跑臺運動對帕金森病模型小鼠黑質、海馬CA1區ERK1/2信號通路及記憶能力的影響
2021-09-24 02:28:28張康付燕張業廷宋文靜
中國運動醫學雜志 2021年6期
張康 付燕 張業廷 宋文靜
1 四川師范大學體育教育研究中心(成都610101)
2 北京體育大學(北京100084)
3 西南民族大學體育學院(成都610225) 4 成都大學體育學院(成都610106)
帕金森病(Parkinson’s disease,PD)是繼阿爾茨海默氏癥(Alzheimer’s dementia,AD)之后最常見的神經退行性疾病。其主要病理特征為黑質紋狀體多巴胺能神經元進行性死亡,導致多巴胺耗竭和運動障礙[1]。除運動障礙外,認知、嗅覺、抑郁以及視幻覺等非運動癥狀在PD 發病過程中也較常見。這些非運動癥狀往往在未表現出明顯運動障礙之前已經出現,如:認知功能下降,尤其是海馬依賴性記憶能力下降[2]。
多巴胺能神經元主要分布于中腦黑質、腹側被蓋區、下丘腦以及各腦室周圍。其中,黑質多巴胺能神經元可以通過中腦邊緣系統途徑投射到海馬區,表明黑質多巴胺分泌不足可能會影響海馬依賴性記憶能力[3]。基于影像學的研究發現,利用海馬皮質厚度和海馬體積鑒別帕金森癡呆(Parkinson disease with demen?tia,PDD)的準確率高達80%[4]。因此,黑質及其神經纖維支配的海馬區可能是PD 認知功能下降的基礎。目前,為了更深入研究PD,研究者通過1-甲基-4-苯基-1,2,3,6-四氫吡啶(1-methyl-4-phenyl- 1,2,3,6-tet?rahydropyridine,MPTP)染毒的方式成功模擬了PD 的發病特征,現已被廣泛應用于PD嚙齒類動物模型的建立中[5]。許多研究者也證實了MPTP可以通過損傷黑質多巴胺能神經元引起PD 癥狀,并且在MPTP 模型中通常可以觀察到與PD 相似的海馬依賴性記憶等認知功能明顯受損[6]。
細胞外信號調節激酶(extracellular signal-regulat?ed kinase,ERK)是多巴胺及谷氨酸N-甲基-D-天門冬氨酸受體(N-Metllyl-D-Aspanate receptor,NMDAR)通路中共同的下游靶點,也是海馬依賴性記憶能力中不可或缺的信號分子之一[7,8]。體內的腦源性神經營養因子(brain derived neurotrophic factor,BDNF)與受體酪氨酸激酶B(Tropomyosin receptor kinase B,TrkB)結合可以激活ERK1/2 及其下游的環磷腺苷效應元件結合蛋白(cAMP- response element binding protein,CREB),保護神經元存活、神經遞質釋放以及突觸可塑性,調節海馬依賴性記憶能力[9]。此外,多巴胺(dopa?mine,DA)與多巴胺D2 受體(dopamine 2 receptor,D2R)結合,可以直接上調ERK1/2蛋白表達,促進細胞增殖[8]。目前,臨床研究中已經發現PD 患者黑質中存在較密集的磷酸化ERK1/2[10]。改善黑質和海馬區ERK1/2 信號通路中磷酸化蛋白和總蛋白表達都可能是預防和延緩PD記憶能力下降的重要途徑。
