腐植酸對(duì)干旱脅迫下燕麥葉片碳同化酶活性及產(chǎn)量的影響
2021-09-23 08:41:08朱珊珊米俊珍趙寶平孫夢(mèng)媛李英浩劉景輝
麥類(lèi)作物學(xué)報(bào) 2021年6期
關(guān)鍵詞:產(chǎn)量
朱珊珊,米俊珍,趙寶平,孫夢(mèng)媛,李英浩,劉景輝
(內(nèi)蒙古農(nóng)業(yè)大學(xué)農(nóng)學(xué)院,內(nèi)蒙古呼和浩特 010019)
燕麥?zhǔn)俏覈?guó)特色糧飼兼用型作物,具有營(yíng)養(yǎng)成分高,抗逆性強(qiáng)等優(yōu)點(diǎn),在我國(guó)西北、華北、西南等干旱半干旱少雨地區(qū)種植面積較大,干旱是限制其生長(zhǎng)發(fā)育和高產(chǎn)的重要因素[1]。干旱脅迫會(huì)抑制作物旗葉光合作用的酶活性[2],影響光合碳同化,導(dǎo)致作物減產(chǎn)[3]。嚴(yán)重干旱會(huì)導(dǎo)致植物葉片碳同化酶活力降低[4-5]。磷酸烯醇式丙酮酸羧化酶(PEPC)可通過(guò)植物內(nèi)部釋放的二氧化碳和氮來(lái)提供碳骨架以進(jìn)行生物合成,緩解干旱脅迫[6-7]。光合產(chǎn)物的積累量和作物產(chǎn)量與其葉片中甘油醛-3-磷酸脫氫酶(GAPD)和乙醇酸氧化酶(GO)活性有關(guān)[4,8]。核酮糖-1,5-二磷酸羧化酶(Rubisco)、GO、PEPC及GAPD是植物碳同化的關(guān)鍵酶,提高其效率和活力是促進(jìn)植物光合作用的重要途徑[4,8-10]。腐植酸是存在于土壤、河泥和煤炭之中的天然有機(jī)混合物[11],有促進(jìn)作物生長(zhǎng)、節(jié)肥增效、增強(qiáng)作物抗逆性等功效,是一種多功能葉面肥[12]。張志芬[13]研究發(fā)現(xiàn),噴施腐植酸可抑制燕麥葉片的氣孔開(kāi)度,增加氣孔阻力,降低蒸騰作用,緩解燕麥干旱脅迫。腐植酸能夠緩解干旱對(duì)麻黃葉片中葉綠體片層結(jié)構(gòu)的破壞,增加葉片的葉綠素含量和凈光合速率,顯著減輕由干旱脅迫造成的氧化損傷,提高麻黃的耐旱性,從而提高其產(chǎn)量[14]。梁太波[15]研究發(fā)現(xiàn),腐植酸肥料可以提高旱作小麥旗葉實(shí)際光化學(xué)效率和光合電子傳遞速率,延緩衰老,提高水分利用效率。以上研究表明,干旱脅迫下,腐植酸類(lèi)物質(zhì)能通過(guò)改變作物葉綠素?zé)晒鈪?shù)和光合參數(shù)、降低電子傳輸通量、提高植物體內(nèi)氣體交換率,顯著提高作物光合作用、抗旱能力以及產(chǎn)量[16-18]。但有關(guān)干旱脅迫下腐植酸對(duì)燕麥葉片的碳同化酶活性及產(chǎn)量影響的研究鮮有報(bào)道。本研究以燕科2號(hào)為材料,分析不同程度干旱脅迫下噴施腐植酸對(duì)燕麥不同生育時(shí)期葉片碳同化酶活性和產(chǎn)量的影響,以期為燕麥抗旱、增產(chǎn)措施的制定提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料及設(shè)計(jì)
