降水量和生長方式對紅砂和珍珠根系形態特征的影響研究
2021-09-17 10:36:20單立山張正中解婷婷
草地學報 2021年5期
關鍵詞:植物
楊 潔,單立山,蘇 銘,張正中,解婷婷,李 毅
(甘肅農業大學林學院,甘肅 蘭州 730070)
大氣環流模型預測顯示,未來100年內,全球總降水量會產生巨大變化,極端天氣和干旱強降水事件的發生頻率將增加,季節性降水也會發生非常明顯的改變,總體表現為夏季單次降水量加大、降水時間延長[1]。西北干旱區降水的季節性波動表現更加明顯,廣泛分布于我國干旱和半干旱地區的荒漠生態系統由于自身的脆弱性和不穩定性,對極端干旱等氣候條件的響應更為敏感[1-3]。水分作為荒漠生態系統中植被生長的主要限制因子,其變化會直接影響植物根系的形態特征及分布[4-5],使植物不斷調整自身根系形態以適應環境水分變化。因此,加強降水變化對荒漠植物根系形態變化特征的影響的研究,對干旱區荒漠植被的保護、恢復和重建等工作具有重要意義。
根系作為最先感應到水分存在及變化的器官[6],水分不足或過量都會直接影響植物根系的形態特征、數量和空間分布,使根系各項生理生化功能受到阻礙并且生長異常[7]。良好的根系形態構型不僅提高了根系對土壤水分和養分利用效率,也間接反映了根系對環境變化的積極響應[8]。因此,在環境條件發生變化時,植物通過不斷改變自身根系形態和生理生化特性以調整生長策略適應環境[9]。已有研究表明,隨著水分條件的改善,植物根系細根長、根表面積[10]、根冠比[6]逐漸增大。但也有研究發現,隨著水分梯度的下降,不同退化程度草地內的植物均表現為主根長、根系體積和根表面積逐漸增加[11]。還有研究指出,可能由于根系周轉過快,隨著降水量的增加,植物根系生產未發生顯著改變[12]。可見,降水變化對植物根系形態特征的影響的研究結果因試驗條件及物種的不同而存在差異。
植物種間主要靠地下根系與土壤之間的物理、化學和生物過程表現出的競爭來實現對資源的合理分配利用[13]。在判定資源競爭利用強度方面,地下部分明顯重要于地上部分,從地下根系形態和分布特征來認識物種間競爭和互惠現象,才能更深入地了解種間關系的實質[14]。有研究表明種間關系會對植物根系形態產生影響,王政權等[15]發現水曲柳(Fraxinusmandshurica)和落葉松(Larixgmelinii)混栽顯著增大了水曲柳細根、粗根生物量及根長,提高了水曲柳競爭效率;張彥東等[16]和廖利平等[17]對不同混交林的研究均得出類似的結論。然而由于荒漠生態系統的復雜性和植物地下部分的不可見性及研究方法的局限性,目前人們對荒漠植物根系間的相互作用關系了解甚少[18]。針對植物根系形態特征對不同降水量的響應的研究主要集中在某單一植物種,對荒漠灌木混生植物群落的相關研究極其缺乏。因此,結合種間關系開展不同降水量條件下荒漠植物的根系形態變化特征的研究,對揭示干旱荒漠區物種共存和生物多樣性維持具有重要意義。