PD記憶能力下降往往在治療過程中被低估,雖然記憶能力下降是PD迫切需要討論和解決的重要問題,但迄今為止,很少有研究從整體上解決PD患者的記憶問題。運動可以改善多種神經退行性疾病引起的神經元受損,在神經發生、神經元存活以及神經可塑性中發揮著顯著效用。已有研究者嘗試從不同角度解釋體育鍛煉對多巴胺耗竭引起的海馬神經發生以及海馬依賴性記憶能力產生積極效應的原因,但黑質和海馬CA1區ERK1/2信號通路在運動改善PD記憶能力中的作用尚未被研究。本研究主要通過觀察運動干預后小鼠記憶能力、突觸可塑性蛋白和黑質、海馬CA1 區ERK1/2信號通路變化,探討運動預防和改善PD記憶能力的可能生物學機制;同時,通過阻斷ERK1/2 分子觀察黑質和海馬CA1區ERK1/2信號通路在運動改善PD記憶能力中的作用。
1 對象與方法
1.1 實驗對象及分組
36 只正常健康雌性C57BL/6 小鼠,體重19.02 ±0.98 g,于SPF級動物實驗室分6籠飼養,每籠6只。自然喂養7天后,隨機分為6組:生理鹽水對照組(SC組)、PD 模型組(PD 組)、生理鹽水+運動組(SE 組)、PD+運動組(PDE組)、PD+運動+二甲基亞砜(dimethyl sulf?oxide,DMSO)組(PDED 組)和PD+運動+PD98059 組(PDEP 組),每組6只。整個實驗過程保持飼養環境干凈整潔,通氣良好,飼養溫度保持在23℃± 2℃,濕度保持在40%~60%,每日保持12小時光照/黑暗循環。
1.2 實驗方案及模型評價
實驗流程見圖1。
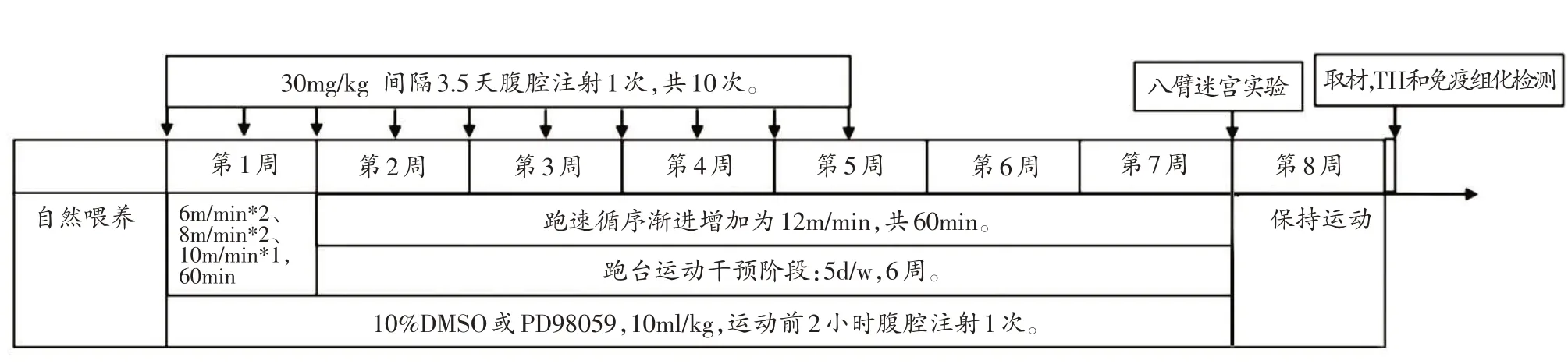
圖1 實驗流程
1.2.1 PD模型構建
PD、PDE、PDED和PDEP組小鼠按照30 mg/kg[11]的劑量每隔3.5 天腹腔注射MPTP 溶液1 次,2 次/周,共5周。SC 和SE 組在相同的時間段腹腔注射30 mg/kg 劑量的生理鹽水,2次/周,共5周。
1.2.2 跑臺運動方案
SE、PDE、PDED 和PDEP 組小鼠進行跑臺運動干預,跑臺運動于MPTP 注射第一天開始,每天19:00~20:00 固定時間進行。自然喂養結束后,第1 周為適應跑臺階段,采用較低運動強度進行。其中,第1~2 天跑速為6 m/min,第3~4 天跑速為8 m/min,第5 天為10 m/min,每天運動時間均為60 min。第2 周循序漸進增加跑臺速度,最終跑速設定為12 m/min[12],運動60 min/天,共5天/周,持續6周。八臂迷宮實驗期間小鼠持續運動1 周;整個干預過程中跑臺坡度始終設置為0°。
1.2.3 ERK1/2信號阻斷
PDEP 組按照10 ml/kg 體重腹腔注射PD98059 溶液阻斷ERK1/2信號分子[13,14],PDED組腹腔注射同等劑量10% DMSO 溶液作為對照。PD98059和DMSO 的注射時間選擇每次運動干預前2小時進行。
1.2.4 PD模型評價
PD 模型評價標準:運動干預結束后,通過震顫麻痹評分量表將小鼠的震顫麻痹分為0~4等級,0級:無任何癥狀;1級:出現弓背、豎毛、間歇性小震顫,但活動自如;2級:吞咽頻繁,頻繁性震顫,后肢張開,活動逐漸受限;3 級:出現流涎、持續性震顫,四肢僵硬,活動受限;4 級,因全身麻痹而死亡[15]。通過黑質酪氨酸羥化酶(tyrosine hydroxylase,TH)染色判斷PD 模型小鼠黑質多巴胺能神經元的功能受損情況。
1.3 測試方法、指標
1.3.1 八臂迷宮測試小鼠記憶能力