試驗(yàn)材料為燕科2號(hào),由內(nèi)蒙古農(nóng)牧業(yè)科學(xué)院提供。腐植酸水溶肥料采購(gòu)于內(nèi)蒙古永業(yè)農(nóng)豐生物技術(shù)有限責(zé)任公司,其水溶腐植酸含量≥50 g·L-1,N+P+K≥200 g·L-1,微量元素(錳、硼、鉬、鋅等)≥10 g·L-1。于2019年4月至8月在內(nèi)蒙古農(nóng)業(yè)大學(xué)燕麥產(chǎn)業(yè)研究中心溫室進(jìn)行盆栽試驗(yàn)。試驗(yàn)選用直徑為23 cm、高18 cm的塑料盆進(jìn)行,內(nèi)裝混合土5 kg(河沙∶泥炭土體積比1∶1),泥炭土中N+P+K> 10 g·kg-1,有機(jī)質(zhì)含量> 50 g·kg-1,pH值為5.5~8.5,土壤含水率為9%。播種前每盆施入2 g磷酸二銨(N 18%,P2O546%)做底肥。設(shè)置3個(gè)水分處理:正常供水(田間最大持水量的75%,M1)、中度干旱脅迫(田間最大持水量的45%,M2)和重度干旱脅迫(田間最大持水量的30%,M3);2個(gè)噴施處理:噴施500倍稀釋的腐植酸水溶肥料(HA)和等量清水(CK),共6個(gè)處理。采用隨機(jī)區(qū)組排列,每個(gè)處理重復(fù)3次,每重復(fù)種植10盆,每盆播種30株,三葉期定苗為20株。2019年4月14日播種,8月21日收獲。從拔節(jié)期(5月18日)開(kāi)始控水至成熟期,控水方法采用稱(chēng)重差值法[19]。分別在拔節(jié)期、抽穗期、灌漿期的上午 9:00-11:00進(jìn)行HA和CK處理,在植株葉片均勻噴施至葉面濕透無(wú)滴水,每盆噴施量約14 mL;噴施7 d后取燕麥中上部葉片,投入液氮速凍后,放入-80℃冰箱冷藏,用于碳同化酶活性的測(cè)定。
1.2 測(cè)定項(xiàng)目與方法
1.2.1 碳同化酶測(cè)定
核酮糖-1,5-二磷酸羧化酶(Rubisco)、磷酸烯醇式丙酮酸羧化酶(PEPC)和乙醇酸氧化酶(GO)活性均采用蘇州科銘生物技術(shù)有限公司試劑盒測(cè)定,3-磷酸甘油醛脫氫酶(GAPD)活性采用上海繼錦化學(xué)科技有限公司試劑盒測(cè)定。
1.2.2 地上部干物質(zhì)積累量和產(chǎn)量及其構(gòu)成因素測(cè)定
地上部干物質(zhì)積累量:分別于拔節(jié)、抽穗、灌漿期葉面噴施處理后7 d取樣,每個(gè)重復(fù)取10株,放入紙袋,烘箱105℃殺青30 min,80℃烘干到恒重。
