油茶果生刺盤孢液泡分選蛋白CfVps26的功能*
2021-10-09 05:49:46李茜雅張盛培
林業科學 2021年8期
李茜雅 張盛培 李 河
(經濟林培育與保護省部共建教育部重點實驗室 中南林業科技大學 長沙 410004)
油茶(Camelliaoleifera)樹不僅可以保持水土、涵養水源,而且還能提供優質的食用油,具有極高的生態、經濟和社會價值(劉躍進等, 2007; 陳永忠等, 2013)。由刺盤孢屬(Colletotrichumspp.)真菌導致的炭疽病能造成油茶落葉、落花和落果(靳愛仙等, 2009)。油茶籽產量因炭疽病減產10%~30%,重病區甚至減產50%以上,造成巨大的經濟損失(徐麗萍等, 2015)。目前,我國防治油茶炭疽病主要采用苯并咪唑類(多菌靈或甲基托布津等)內吸性殺菌劑(鄭少華, 2012),但由于其作用位點和機制較為單一,容易使病原菌產生抗藥性。而病原菌對多靶標位點的殺菌劑不易產生抗藥性(詹家綏等, 2014),因此亟需研究篩選出更多新的藥物靶標位點,為新型殺菌劑的研發提供依據。
Retromer復合體主要介導跨膜蛋白從內涵體到反式高爾基體的運輸和從內涵體到細胞膜的運輸,使得蛋白受體可以反復循環利用,避免運輸到溶酶體被降解,維持細胞內的代謝平衡(Bonifacinoetal., 2008; Hierroetal., 2007)。該復合體由液泡分選蛋白(vacuolar protein sorting,簡稱vps)組成,分為由Vps26、Vps29和 Vps35構成的貨物識別三聚體和由Vps5和Vps17構成的微管分選蛋白二聚體(Cullenetal., 2011)。禾谷鐮刀菌(Fusariumgraminearum)Retromer復合體Vps5、Vps17、Vps26、Vps29和 Vps35五個分選蛋白均參與調控菌絲生長、產孢和致病過程(張瑩, 2011)。在稻瘟病菌(Magnaporthegrisea)Vps26、Vps29和 Vps35組成的貨物識別三聚體均影響該病菌的產孢量和致病力(鄭文輝, 2014)。
果生刺盤孢(Colletotrichumfructicola)是油茶炭疽病優勢致病菌(李河等, 2014; 2017; 2018; 2019)。筆者課題組從果生刺盤孢全基因組中鑒定到1個與釀酒酵母(Saccharomycescerevisiae)的Retromer復合體亞基Vps26的同源蛋白,命名為CfVps26,但其生物學功能尚不清楚。本文擬以液泡分選蛋白Vps26為研究對象,其在油茶果生刺盤孢中的生物學功能,為探究Vps26的作用機制和防治油茶炭疽病提供參考。
1 材料與方法
1.1 供試菌株
果生刺盤孢CFLH16為野生型菌株,突變體ΔCfvps26和回補菌株ΔCfvps26/CfVPS26由本試驗獲得。