八臂迷宮是用來評價藥物或大腦受損狀態下學習和記憶表現的一種有效方式。八臂迷宮實驗包括訓練和測試兩個階段。訓練階段:禁食12 小時,保持體重在80%~85%之間。迷宮中央區和各臂均放置3~4粒直徑約為2 mm 的圓形食物,將3~4 只小鼠放入中央區30 s后,打開各臂通道自由活動10 min,2次/天。之后,將每只小鼠單獨放入迷宮,在1、3、5、7號臂食盒的近端放一粒食物,小鼠吃盡食物或停留10 min 取出小鼠,2次/天。
測試階段:1、3、5、7 號臂中放入一粒直徑約為2 mm的圓形食物,將小鼠放入中央區30 s后自由活動并尋找食物10 min,當吃盡各臂食物或10 min之內未吃完食物,系統均自動終止實驗。計算機自動輸出檢測指標:①工作記憶錯誤:同一次訓練中小鼠重復進入已經吃過食物的臂;②工作記憶錯誤率:工作記憶錯誤與總的入臂次數之比;③參考記憶錯誤:小鼠進入不曾放過食物的臂;④參考記憶錯誤率:參考記憶錯誤與總的入臂次數之比。
1.3.2 取材
行為學測試結束后,斷頸處死。取小鼠完整腦組織,放入液氮速凍,-80℃冰箱保存,待制作黑質、海馬部位的切片。根據腦立體定位圖譜,黑質切片選取前囟-4.8 mm 到-6.3 mm,海馬CA1 區選取約前囟-2.12 mm,切片約10 mm。
1.3.3 TH免疫組化染色
利用免疫組化染色觀察小鼠腦組織切片中黑質TH陽性細胞的變化。腦組織切片脫蠟后放入染色缸,3%甲醛雙氧水室溫10 min;0.1%磷酸緩沖鹽(PBS)清洗切片3 次,5 min/次。洗凈的切片放入0.01 M 檸檬酸鹽緩沖液(pH 6.0),加熱直至沸騰5 min,重復1次。PBS清洗3×5 min/次,滴加10%山羊血清封閉液封20 min,滴加TH一抗(濃度1︰800),4℃孵育過夜。滴加生物素化山羊抗兔二抗,37℃孵化30 min;0.1%PBS清洗3×5 min/次;DAB顯色、復染;中性樹膠封片,自然晾干。將載玻片放入蔡司光學顯微鏡上進行掃描成像,并使用MBF生物科學數字顯微鏡攝像機(Willis?ton,VT)在計算機上進行成像;在100倍放大倍數下,通過人工計數每張拍照圖像來統計TH細胞的數量。
1.3.4 免疫熒光染色
利用免疫熒光觀察小鼠腦組織切片中黑質和海馬CA1 區中ERK1/2、CREB、突觸素(synaptophysin,SYN)和突觸后致密蛋白95(postsynaptic density-95,PSD-95)PSD-95 蛋白表達。脫蠟至水;0.01 M 檸檬酸鹽緩沖液(pH=6.0)中加熱至沸騰,重復1 次;PBS 清洗5 min/次,反復清洗3次。37℃室溫下使用10%山羊血清封閉30 min;滴加一定比例ERK1/2、CREB、SYN 和PSD-95 一抗(濃度:1︰50、1︰50、1︰400 和1︰100),4℃過夜。PBS 清洗3×5 min/次;滴加二抗,37℃孵育30 min。PBS 清洗切片3×5 min/次;滴加4’,6-二脒基-2-苯基吲哚(4’,6-Diamidino-2-Phenylindole,Di?hydrochloride,DAPI)染液,37℃孵育30 min;PBS 清洗切片3×5 min/次;用抗熒光衰減封片劑封片。采用免疫熒光顯微鏡觀察并采集黑質及海馬區的圖像并分析結果;使用image J 軟件測量平均熒光強度值,對黑質和海馬神經元蛋白表達進行半定量分析。特定區域熒光強度公式為:平均熒光強度=該區域熒光強度總和/該區域面積。