產(chǎn)量及構(gòu)成因素:燕麥成熟時(shí)收獲、考種,每個(gè)處理取5株, 測(cè)定主穗長(zhǎng)、單株小穗數(shù)、穗粒數(shù)、穗粒重和千粒重,并測(cè)每盆的籽粒產(chǎn)量,重復(fù)3次。
1.3 數(shù)據(jù)統(tǒng)計(jì)分析
采用Excel 2010進(jìn)行數(shù)據(jù)處理,用SPSS 22.0進(jìn)行顯著性和相關(guān)性分析。
2 結(jié)果與分析
2.1 腐植酸對(duì)不同水分處理燕麥葉片Rubisco活性的影響
由圖1可見(jiàn),從拔節(jié)期到灌漿期,不同水分處理燕麥葉片的Rubisco活性均呈先升高后降低趨勢(shì)。干旱脅迫導(dǎo)致葉片Rubisco活性顯著降低。M1水分條件下,HA處理拔節(jié)期、抽穗期和灌漿期燕麥葉片Rubisco活性較CK分別增加 5.62%、2.79%、6.88%,僅在灌漿期差異達(dá)到顯著水平。M2水分條件下,HA處理各時(shí)期葉片Rubisco活性較CK均顯著提高,拔節(jié)期、抽穗期和灌漿期分別增加了22.38%、25.83%、 19.17%。水分條件為M3下,拔節(jié)期、抽穗期和灌漿期HA較CK處理葉片Rubisco活性分別增加了28.45%、34.41%、32.22%,差異均達(dá)到顯著水平。以上結(jié)果說(shuō)明,干旱脅迫顯著降低了燕麥葉片Rubisco活性;噴施腐植酸可有效緩解干旱脅迫對(duì)燕麥葉片Rubisco活性的影響,尤其在M3水分條件下效果更佳。
2.2 腐植酸對(duì)不同水分處理下燕麥葉片GAPD活性的影響
隨著干旱程度的加深,各時(shí)期葉片中的GAPD活性均呈降低趨勢(shì)(圖2)。與CK相比,M1水分條件下噴施腐植酸可以提高GAPD活性,但只在抽穗期效果顯著;在M2和M3水分條件下,噴施腐植酸均能使各時(shí)期GAPD活性顯著增加,其中在M3水條件下增加的幅度最大,拔節(jié)期、抽穗期和灌漿期分別增加36.55%、31.80%、39.60%。這說(shuō)明噴施腐植酸可不同程度地減小干旱脅迫對(duì)燕麥葉片GAPD活性的影響,在M3處理下效果較好。
2.3 腐植酸對(duì)不同水分處理下燕麥葉片PEPC活性的影響
由圖3可知,燕麥葉片的PEPC活性在抽穗期達(dá)到最高。在M1水分條件下,噴施腐植酸后燕麥葉片PEPC活性與CK相比僅在拔節(jié)期差異顯著;當(dāng)水分條件為M2和M3時(shí),噴施腐植酸后葉片中的PEPC活性較CK在各時(shí)期均顯著增加,其中在M2水分條件下,拔節(jié)期、抽穗期和灌漿期噴施腐植酸較CK分別增加了21.43%、 24.04%、25.42%,M3水分條件下,拔節(jié)期、抽穗期和灌漿期噴施腐植酸較CK分別增加了 38.24%、32.96%、33.81%。這說(shuō)明噴施腐植酸提高了干旱脅迫下燕麥葉片的PEPC活性,且M3水分條件下提升幅度最大。