1.2 CfVps26蛋白系統發育分析
根據釀酒酵母中Vps26蛋白氨基酸序列(XP_031879784.1)在果生刺盤孢全基因組數據庫(ANPB00000000.1)中進行搜索,獲得與其同源的CfVps26蛋白氨基酸序列,與NCBI數據庫中其他真菌的Vps26蛋白氨基酸序列進行系統發育分析。
1.3 CfVPS26基因敲除載體構建及突變體篩選
以野生型菌株 CFLH16 基因組 DNA 為模板,設計引物CfVPS26-1F/CfVPS26-2R、CfVPS26-3F/CfVPS26-4R(表1)分別PCR擴增CfVPS26目的基因上下游各1 kb左右的DNA片段,將其作為構建敲除載體片段的上下臂。引物 Hyg F/Hyg R(表1)用來擴增潮霉素抗性基因片段(HPH)。利用引物CfVPS26-1F/CfVPS26-4R,采用Over-lap方法將上臂、潮霉素和下臂DNA片段順序連接,得到CfVPS26基因敲除載體片段。
參照Zhang等(2019)方法制備果生刺盤孢CFLH16原生質體。參考高亞蘭等(2020)的方法將CfVPS26基因敲除載體片段轉化至原生質體中。采用基因內探針引物CfVPS26-7F/CfVPS26-8R和基因外引物CfVPS26-5F/H855R(表1)驗證轉化子。基因內探針引物不能擴增出目的基因條帶,且基因外引物可擴增出正確大小條帶的轉化子為CfVPS26基因敲除突變體。

表1 CfVPS26在果生刺盤孢研究中使用的引物Tab.1 CfVPS26 primers used in Colletotrichum fructicola study
1.4 CfVPS26基因敲除突變體回補菌株的獲得
采用引物CfVPS26-9F/CfVPS26-10R PCR擴增含有啟動子在內的CfVPS26基因回補片段,參照高亞蘭等(2020)方法構建回補質粒; 將回補質粒轉化至突變體ΔCfvps26原生質體中,在添加博來霉素的TB3培養基上培養3 天,熒光篩選回補菌株。
1.5 突變體ΔCfvps26的表型測定
1.5.1 不同種類培養基上生長測定 參照姚權等(2019)方法對菌株的生長速率進行測定。
1.5.2 細胞壁敏感性測定 在野生型、突變體和回補菌株菌落邊緣用無菌打孔器(Φ=8 mm)切取菌餅,分別接種含0.01%十二烷基硫酸鈉(sodium dodecyl sulfate,SDS)和200 μg·mL-1熒光增白劑(calcofluor white,CFW)的PDA平板上,培養3天后測量菌落直徑。
1.5.3 產孢量與附著胞形成測定 將野生型、突變體和回補菌株菌落分別接種在PDB培養基中,培養4 天后統計產孢量。得到等濃度(105個·mL-1)分生孢子懸浮液,滴加10 μL 置于疏水玻片上,保濕培養12 h后觀察記錄附著胞形成率情況。