1.4 主要實驗試劑與儀器
MPTP(proteintech 公司),Anti-Tyrosine Hydroxy?lase 抗體(abcam 公司),ERK1/2 Antibody(proteintech公司),Anti-SYN Antibody(abcam公司),PSD-95 Anti?body(proteintech 公司),CERB Antibody(proteintech 公司),β-actin 兔克隆抗體(abcam 公司),冰凍切片機(Leica 公司),蓋玻片(江蘇世泰實驗器材有限公司),載玻片(江蘇世泰實驗器材有限公司),熒光顯微鏡(德國Leica公司)等。
1.5 統計學分析
采用SPSS19.0對各組小鼠行為學和分子生物學數據進行分析,結果均采用均值±標準差(±s)表示,組間采用多因素方差分析(ANOVA)法分析不同指標的差異性,采用最小顯著性差異(LSD)法對各指標進行多重比較檢驗,P<0.05為差異具有統計學意義。
2 結果
2.1 造模后PD小鼠黑質多巴胺能神經元及行為學比較
通過震顫麻痹評分表對PD組小鼠進行評級發現,PD 小鼠成模率約83.33%。具體表現為,約66.67%的小鼠表現出第二等級的典型特征:豎毛、豎尾、流涎、持續震顫、四肢僵硬;16.67%的小鼠表現出第一等級特征:豎毛、間歇性小震顫和弓背。從黑質TH 染色結果中發現,與SC組相比,PD組黑質TH陽性細胞數量平均減少約45.75%(8.23 ± 1.21 vs.17.99 ± 0.95,P<0.01),SE組和PDE組小鼠黑質TH陽性細胞數量比SC組和PD 組小鼠明顯增加(26.31 ± 2.14 vs.17.99 ±0.95,13.85 ± 1.27 vs.8.23 ± 1.21,P<0.05)。見圖2。
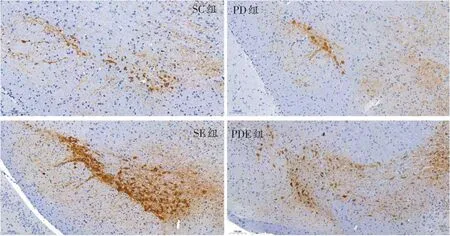
圖2 SC、PD、SE和PDE組小鼠黑質TH染色(標尺:100μm)
2.2 八臂迷宮測試中各組小鼠記憶能力比較
由表1 可知,與SC 組相比,PD 組小鼠工作記憶錯誤次數和工作記憶錯誤率顯著升高(P<0.05);SE 組小鼠工作記憶錯誤次數和工作記憶錯誤率與SC 組相比僅表現出下降的趨勢。與PD組小鼠相比,PDE組小鼠工作記憶錯誤次數和工作記憶錯誤率均顯著降低(P<0.01);與PDE組相比,PDED組小鼠工作記憶錯誤次數和工作記憶錯誤率均未表現出顯著差異。PDEP 組小鼠與PDED 組小鼠之間工作記憶錯誤次數和工作記憶錯誤率也未顯示顯著差異,但從數據來看,PDEP 小鼠工作記憶錯誤率平均增加幅度更大。
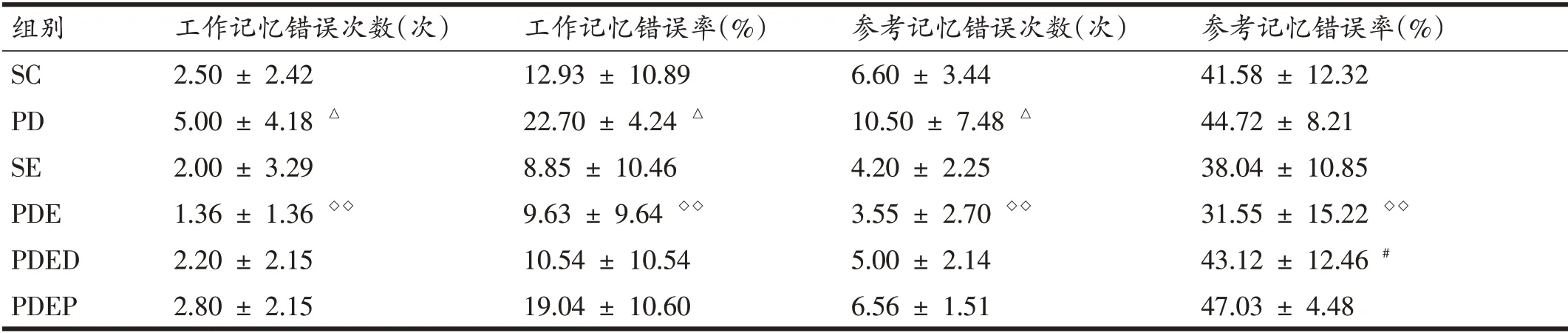
表1 八臂迷宮測試中各組小鼠記憶能力比較(n=6)
PD 組小鼠參考記憶錯誤次數顯著高于SC 組(P<0.05);參考記憶錯誤率未表現出顯著變化。PDE 組小鼠參考記憶錯誤次數和參考記憶錯誤率比PD 組均顯著降低(P<0.05);與SC 組相比,SE 組小鼠參考記憶錯誤次數和參考記憶錯誤率均表現出下降的趨勢,但不顯著。PDED 組小鼠參考記憶錯誤率明顯高于PDE 組(P<0.05),PDED組與PDEP組之間未表現出顯著差異;但從數據來看,PDEP組小鼠參考記憶錯誤率和參考記憶錯誤次數均存在增加的趨勢。
2.3 各組小鼠黑質、海馬CA1 區SYN 和PSD-95 蛋白表達比較