紅砂(Reaumuriasoongorica)和珍珠(Salsolapasserina)是干旱荒漠和荒漠草原區分布最廣的植物種之一,兩者常互為共建種組成混生群落[19]。混生群落分布生境類型多樣,珍珠主要見于年降雨量100~200 mm的低山丘陵、山間盆地和山前地帶;紅砂除此以外,還遍布于年降雨量100 mm以下的遼闊山前平原和廣大的戈壁灘[20]。研究發現兩者組成的混生群落適應極端環境變化的能力要顯著高于單生群落[21],混生群落對提高植物在極端環境內對水分和養分的吸收利用效率及維持生態穩定方面具有重要作用[22]。近年來,針對紅砂和珍珠混生后根系分布規律有零星報道。例如,研究發現相同生境下紅砂和珍珠混生后,混生紅砂根系生物量大于單生紅砂,珍珠則相反[23]。也有研究指出兩種植物混生后,在適宜水分下紅砂細根的生長得到促進,而珍珠是在水分脅迫嚴重時通過增加細根比重提高其競爭力;低水條件下混生后,紅砂和珍珠的地下生物量以及根冠比顯著降低,珍珠細根的比根長和比根面積顯著降低[21]。可見,兩種植物為適應不同的生境其各自的根系形態會產生不同變化。因此,本研究擬通過人工模擬自然降水變化,探討不同降水量條件下兩種生長方式的紅砂和珍珠的根系形態特征變化,以此揭示兩種荒漠植物根系形態特征變化對降水量和生長方式的響應機制,為深入開展脅迫環境下的混生植被群落間的相互作用關系的研究提供一定的理論依據。
1 材料和方法
1.1 材料培養
2017年3月底4月初土壤解凍之前,在甘肅省白銀市景泰縣(36°43′ N,104°43′ E)紅砂和珍珠生長的山前戈壁進行樣方調查。根據樣方調查數據,以自然生境中紅砂和珍珠的平均樣株大小為標準,分別選取株高(10.0±0.2)cm、冠幅(10.0±0.2)cm,根系形態基本一致的一年生單生紅砂、單生珍珠以及混生紅砂和混生珍珠幼苗各36 株,將所有幼苗挖掘后帶回校內試驗地,對幼苗根系進行適當修剪后分別定植到自行設計的內徑為15 cm、高度為80 cm的PVC管內,管內裝入取自紅砂和珍珠采樣地的自然生境中的黃土,加入少量蛭石和珍珠巖混合,避免土壤板結影響植物根系生長。緩苗1個月,為保證幼苗成活對幼苗進行水分和病蟲害管理。試驗過程中為防止水分流失和土壤下沉,在管子底部均放置了面積稍大于管子底面面積的托盤。試驗地位于甘肅農業大學校內試驗場,場地周圍開闊,無遮擋物,光照適宜。在試驗地搭設了簡易遮雨棚,出現降雨天氣時對紅砂和珍珠進行及時遮蓋,降雨停止后,及時取走防水篷布,保證植物可以進行充分的光合作用和呼吸作用。
1.2 試驗設計
因紅砂和珍珠混生群落由東向西氣候形成光-溫-濕的明顯梯度和多種多樣的水熱組合,在我國形成了從荒漠草原區、典型荒漠區到極端荒漠區的荒漠環境梯度分布格局,本試驗模擬3種荒漠環境梯度下的降水條件,自西向東分別選取酒泉、張掖、武威、銀川、白銀、蘭州6個地點的年均降水量作為參照(6個參照點的水熱條件見表1),人工模擬降水量分別設置為50,100,150,200,250和300 mm。6個選取點自西向東隨著自然降水量的增加,自然生境中紅砂和珍珠的植物密度(即平均每平方米上的株數)均逐漸增大,紅砂植株密度范圍為0.61 株·m-2~1.57 株·m-2,珍珠植株密度范圍為0.08 株·m-2~1.02 株·m-2。調查研究發現,西北荒漠區氣候干燥,降水少而不穩定,分配極不均勻,降水主要集中在夏秋季,占全年降水量70%~80%[24]。植物生長集中于夏季,在秋季進入凋零期。因此在模擬降水量變化的試驗中,計算出6,7,8,9這4個月的月降水量占比并分別換算為澆水量。降水量換算方法參考閆建成等[25]和寇祥明等[26]。具體如下:在直徑為20 cm的雨量器內,每降水31.4 mm相當于自然降水1 mm。