1.5.4 致病力測定 在培養3天的野生型、突變體和回補菌株菌落邊緣用無菌打孔器(Φ=8 mm)切取菌餅,將菌餅有菌絲的一面接種油茶葉片上,空白PDA培養基作為對照。保濕培養,觀察發病情況并統計直徑。
1.5.5 內質網壓力脅迫劑敏感性測定 在野生型、突變體和回補菌株菌落邊緣用經滅菌的打孔器(Φ=8 mm)切取菌餅,接種于含5 mmol·L-1二硫蘇糖醇DTT的PDA平板上,培養4 天,測量菌落直徑。
1.5.6 糖原染色 參照鄭文輝(2014)的方法對附著胞進行糖原染色,制備濃度為105個·mL-1的野生型CFLH16和突變體ΔCfvps26分生孢子懸浮液,各取10 μL滴于疏水玻片上黑暗保濕培養24 h,吸取多余水分后滴加30 μL糖原染料(I2/KI,120 mg·mL-1KI + 20 mg·mL-1I2),黑暗靜止染色30 min,顯微鏡觀察附著胞糖原代謝情況。
2 結果與分析
2.1 CfVps26的鑒定及系統進化分析
鑒定到1個與釀酒酵母中Retromer復合體亞基Vps26同源的蛋白CfVps26,對其結構域進行分析發現,從第5~279個氨基酸僅有1個Vps26結構域(圖1A)。系統發育分析表明果生刺盤孢中Vps26氨基酸序列與希金斯刺盤孢(C.higginsianum)親緣關系最近,與釀酒酵母親緣關系最遠(圖1B)。
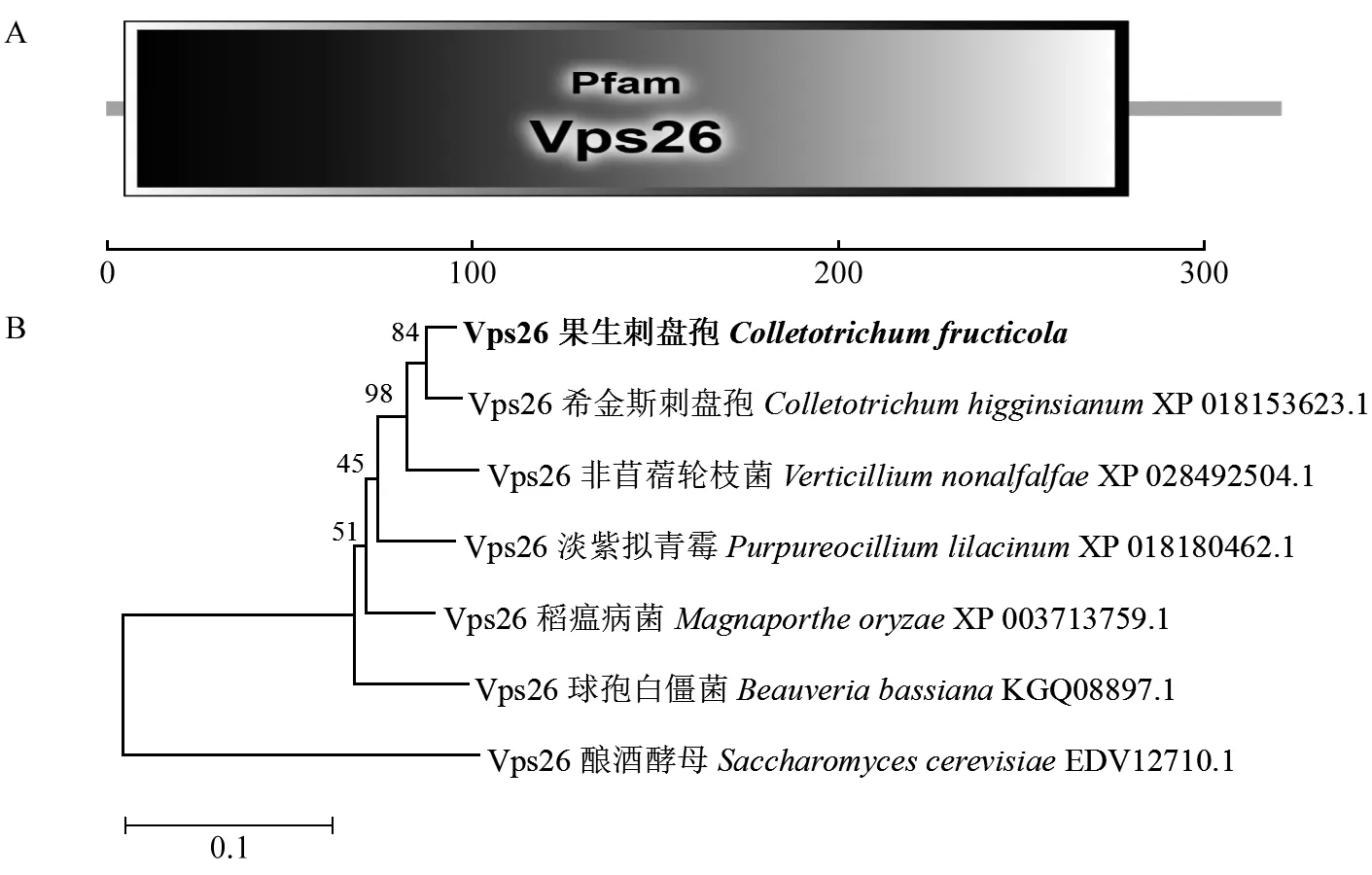
圖1 CfVps26結構域和系統發育分析Fig. 1 Domain and phylogenetic analysis of CfVps26A: CfVps26蛋白的結構域。B: 鄰近相連法構建系統發育樹。A: The structure of CfVps26; B: Phylogenetic tree was constructed by neighbor joining method.
2.2 CfVPS26基因敲除突變體及回補菌株的獲得
根據同源重組原理構建CfVPS26基因敲除策略(圖2A)。利用引物CfVPS26-7F/CfVPS26-8R和CfVPS26-5F/H855R進行驗證,獲得突變體ΔCfvps26。回補菌株ΔCfvps26/CfVPS26通過熒光篩選以及PCR驗證獲得(圖2B)。
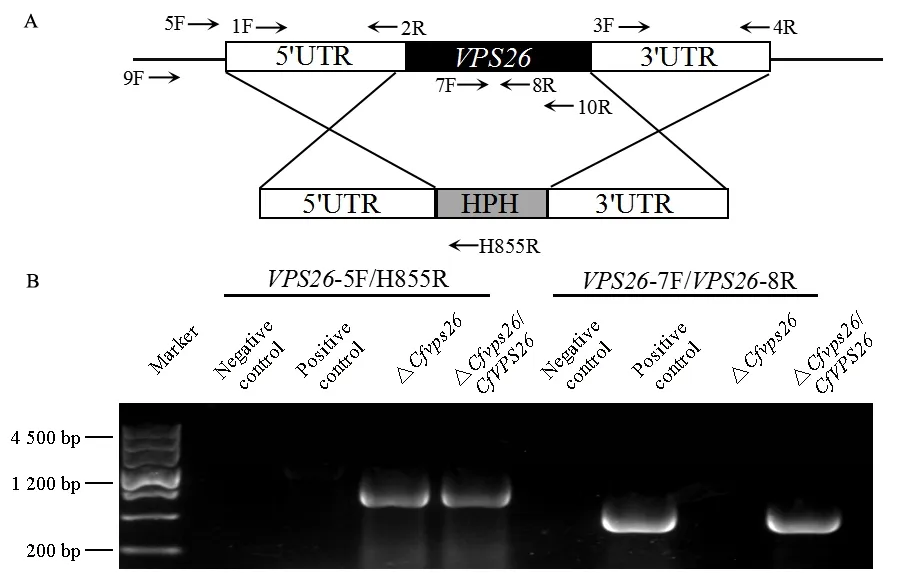
圖2 獲得CfVPS26基因敲除突變體Fig. 2 Generation of the CfVPS26 gene deletion mutantA: 基因敲除策略圖; B: 電泳驗證圖。A: Schematic of the deletion strategy; B: Electrophoregram for verification.