2.4 腐植酸對(duì)不同水分處理下燕麥葉片GO活性的影響
從拔節(jié)期到灌漿期,燕麥葉片GO活性先降低后升高(圖4)。干旱脅迫導(dǎo)致燕麥葉片GO活性升高,且隨著干旱程度的加劇而增加。在M1水分條件下,噴施腐植酸后燕麥葉片GO活性與CK相比差異均未達(dá)到顯著水平;當(dāng)水分條件為M2和M3時(shí),噴施腐植酸后葉片GO活性在各時(shí)期與CK相比均顯著降低,其中M2條件下,拔節(jié)期、抽穗期和灌漿期較CK分別降低16.18%、13.64%、14.65%,M3條件下,拔節(jié)期、抽穗期和灌漿期分別降低16.85%、16.75%、18.39%。這表明干旱脅迫顯著提高了燕麥葉片 GO的活性,噴施腐植酸能有效緩解這種增長(zhǎng)趨勢(shì)。
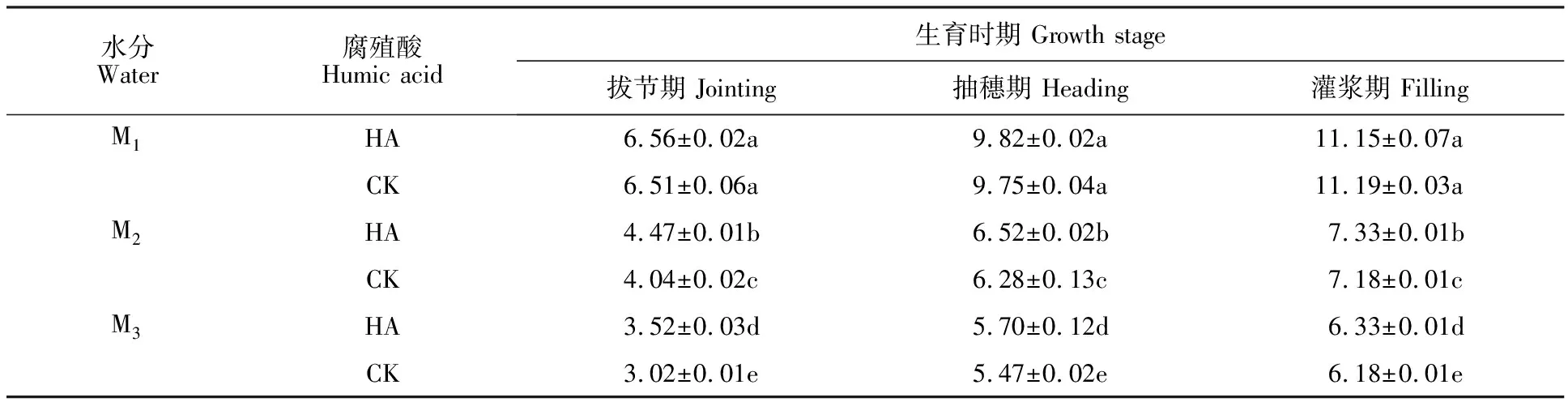
表1 不同處理下燕麥地上部干物質(zhì)積累量的影響Table 1 Effect of different treatments on above-ground dry matter mass of oat g·10-1 plants
2.5 腐植酸對(duì)不同水分處理下燕麥地上部干物質(zhì)積累量的影響
由表1可知,隨著干旱程度的加深,燕麥地上部干物質(zhì)積累量顯著減少。與CK相比,在M1水分條件下,HA處理對(duì)燕麥各時(shí)期地上部干物質(zhì)積累量沒(méi)有顯著影響;在M2和M3水分條件下,拔節(jié)期、抽穗期、灌漿期HA處理燕麥地上部干物質(zhì)積累量均較CK顯著增加,增幅分別為10.64%、 3.82%、2.09%和16.56%、4.20%、 2.43%。
2.6 腐植酸對(duì)不同水分處理下燕麥產(chǎn)量及其構(gòu)成因素的影響
表2顯示,干旱脅迫導(dǎo)致燕麥產(chǎn)量及其構(gòu)成因素均顯著下降,且均表現(xiàn)為M1>M2>M3。與CK相比,噴施腐植酸能使燕麥產(chǎn)量增加,M1、M2、M3水分條件下分別增加2.63%、8.46%和10.26%,其中在M2和M3水分條件下差異達(dá)到顯著水平。從產(chǎn)量構(gòu)成因素來(lái)看,噴施腐植酸后,穗長(zhǎng)、穗粒重和千粒重較CK均增加,增加幅度均表現(xiàn)為M3>M2>M1,其中穗粒重和千粒重在M2和M3水分條件下增加顯著,分別增加 15.38%、16.07%和10.74%、15.67%;與CK相比,噴施腐植酸可以提高燕麥單株小穗數(shù)、穗粒數(shù)、穗數(shù),但差異均不顯著。說(shuō)明噴施腐植酸主要通過(guò)降低干旱對(duì)千粒重的影響而增加產(chǎn)量。