表1 試驗設計參照的試驗6個選取點的水熱條件比較
(1)
式中,A代表降水量,S代表降水面積,即在面積為S的地面上降水AmL相當于1 mm降水。單生紅砂、單生珍珠、混生紅砂和混生珍珠分別設置6種降水處理,每種降水處理下設置3個重復。年降水量為50 mm和100 mm的澆水頻次為4次·月-1,150 mm和200 mm的澆水頻次為6次·月-1,250 mm和300 mm的澆水頻次為8次·月-1,具體模擬的降水總量和單次降水量見表2。澆水時間從6月1日起至9月30日結束,降水時間設在19點,同時對植被進行等質精細化管理,及時除草和預防病蟲害。

表2 降水量占比及澆水量換算
1.3 根系取樣及相關指標計算
在植物的生長季末,將所有植株進行全根挖掘,以統計根系分布特征和植株生物量。收獲時,首先將已經減去植株地上部分的PVC管小心傾倒至地面用小刀割開,至此可以看到完整含根系的土壤,用鋒利的刀小心地將完整的、圓柱狀的土壤切開,將每一次取得的含根土壤置于200 μm的篩一遍遍過濾,篩去土粒,挑出石子等雜質后將根系清洗干凈,置于玻璃皿中。其中,混生紅砂和珍珠的根系根據顏色、形態和柔韌性進行區分。將清洗干凈后的根系用電子游標卡尺分成粗根(直徑≥2 mm)和細根(直徑<2 mm),然后分裝置于不同的信封并標記。將這些根樣用掃描儀掃描,最后用根系分析軟件(Win-RhIZO 2008a)得到植物根系的形態特征指標,如根長、根表面積等。將掃描過的根系重新裝入信封,與植物的地上部分一同放入75℃的烘箱烘至恒重,得到植物地上和地下部分的干重。其余指標計算方法如下:
1.4 數據統計分析
所有數據采用SPSS 22.0進行統計分析,生長方式和降水量對植物根系形態指標的影響采用雙因素方差分析;對同一降水量不同生長方式下同種植物根系形態指標進行了單因素方差分析;對同一生長方式不同降水量下同種植物根系形態指標進行了單因素方差分析。作圖采用Origin 2018,圖中數值均為平均值±標準誤差。
2 結果與分析
2.1 不同降水量下單生和混生紅砂與珍珠的細根和粗根的生物量及其比重比較
由表3可知,生長方式對紅砂細根生物量有極顯著影響(P<0.01),降水量對紅砂細根和粗根生物量影響均顯著(P<0.05)。生長方式對珍珠細根、粗根生物量、細根生物量比重、粗根生物量比重影響均為極顯著(P<0.01),降水量對其生長無顯著影響。

表3 不同降水量條件下單生、混生紅砂珍珠的細根和粗根生物量及其比重的雙因素方差分析
隨著降水量的增加,兩種生長方式下的紅砂細根生物量均呈現增-減-增的趨勢,且同一降水量下的單生紅砂細根生物量均大于混生紅砂,100 mm和150 mm降水量下差異顯著(P<0.05),單生紅砂細根生物量均值較混生紅砂高81.21%(圖1a)。由圖1b可知,同一降水量條件下,單生珍珠細根和粗根生物量均分別大于混生珍珠,均在300 mm降水量下達到顯著差異(P<0.05),均值分別高210.35%和367.06%。單生珍珠細根生物量隨降水量的增加呈增-減-增的變化趨勢,300 mm降水量下的最大值比50 mm降水量下的最小值高82.73%。