如圖3、圖4、表2所示,與SC 組相比,PD 組小鼠黑質SYN和PSD-95染色面積明顯減少,但蛋白表達僅表現出了下降的趨勢。與PD 組相比,PDE 組小鼠黑質SYN和PSD-95染色面積增加,但蛋白表達未表現出顯著變化,僅表現出升高的趨勢。與PDE 組相比,PDED組小鼠黑質SYN 和PSD-95 染色面積及蛋白表達均無顯著差異。與PDED組相比,PDEP組小鼠黑質PSD-95蛋白表達顯著下降(P<0.01)。
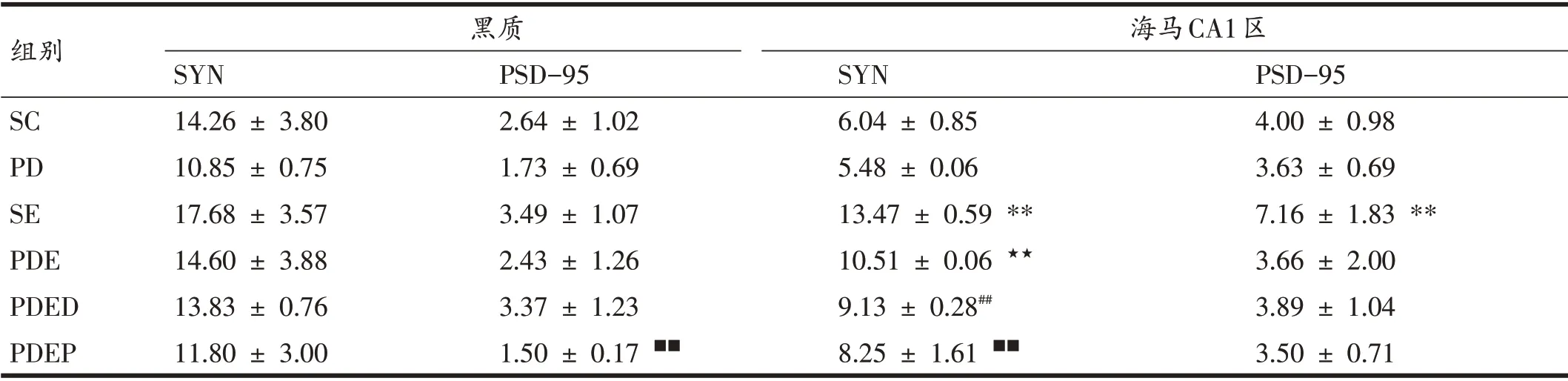
表2 各組小鼠黑質、海馬CA1區SYN和PSD-95蛋白表達比較(n=6)
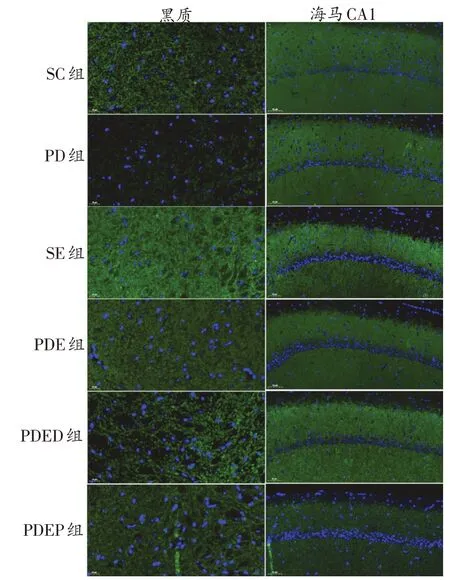
圖3 各組小鼠黑質、海馬CA1區SYN蛋白表達比較(綠色為SYN染色,藍色為DAPI染色)
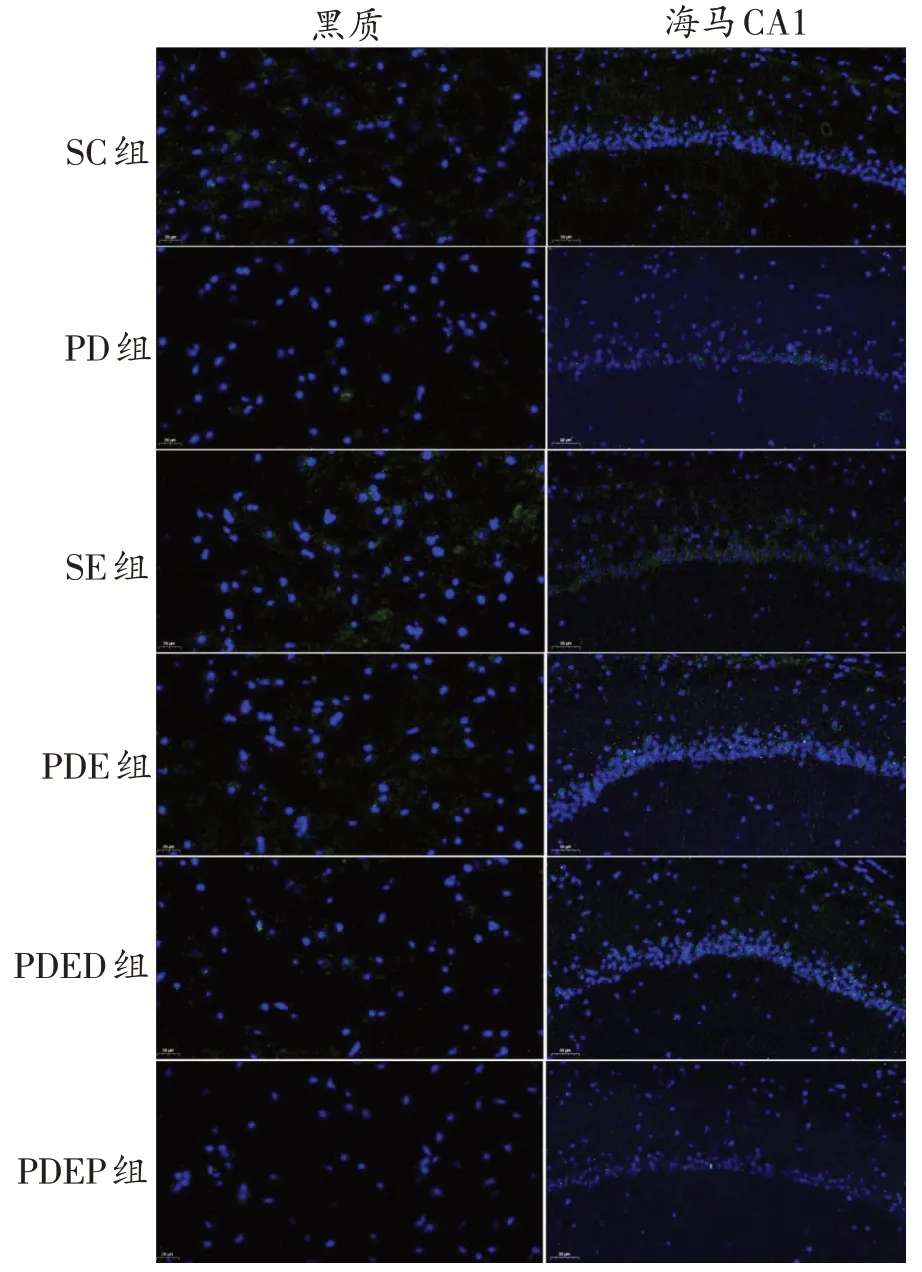
圖4 各組小鼠黑質、海馬CA1區PSD-95蛋白表達比較(綠色為PSD-95染色,藍色為DAPI染色)
與SC 組比,PD 組小鼠海馬CA1 區SYN 和PSD-95染色面積明顯減少,但蛋白表達僅表現出了下降的趨勢;SE 組小鼠海馬CA1 區SYN 和PSD-95 染色面積明顯增加,蛋白表達顯著升高(P<0.01)。與PD 組相比,PDE 組小鼠SYN 染色面積明顯增加,蛋白表達顯著升高(P<0.01),PSD-95未表現出顯著變化。與PDE組相比,PDED小鼠海馬CA1區SYN蛋白表達顯著降低(P<0.01)。與PDED組相比,PDEP組SYN蛋白表達顯著降低(P<0.01),PSD-95未顯示出顯著差異。
2.4 各組小鼠黑質、海馬CA1 區ERK1/2、CREB 蛋白表達比較
如圖5、圖6、表3所示,與SC 組相比,PD 組小鼠黑質ERK1/2 和CREB 免疫熒光染色面積均明顯減少,蛋白表達均顯著降低;SE組小鼠黑質ERK1/2免疫熒光染色面積明顯增加,蛋白表達顯著升高(P<0.01)。與PD組相比,PDE 組黑質ERK1/2 和CREB 免疫熒光染色面積均明顯增加,蛋白表達顯著升高(P<0.01)。與PDE組相比,PDED 組小鼠黑質ERK1/2 和CREB 免疫熒光染色面積和蛋白表達均未表現出顯著變化。PDEP 組比PDED 組黑質ERK1/2 和CREB 蛋白表達顯著降低(P<0.01和P<0.05)。
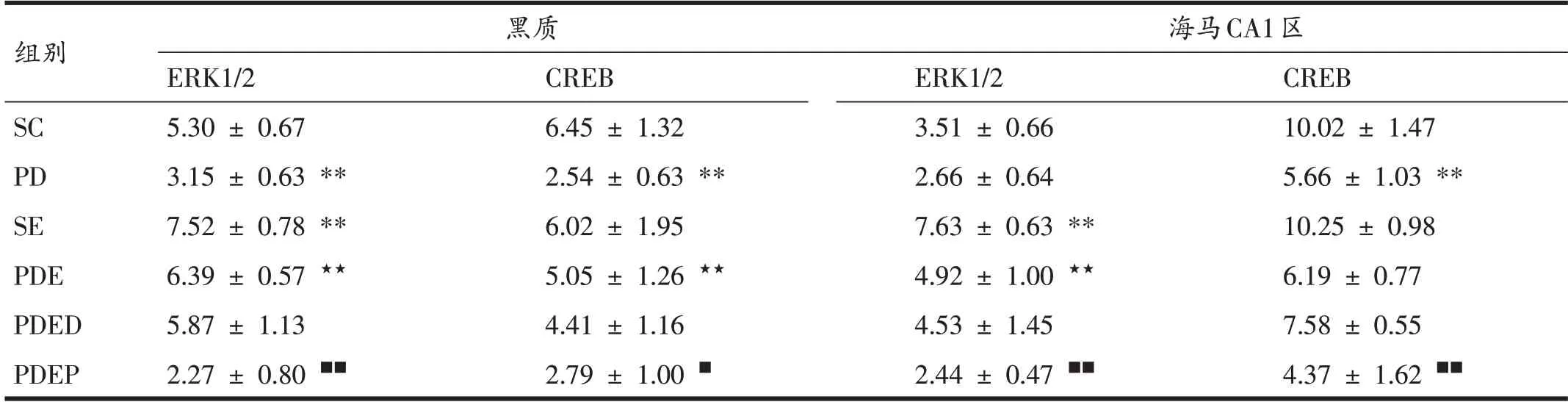
表3 各組小鼠黑質、海馬CA1區ERK1/2信號通路中的相關蛋白表達比較(n=6)
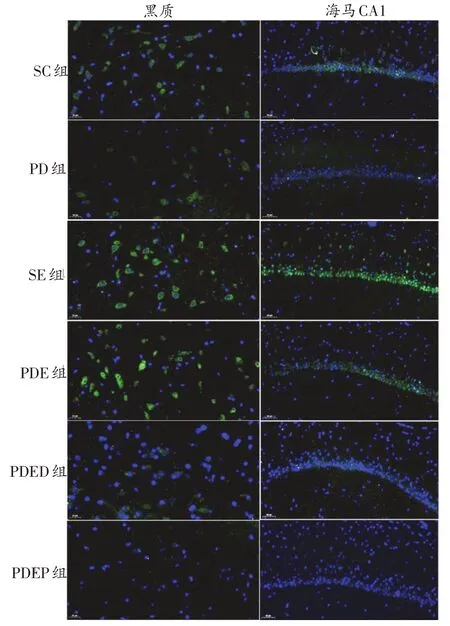
圖5 各組小鼠黑質、海馬CA1區ERK1/2蛋白表達比較(綠色為ERK1/2染色,藍色為DAPI染色)
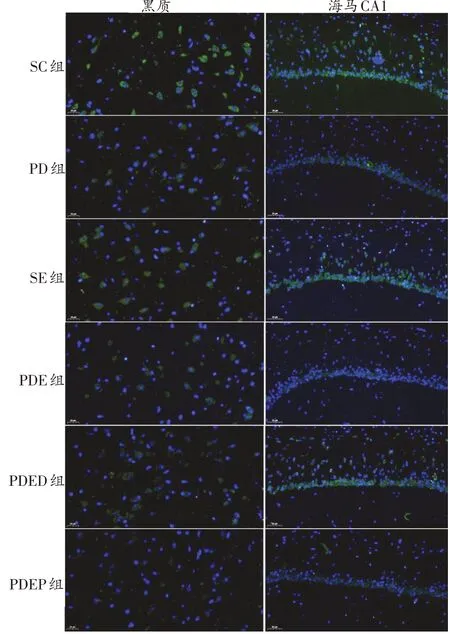
圖6 各組小鼠黑質、海馬CA1區CREB蛋白表達比較(綠色為CREB染色,藍色為DAPI染色)