2.7 碳同化酶活性與產(chǎn)量及其構(gòu)成因素的相關(guān)性分析
Rubisco活性與穗粒重和產(chǎn)量的相關(guān)性均達(dá)到極顯著水平,與千粒重呈顯著正相關(guān),與穗粒重的相關(guān)性最大。PEPC和GAPD活性與穗粒重、千粒重和產(chǎn)量均呈極顯著正相關(guān),與穗粒重的相關(guān)性最大。GO活性與穗粒重、千粒重和產(chǎn)量均極顯著負(fù)相關(guān),與產(chǎn)量的相關(guān)性最大。穗粒重與千粒重與產(chǎn)量呈極顯著正相關(guān)(表3)。
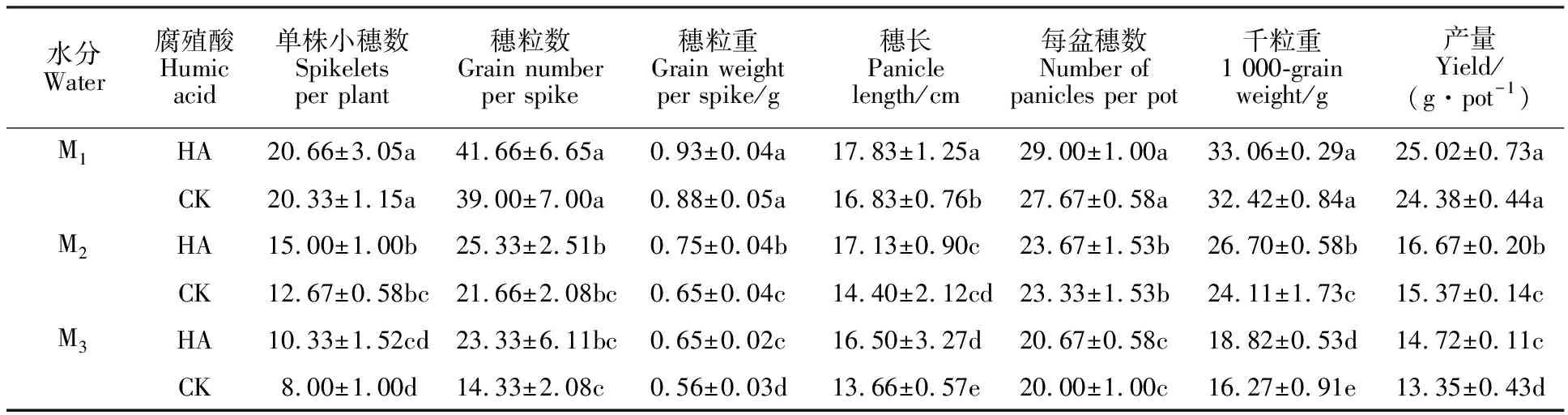
表2 不同處理對(duì)燕麥產(chǎn)量及其構(gòu)成因素的影響Table 2 Effect of different treatments on oat yield and its components
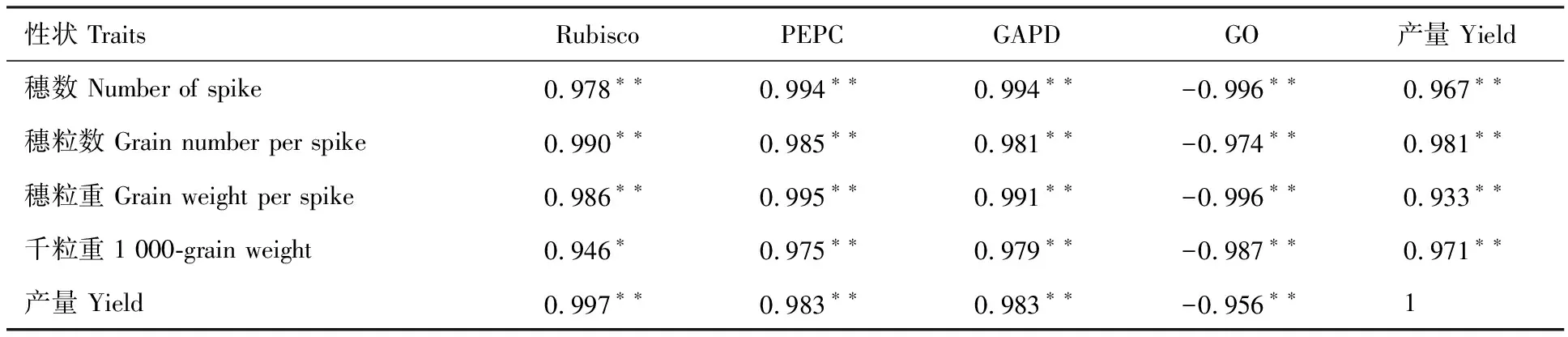
表3 燕麥葉片碳同化酶活性與產(chǎn)量及其構(gòu)成因素的相關(guān)性分析Table 3 Correlation analysis of carbon assimilating enzyme activity in oat leaves with yield and its components
3 討 論
植物的光合作用是干物質(zhì)積累和產(chǎn)量形成的基礎(chǔ),較高的光合碳同化能力是獲得高產(chǎn)的前提。卡爾文循環(huán)中,Rubisco 是影響光合碳同化的關(guān)鍵酶,能使逆境脅迫中的植物維持光合作用[20]。干旱脅迫會(huì)導(dǎo)致 Rubisco 活性降低,光合作用產(chǎn)生限制[16,21-22],本研究結(jié)果與此相似,隨著干旱脅迫程度的增加,Rubisco活性逐漸下降,可能干旱脅迫使相應(yīng)酶量降低,進(jìn)而導(dǎo)致 Rubiso 活性下降,影響了光合碳同化[23]。在重度干旱脅迫中,作物葉片光合能力的降低主要受非氣孔因素限制,葉片光合放氧能力降低,光合磷酸化解偶聯(lián),阻礙CO2擴(kuò)散,RuBP羧化酶活性下降,光合器官的生理功能遭到破壞,影響作物光合作用[24-25],噴施腐植酸使植物在干旱脅迫下維持光合系統(tǒng)的穩(wěn)定和相對(duì)較高的光合作用,對(duì)非氣孔因素引起的光合限制具有緩解作用[26-27]。有研究表明,在氮脅迫下添加腐植酸比未添加處理RuBPCase活性顯著提高[28]。Jannin等[29]提出,腐殖質(zhì)處理后的植物Rubisco活性增加可能是由于細(xì)胞內(nèi)葉綠體數(shù)量增加所致。本研究表明,重度干旱脅迫下,噴施腐植酸后燕麥葉片中Rubisco酶活性與CK相比提高幅度最大,中度脅迫條件下其活性雖有提高,但幅度較小。這可能是由于腐植酸在重度干旱脅迫下更能緩解干旱對(duì)葉綠體的傷害[13],增加RuBP羧化酶固定CO2的量,進(jìn)而提高Rubisco的羧化效率,提高作物的光合能力。