在50 mm和100 mm降水量條件下,混生紅砂細根生物量比重大于單生紅砂,但隨著降水量的增加,其余降水量下單生紅砂細根生物量比重均大于混生紅砂(圖1c)。同一降水處理下的混生珍珠細根生物量比重均大于單生珍珠,粗根生物量比重均小于單生珍珠(圖1d)。同一降水量下的單生紅砂的細根生物量比重均大于單生珍珠;除250 mm降水量外,其余降水量條件下混生紅砂的細根生物量比重均大于混生珍珠(圖1c,1d)。

圖1 不同降水量條件下單生、混生紅砂珍珠的細根、粗根生物量及其比重
2.2 不同降水量下單生和混生紅砂與珍珠的細根和粗根的比根長比較
由表4可知,生長方式、降水量及兩者的交互作用對紅砂比根長均無顯著影響,但生長方式對珍珠細根比根長有極顯著影響(P<0.01)。

表4 不同降水量條件下單生、混生紅砂珍珠的細根和粗根比根長的雙因素方差分析
由圖2a可知,50 mm降水量下單生和混生紅砂細根比根長差異顯著(P<0.05)。在50~200 mm降水量范圍內,單生和混生紅砂細根比根長均呈現先增后減變化趨勢。200~300 mm降水量范圍內,單生紅砂細根比根長先減后增,而混生紅砂細根比根長先增后減。由圖2b可知,150~300 mm降水量范圍內,混生珍珠細根比根長顯著大于單生珍珠(P<0.05)。在50~200 mm降水量范圍內,單生珍珠細根比根長呈現減-增-減的變化趨勢,而混生珍珠則呈現相反變化趨勢(增-減-增),200~300 mm降水量內,單生和混生珍珠細根比根長均先增大后減小。

圖2 不同降水量條件下單生、混生紅砂珍珠的細根、粗根比根長
2.3 不同降水量下單生和混生紅砂與珍珠的細根和粗根的比根面積比較
由表5可知,生長方式對珍珠細根比根面積影響顯著(P<0.05),降水量及兩者的交互作用對兩種植物根系比根面積均無顯著影響。

表5 不同降水量條件下單生、混生紅砂珍珠的細根和粗根比根面積的雙因素方差分析
兩種生長方式下,紅砂細根比根面積變化均未呈線性變化,單生紅砂細根比根面積在100 mm降水量下的最大值與200 mm降水量下的最小值差異顯著(P<0.05),最大值比最小值高89.24%。混生紅砂粗根比根面積隨降水量的增加逐漸減小(圖3a)。由圖3b可知,對珍珠而言,在100~300 mm降水量范圍內,混生細根比根面積均大于單生,其均值較單生增加了84.00%。單生珍珠細根比根面積在50 mm處的最大值與在300 mm處的最小值差異顯著(P<0.05),最大值較最小值高137.36%;混生珍珠細根比根面積在200 mm處的最大值與降水量為50 mm時的最小值差異顯著(P<0.05),最大值較最小值高180.25%。
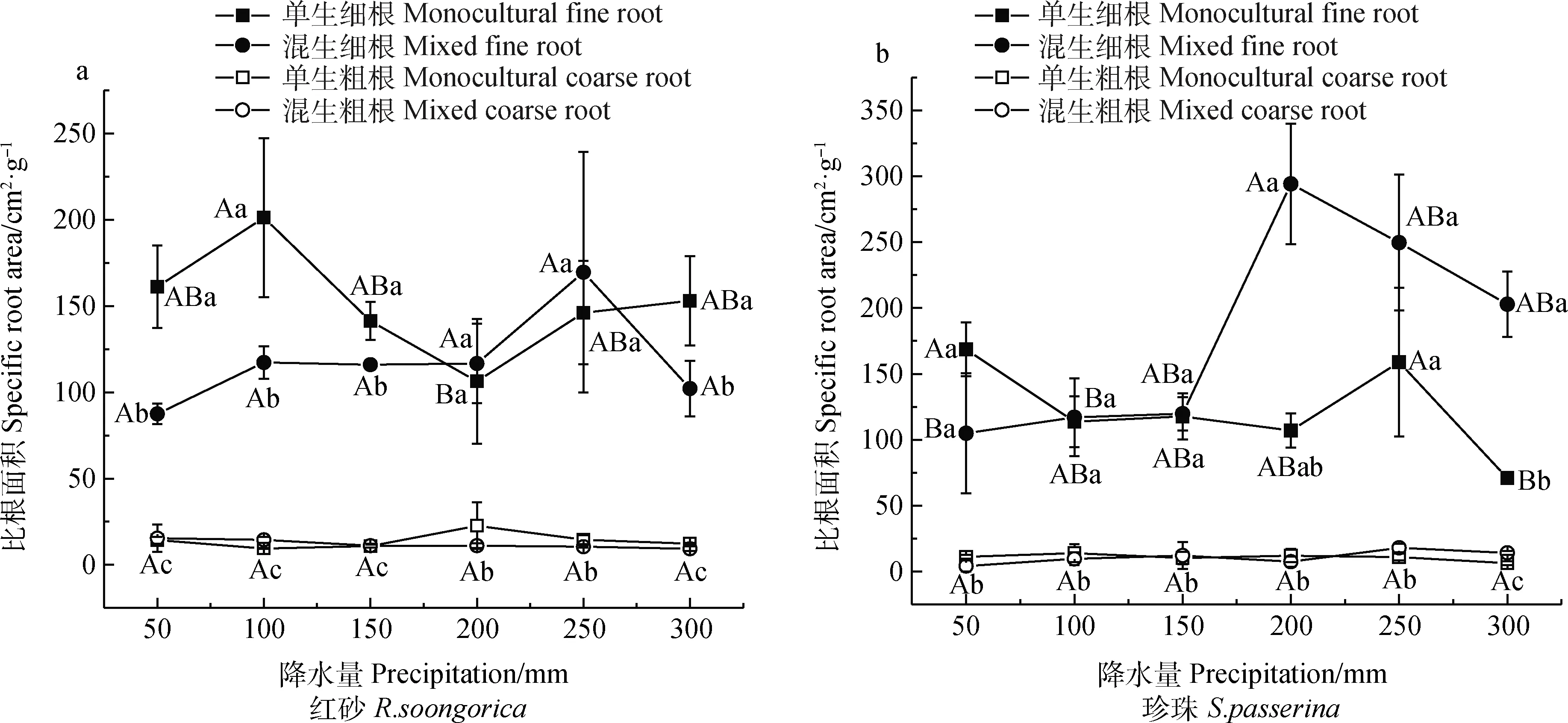
圖3 不同降水量條件下單生、混生紅砂珍珠的細根、粗根比根面積
2.4 不同降水量下單生和混生紅砂與珍珠的根冠比比較
由圖4a可知,在同一降水量條件下,混生紅砂根冠比均小于單生紅砂。隨著降水量的增加,混生后紅砂根冠比較單生分別降低了5.00%,12.50%,28.81%,21.43%,41.46%,14.55%。在150 mm降水量條件下,單生紅砂根冠比達到最大值(0.59)。