與SC組相比,PD組小鼠海馬CA1區CREB免疫熒光染色面積明顯減少,蛋白表達顯著減少(P<0.01),ERK1/2 僅表現出下降的趨勢;SE 組小鼠海馬CA1 區ERK1/2免疫熒光染色面積明顯增加,蛋白表達顯著升高(P<0.01)。與PD 組相比,PDE 組小鼠海馬CA1 區ERK1/2免疫熒光染色面積明顯增加,蛋白表達顯著升高(P<0.01);CREB 變化不顯著,僅表現出升高的趨勢。與PDED 組相比,PDEP 組小鼠CA1 區ERK1/2 和CREB免疫熒光染色面積明顯減少,蛋白表達顯著減少(P<0.01);PDED組與PDE組之間無顯著差異。
3 討論
TH 是兒茶酚胺(catecholamine,CA)(多巴胺、去甲腎上腺素和腎上腺素等)生物合成的起始和限速酶[16],參與了PD等多種CA疾病的發病機制,對于依賴于CA細胞活性的神經元功能至關重要。通過TH染色觀察黑質多巴胺能神經元的完整性是目前判斷PD 病理進程的“金標準”[17]。目前,睪丸激素已被認為是導致PD 的潛在危險因素,因此本研究采用與Lu[17]、Gong[18]、Torres[19]等研究相同的雌性小鼠作為研究對象。本研究結果顯示,正常小鼠黑質TH陽性細胞較多,MPTP處理的小鼠黑質TH 陽性細胞計數明顯降低,說明MPTP染毒誘導小鼠產生了與PD 相似的多巴胺能神經元功能異常。以往研究指出,MPTP處理后小鼠黑質致密部TH 減少54.0% ± 4.3%[20]。Churchill 等[21]也發現MPTP處理的小鼠黑質致密部TH 計數下降50%,4 周后下降55%,均與本研究結果相近。本研究中經過跑臺運動的PD 模型小鼠黑質TH 陽性細胞計數明顯高于PD 模型組,說明跑臺運動可以有效抑制多巴胺能神經元功能受損,延緩PD的發展速度。基于人體或動物實驗的研究均發現,運動可以有效緩解PD導致的運動行為障礙和黑質TH活性降低或數量減少。Cugusi等[22]研究發現,跑步機運動可以顯著改善PD 患者的步行距離、下肢肌肉力量、平衡能力、活動安全性、抑郁和冷漠等癥狀,揭示了跑步機運動在改善PD患者運動癥狀和非運動癥狀方面的有效性。Koo 等[23]究發現跑臺運動能夠促進PD 小鼠黑質TH 表達,抑制多巴胺能神經元的丟失。因此,運動在延緩或改善PD 病程中具有重要價值,可以作為PD治療中的一種重要的輔助手段。
此外,本研究結果還顯示PD組小鼠的工作記憶錯誤次數、工作記憶錯誤率和參考記憶錯誤次數明顯高于生理鹽水對照組,運動干預的PD組小鼠工作記憶錯誤次數、工作記憶錯誤率、參考記憶錯誤次數以及參考記憶錯誤率比PD 組小鼠明顯降低,說明PD 小鼠表現出了明顯的記憶能力減退,跑臺運動可以有效改善PD小鼠和正常小鼠的記憶能力。這一結果與許多國外臨床研究的結論相同。如:Bello等[24]研究證實,跑步機運動能夠有效提高PD患者的認知任務成績。Cruise等[25]發現有氧運動可以選擇性改善PD 患者的認知功能,如:工作記憶和執行功能。現在越來越多的證據表明,記憶能力與突觸可塑性有關,突觸功能減退或突觸相關蛋白表達下降是早期神經退行性疾病常見的病理特征[26]。SYN 和PSD-95 蛋白是神經元突觸的一種特異性突觸前標志蛋白和突觸后致密蛋白,參與突觸生長、發育以及重塑,在各種記憶中發揮著不可或缺的作用[17,27,28]。一些基于PD 動物模型的研究發現,PD 小鼠海馬突觸結構、功能可塑性受損和突觸標志蛋白表達失調。Qu等[29]在MPTP 誘導的PD小鼠中發現海馬DG區和CA3區苔蘚纖維強度明顯降低,突觸可塑性受損,認為其機制可能涉及到MPTP 下調了SYN 和PSD-95 等的表達,以及降低BDNF-TrkB 通路的活性。