GAPD是糖異生和糖酵解過(guò)程中的關(guān)鍵酶,催化3-磷酸甘油酸(3-PGA)還原成3-磷酸甘油醛,影響光合環(huán)的運(yùn)轉(zhuǎn)效率及光合產(chǎn)物的積累[30]。Ziaf等[31]發(fā)現(xiàn),干旱脅迫下GAPD基因的表達(dá)減輕了植物膜脂過(guò)氧化的程度。有研究表明,隨著水分脅迫時(shí)間的延長(zhǎng)和脅迫強(qiáng)度的加劇,小麥旗葉的GAPD活性逐漸降低[23],與本研究結(jié)果一致。在逆境條件下,生物體中GAPD基因的轉(zhuǎn)錄及表達(dá)水平不穩(wěn)定,大多數(shù)生物體都增加表達(dá)GAPD[32]。郭子平[33]研究表明,干旱脅迫下,小麥葉片總GAPD活性隨著時(shí)間的延長(zhǎng)而增加。在本研究中,隨著干旱脅迫程度的加劇,葉片中的GAPD活性逐漸降低,與上述研究結(jié)果不同,可能是因?yàn)椴煌购殿?lèi)型作物受到干旱脅迫時(shí)GAPD基因的表達(dá)調(diào)控因子和信號(hào)傳遞途徑存在差異,也可能是不同作物、不同品種的GAPD蛋白在植物細(xì)胞的失水耐受過(guò)程中作用不同[34]。有研究表明,腐植酸可以誘發(fā)含有NADP依賴(lài)性甘油醛-3-磷酸脫氫酶(GAPN)蛋白產(chǎn)生[35]。在本研究中,兩種干旱脅迫條件下噴施腐植酸,燕麥葉片的GAPD活性均能提高,但在重度脅迫下變化幅度較大。可能是因?yàn)楦菜嵩谥参锛?xì)胞過(guò)度失水時(shí)誘導(dǎo)了GAPDH蛋白的大量表達(dá),導(dǎo)致酶活性提高,促進(jìn)光合產(chǎn)物的積累,最終增加燕麥的穗粒重,使產(chǎn)量增加。
PEPC在CAM和C4植物中的主要生理作用是初級(jí)CO2的固定還原[36]。有研究表明,C3植物中也存在PEPC[37],在大部分C3植物體內(nèi)中,PEPC具有回補(bǔ)作用,補(bǔ)充三羧酸循環(huán),其中間產(chǎn)物能被用于多種生物合成及氮同化消耗[36]。關(guān)于干旱脅迫對(duì)PEPC活性的影響,各試驗(yàn)結(jié)果有所不同。有研究表明,在水分虧缺下,小麥PEPC活性明顯增強(qiáng)[18];而張 旭等[38]認(rèn)為,干旱導(dǎo)致小麥旗葉PEPC活性下降。玉米葉片中PEPC活性在水分脅迫下先升后降[39]。