由圖4b可知,混生珍珠在250 mm降水量處理下根冠比達到最大值(0.58),與其他降水量條件下的根冠比值均差異顯著(P<0.05),比其它降水量條件下的根冠比分別高58.62%,58.62%,60.34%,70.69%,48.28%,比該降水量處理下的單生珍珠根冠比高56.90%。同一降水量下單生紅砂的根冠比均大于單生珍珠,其均值較單生珍珠高44.90%(圖4a,4b)。

圖4 不同降水量下單生、混生紅砂珍珠的根冠比
3 討論
3.1 混生對紅砂-珍珠根系形態特征的影響
物種共存的一條重要策略是相鄰同種和異種個體由于根系競爭調整其根系的空間分布格局[23,27],這樣的共存機制總體上有利于兩種植物協同適應各種脅迫生境[28]。但也有研究指出,干旱環境中緊密共存的物種之間的根系分離不是普遍現象[29]。研究表明,相鄰植物可通過改變比根長和比根面積來適應根系間的相互競爭實現共存[30-31]。本研究發現,紅砂和珍珠混生顯著提高了珍珠的細根比根長和比根面積,降低了紅砂細根比根長和比根面積,這與張海娜[21]對紅砂和珍珠的研究結果一致。同樣,對草本、喬木和農作物的相關研究也發現混交或混播會提高其中一種植物的細根比根長和比根面積[32-34]。說明紅砂和珍珠混生后,珍珠通過不斷增大細根比根長和比根面積來擴大根系與水分和養分的接觸面積,提高自身競爭力以獲取適合自身生長所需的水分和養分。紅砂細根比根長和比根面積降低,這可能是因為兩種植物混生后根系的重疊主要促進珍珠根系的生長,并改善了其周圍土壤的緊實度,從而對紅砂根系產生庇護效應[21],因此紅砂沒必要通過增加自身根長和根表面積等形態特征指標來消耗更多的水分和養分對根系進行重建投資[35],而是選擇將更多的水分和養分輸送至地上部分以滿足地上部分的生理需求。可見,不同植物混生生長時,植物會通過整合資源和相鄰植物信息來調整自身根系形態和資源分配策略[36]。
有研究指出不同種的植物,其根系交錯會促進細根的生長,更有利于植物從周圍環境中獲取養分和水分,對粗根卻幾乎不產生促進作用[37];Dudley和File[38]發現某些植物在與其它植物發生種間競爭時會分配給細根更多的生物量;Padilla等[39]的研究發現植物對水分和養分的吸收與土壤單位體積中細根生物量之間存在著很強的關系,但與粗根生物量無關。本研究發現,混生顯著提高了珍珠細根生物量比重,降低了粗根生物量比重,這與張海娜[21]研究發現將紅砂和珍珠混生,細根會產生更多的重疊,促進了珍珠細根的伸長生長的結果一致。說明兩種植物混生后,珍珠地下部分的競爭能力增強,珍珠將更多的養分水分分配到細根,促進細根的生長發育,細根數量的增加使總根系對水分和養分的獲取能力得以補充,根系地下部分的生物量增大,使植物能夠更好的利用地下資源促進自身生長發育,體現了珍珠為適應環境變化對自身粗細根的資源投入產生的一種分配策略。究其原因可能是因為混生加劇了兩種植物對資源的競爭利用強度,而根系木質化程度會影響植物對脅迫條件的抵抗能力[40]。細根(直徑<2 mm)作為植物組織中的幼嫩部分,多數未木質化,其對土壤環境因子的變化反應極其敏感,粗根多為木質化組織,其對環境變化的敏感程度較低[40]。紅砂和珍珠混生后根系對地下資源的競爭作用加強,珍珠作為淺根性植物,由于土壤表層水分養分資源豐厚[23],其細根能吸收充足的水分養分促進自身細根生物量增加,從而提高了細根比重。而深根性植物紅砂的細根在養分水分較少的深層土壤中獲取資源則處于一定的劣勢[23],只能不斷下扎自身根系探索更深層能夠促進自身發育的資源,越深層的土壤中細根生物量越小,這也體現了兩種植物混生后根系發生的生態位分離。可見,細根在植物根系競爭過程中在吸收養分水分方面展現的較強的可塑性和活躍性。