本研究結果發現,PD 小鼠黑質和海馬CA1 區SYN 和PSD-95 表達均表現出了降低的趨勢,跑臺運動明顯增強了海馬CA1區SYN蛋白表達,同時,黑質SYN、PSD-95蛋白表達存在上調的趨勢,說明跑臺運動具有增強PD小鼠黑質和海馬CA1區突觸可塑性的潛力,最終導致PD小鼠記憶能力得到改善。雖然運動干預的正常小鼠工作記憶和參考記憶錯誤次/率僅表現出了下降的趨勢,但海馬CA1 區SYN 和PSD-95 蛋白表達均顯著升高。本研究考慮到PD 的運動障礙,選擇較小運動強度干預,因此,可能較小強度運動引起的正常小鼠突觸可塑性的改變程度未能達到記憶能力改善的閾值,新生成的神經元突觸或者增強的突觸功能達不到改善記憶能力的效果,后期將對不同運動強度的干預效果進行進一步研究。Klein[2]在研究中也發現運動可以改善MPTP 小鼠Morris 水迷宮測試的認知功能,但隨著蛋白或基因表達的升高將出現天花板效應,升高到一定水平后,運動不再有助于認知行為的進一步改善,該研究與本研究結果相似。以往關于PD黑質、海馬突觸可塑性和認知功能機制的研究指出,PD記憶能力和突觸可塑性的改變與BDNF 表達上調有關。Hirsch 等[30]系統回顧了2017年6月之前收錄在MEDLINE、EMBASE、Cochrane Library、PsycINFO 和PubMed 等數據庫中關于運動與PD 患者血清腦源性神經營養因子(brain-derived neu?rotrophic factor,BDNF)的研究,其中兩項隨機對照試驗和四項預實驗研究提供了運動療法增加PD 患者血清BDNF的直接證據。基于MPTP小鼠的文獻均證實,振動和跑臺運動均能夠上調PD 小鼠腦組織BDNF 表達,預防多巴胺能神經元功能受損,并提出運動時間越長對細胞水平的增益效果越明顯[31,32]。因此,本研究認為BDNF及其下游因子的改變可能是運動改善PD記憶能力的重要原因。
ERK1/2 信號通路是BDNF 下游調控神經元存活、神經遞質釋放以及突觸可塑性的重要途徑,也是目前研究最多的信號通路之一。越來越多的證據表明,PD與ERK1/2 活性密切相關,甚至一些研究者將PD 歸因于ERK 基因表達的依賴性變化[33]。靜息狀態下,ERK位于細胞質中,當ERK被激活時可以迅速磷酸化,進入細胞核調控其他轉錄因子的活性,影響SYN和PSD-95等特異性蛋白表達,引起細胞能量代謝、生理生化功能改變,進而發揮維持神經元發育和突觸可塑性穩態的作用[34]。Liang等[35]觀察MPTP誘導的亞急性PD模型小鼠發現,中腦p-ERK1/2/ERK1/2表達顯著降低,治療后隨著PD 癥狀的改善,p-ERK1/2/ERK1/2 表達明顯升高。但也有研究得出不同的結論,Siracusa 等[36]觀察MPTP誘導的亞急性PD模型小鼠發現,PD模型小鼠多巴胺能神經元p-ERK1/2 蛋白表達明顯增強,認為PD癥狀的改善與p-ERK1/2 蛋白表達下降有關。ERK1/2磷酸化水平的改變涉及p-ERK1/2 蛋白的改變和ERK1/2蛋白的改變。我們推測,PD腦組織中p-ERK1/2/ERK1/2 的改變不僅僅是p-ERK1/2 發生了改變,ERK1/2信號通路中相關因子的改變在改善PD癥狀中也發揮著關鍵作用。以往較少有研究從ERK1/2 通路入手探討ERK1/2 信號通路中相關因子表達的改變是否是PD 認知癥狀改變的潛在原因。