花草SgSPX1基因的克隆與表達分析[J]. 熱帶作物學報, 2012, 33(10): 1794-1799.
Ren F, Guo Q Q, Chang L L, et al. Brassica napus PHR1 gene encoding a MYB-like protein functions in response to phosphate starvation[J]. PloS One, 2012, 7(8): e44005.
Jiang?M Q, Sun?L F, Isupov?M?N, et?al. Structural basis for the target DNA recognition and binding by the MYB domain of phosphate starvation response 1[J]. The Febs Journal, 2019, 286(14): 2809-2821.
Wang Y, Zhang F, Cui W X, et al. The FvPHR1 transcription factor control phosphate homeostasis by transcriptionally regulating miR399a in woodland strawberry[J]. Plant Science, 2019, 280: 258-268.
Ruan W Y, Guo M N, Wu P, et al. Phosphate starvation induced OsPHR4 mediates Pi-signaling and homeostasis in rice[J]. Plant Molecular Biology Reporter, 2017, 93(3): 327-340.
Tiwari J K, Buckseth T, Zinta R, et al. Transcriptome analysis of potato shoots, roots and stolons under nitrogen stress[J]. Scientific Reports, 2020,?10(1):?921-941.
Du X Q, Wang F L, Li H, et al. The transcription factor MYB59 regulates K/NO translocation in the Arabidopsis response to low K stress[J]. The Plant Cell, 2019, 31(3): 699-714.
Zhang X Q, Jiang H, Wang H, et al. Transcriptome analysis of rice seedling roots in response to potassium deficiency[J]. Scientific Reports, 2017, 7(1): 5523.
Zhao X M, Liu Y, Liu X, et al. Comparative transcriptome profiling of two tomato genotypes in response to potassium-deficiency stress[J]. International Journal of Molecular Sciences, 2018, 19(8): 2402-2426.
責任編輯:沈德發