3.2 降水量對紅砂-珍珠根系形態特征的影響
荒漠環境中植物生長所需水分主要來源于自然降水,根系作為最先感知到水分存在及變化的器官,當生境中水分減少時,根系首先產生化學信號并迅速傳遞至地上部分促使氣孔關閉,減少水分蒸騰散失;與此同時,地下根系也會通過調整自身形態特征和生物量分配策略來適應環境水分變化[5]。比根長和比根面積是指示根系功能的重要指標,可綜合反映植物吸收資源的能力與生態適應性[41],其值越大,根系吸收水分和養分的能力越強,表明投入相同的成本會產生更高的養分利用效率[23]。很多研究發現,在降水減少的情況下,植物根系的比根長和比根面積會顯著增加[42-45]。但也有研究[46-47]指出,干旱條件下植物保存水分的策略之一是降低比根長,減少細根生長。本研究發現,隨著降水量的增加,單生紅砂細根比根長和比根面積均呈現增-減-增的趨勢,這與種培芳等[48]研究發現降水量增多,單生紅砂比根長和比根面積逐漸降低的結果不一致,這可能是試驗處理和降水梯度不同導致的差異。自然界中紅砂主要分布在100 mm~200 mm降水量范圍的生境中,本研究發現在該降水范圍內,單生紅砂細根比根長和比根面積均隨降水量的增加逐漸減小,這與前人研究結果一致[48-49]。此外,本研究發現,相同降水量下的單生紅砂根冠比均大于單生珍珠。這可能是因為紅砂為深根型植物,維持其生長的水源主要來自于地下水,且紅砂在脅迫條件下其根系會不斷下扎拓寬生長區域,從而增大地下生物量;大氣降水作為維持非深根型植物珍珠生長的主要水源[23],會對珍珠地上生物量的增加產生促進作用,導致相同生境中單生珍珠的地上部分生物量顯著大于單生紅砂[21]。因此,在相同降水條件下,紅砂深層的地下根系吸收水分養分能力較強,導致地下生物量顯著大于珍珠[50],而地上生物量小于珍珠,更易于形成大的根冠比促進根系發育。這與Schwinning等[51]發現在干旱區,以深層土壤水為主要水源的植物,一般以深根性、大根冠比為主要表現特征;以不定降水為主要水源的植物則表現為淺根性、小根冠比的結論一致。Passiour等[52]研究也指出植物根冠比大,將會有更多的同化產物分配到根系,更有利于根系的快速擴張。但無論環境如何變化,植物會通過調整自身根系形態,增加對養分和水分吸收的有效性,以期最大化地滿足自身的生長需求[53]。
4 結論
混生降低了紅砂各根系形態特征指標的大小,而顯著增加了珍珠根系形態特征各指標的值,說明兩者混生時紅砂在根系競爭中表現出一定的劣勢,而混生提高了珍珠的競爭能力,更有利于珍珠塑造良好的根系形態特征以提高其根系對水分養分的吸收利用效率。兩種植物混生生長后,與粗根相比,珍珠細根的各形態指標均隨水分條件變化明顯,說明其細根對根系間相互作用的敏感程度高于粗根,混生生長過程中主要通過調整細根根系形態發育來適應水分變化。可見,在我國西北廣泛分布的紅砂-珍珠混生群落中,兩種植物均可通過各自的生存策略調整根系形態特征,從而降低競爭強度形成共存。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