在本研究中,隨著干旱脅迫程度的加劇,PEPC活性均逐漸降低,有可能是不同種類(lèi)的植物應(yīng)答干旱的生態(tài)適應(yīng)性機(jī)制不同[40]。噴施腐植酸后,PEPC活性均有所提高,且土壤水分含量低時(shí)提升效果更佳,可能是在過(guò)度失水下,腐植酸在一定程度上更能誘導(dǎo)PEPC酶蛋白的表達(dá),具體原因還需進(jìn)一步探討。
GO在植物光呼吸代謝中具有關(guān)鍵作用,能提高植物光合效率[41]。隨著干旱脅迫的加劇,檸條中GO活性逐漸增強(qiáng)[42],與本研究研究結(jié)果相似。這說(shuō)明隨著干旱程度加劇,乙醇酸氧化酶活性也隨之升高。乙醇酸氧化酶催化乙醇酸的氧化過(guò)程中伴隨著 H2O2的生成,是H2O2總數(shù)的70%[43]。水分脅迫能夠增加GO活性,誘導(dǎo)大量H2O2的累積[44]。Garcar等[45]研究發(fā)現(xiàn),腐植酸可通過(guò)降低水稻H2O2的含量,提高膜的通透性,對(duì)干旱引起的氧化應(yīng)激反應(yīng)起到保護(hù)作用。本研究結(jié)果表明,腐植酸能使干旱條件下燕麥葉片GO活性降低,其中在重度干旱脅迫時(shí),噴施腐植酸較CK處理相比降低幅度更大。這說(shuō)明腐植酸可能是通過(guò)減少過(guò)氧化氫的生成來(lái)抑制乙醇酸氧化酶的活性,降低燕麥的光呼吸,增加作物的光合作用。
綜上,干旱脅迫通過(guò)改變碳同化酶活性,降低光合碳同化能力,從而影響燕麥的干物質(zhì)積累和產(chǎn)量形成,噴施腐植酸能顯著提高燕麥葉片Rubisco、GAPD和PEPC活性,提高碳的同化能力,降低GO活性,促進(jìn)光合作用,增加產(chǎn)量,當(dāng)田間最大持水量為30%提升幅度更大。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當(dāng)代水產(chǎn)(2021年10期)2021-12-05 16:31:48
今日農(nóng)業(yè)(2021年14期)2021-11-25 23:57:29
今日農(nóng)業(yè)(2021年13期)2021-08-14 01:37:56
石油與天然氣地質(zhì)(2021年1期)2021-02-22 14:14:44
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
中國(guó)果業(yè)信息(2019年10期)2019-11-13 01:21:34
中國(guó)化肥信息(2019年2期)2019-01-18 15:24:35
中國(guó)化肥信息(2019年1期)2019-01-17 21:31:12
中國(guó)化肥信息(2019年4期)2019-01-17 18:47:06