本研究針對ERK1/2表達進行檢測、分析,研究結果顯示,與生理鹽水對照組相比,PD 模型小鼠黑質和海馬CA1 區CREB蛋白表達和黑質ERK1/2蛋白表達均顯著降低,跑臺運動的PD模型小鼠黑質ERK1/2、CREB以及海馬CA1區ERK1/2蛋白表達明顯升高,說明運動有效地增強了PD模型小鼠黑質和海馬CA1 區ERK1/2 信號通路中相關因子表達。最近的一項研究顯示,ERK1/2-CREB信號通路參與促進海馬神經元樹突棘絲狀偽足、蘑菇狀突起及谷氨酸能興奮性突觸的形成[37]。阻斷ERK1/2 信號可以減少新的樹突棘和偽足的形成,抑制長時程增強(long-term potentiation,LTP)[38,39]。因此,PD 模型小鼠記憶能力和突觸可塑性的改善可能是運動上調了黑質和海馬CA1區ERK1/2信號通路中相關因子表達。
為了進一步證實ERK1/2 信號通路在跑臺運動改善PD小鼠記憶能力中的作用,本研究采用PD98059抑制ERK1/2 分子觀察PD 小鼠記憶能力以及黑質、海馬CA1 區SYN 和PSD-95 蛋白變化,結果顯示ERK1/2 分子阻斷后,跑臺運動干預的PD小鼠黑質和海馬CA1區SYN或PSD-95蛋白表達顯著降低,工作記憶和參考記憶錯誤率存在降低的趨勢;注射10%DMSO溶液的小鼠與僅進行運動干預的PD 小鼠相比無顯著差異,海馬CA1區SYN蛋白下調,說明10%DMSO溶液對小鼠記憶能力結果的影響較小,ERK1/2信號通路在跑臺運動改善PD 小鼠黑質和海馬CA1 區突觸可塑性和記憶能力中發揮著關鍵作用,進一步證實跑臺運動可以通過上調ERK1/2信號通路中相關因子表達,改善黑質和海馬CA1區突觸可塑性,增強PD小鼠記憶能力。ERK1/2分子阻斷后小鼠參考記憶和工作記憶僅表現出了增強的趨勢。可能是由于PD是一種多系統的退行性疾病,除黑質和海馬投射路徑外,額葉ERK1/2蛋白表達異常等也是導致PD認知功能受損的關鍵原因。Luo等[40]也發現ERK激酶抑制劑PD98095定點注射前額葉內可以顯著抑制ERK1/2 和CREB 標記細胞數量,并嚴重損害大鼠長期回避恐懼記憶的恢復。因此,推測PD小鼠記憶能力的下降與改善可能也與額葉功能有關,且記憶能力受損的不同階段可能涉及到不同的神經機制,對此,尚需進一步研究證實。
4 結論
慢性MPTP 染毒誘導的PD 小鼠具備了PD 典型的黑質多巴胺能神經元功能受損,表現出工作記憶和參考記憶減退。跑臺運動可以通過調節ERK1/2 信號通路,改善黑質和海馬CA1 區突觸可塑性,增強PD 小鼠記憶能力。
猜你喜歡
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
學苑創造·A版(2020年9期)2020-10-13 09:41:02
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
作文周刊·小學一年級版(2016年27期)2017-06-03 23:21:17
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
新湘評論·下半月(2016年4期)2016-05-05 22:12:41
海外文摘(2016年4期)2016-04-15 22:28:55
作文與考試·小學高年級版(2015年17期)2015-05-30 10:48:04