2.3 CfVPS26參與調控果生刺盤孢營養生長、產孢和附著胞形成
突變體ΔCfvps26在PDA和MM培養基上,菌落直徑均小于野生型及回補菌株(圖3A),統計分析差異顯著(圖3B),氣生菌絲明顯減少(圖3C),說明CfVPS26能夠調控果生刺盤孢菌體的營養生長。
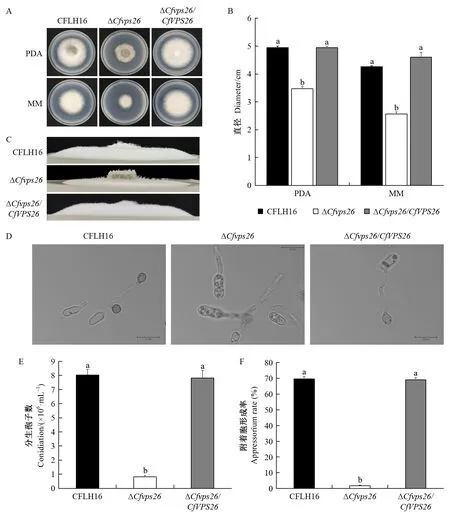
圖3 CfVPS26基因敲除突變體的菌絲生長測定和產孢量、附著胞形成率統計分析Fig. 3 Growth defect, statistical analysis of conidial quantity and appressorium formation rate of CfVPS26 gene deletion mutantA: 突變體在PDA、MM 培養基上生長情況; B: 直徑統計; C: 菌落側視圖; D: 突變體ΔCfvps26能形成少量附著胞; E:產孢量統計; F:附著胞形成率。 不同小寫字母表示不同菌株間差異極顯著(P≤0.01)。下同。A: Growth of ΔCfvps26 strain inoculated on PDA and MM media; B: Diameter; C: Lateral views; D: The ΔCfvps26 can form a small amount appressorium; E: Conidiation; F: Appressorium formation rate. Different small letters indicate significan difference between different strains(P≤0.01). The same below.
對野生型菌株CFLH16、突變體ΔCfvps26以及回補菌株ΔCfvps26/CfVPS26的產孢量進行統計分析,發現突變體菌株ΔCfvps26產孢量顯著下降,只有野生型和回補菌株產孢量的10%左右(圖3E)。附著胞是病原真菌侵染植物組織的關鍵結構。對野生型菌株CFLH16、突變體ΔCfvps26以及回補菌株ΔCfvps26/CfVPS26的附著胞形成率統計分析發現,與野生型和回補菌株相比,突變體ΔCfvps26附著胞形成率顯著下降(圖3D),且差異顯著(圖3F)。以上結果說明CfVPS26基因參與調控果生刺盤孢分生孢子及附著胞的形成,這可能是突變體ΔCfvps26對油茶葉片致病力顯著降低的主要原因。
2.4 CfVPS26參與調控果生刺盤孢對外界環境脅迫的應答
真菌在生長發育和致病過程中會受到多種外界環境脅迫的影響,進一步研究CfVPS26基因是否響應細胞壁脅迫應答,將野生型菌株CFLH16、突變體ΔCfvps26和回補菌株ΔCfvps26/CfVPS26分別接種于含有細胞壁抑制劑CFW和SDS的 PDA培養基上,比較生長情況。結果表明,突變體ΔCfvps26對CFW和SDS敏感性增強,其生長抑制率高于野生型和回補菌株,且差異顯著(圖4A、B),說明CfVPS26基因參與果生刺盤孢對細胞壁脅迫應答。
為確定CfVPS26基因是否參與調控內質網壓力脅迫應答過程,將野生型菌株CFLH16、突變體菌株ΔCfvps26和回補菌株ΔCfvps26/CfVPS26分別接種于含有5 mmol·L-1二硫蘇糖醇DTT的PDA培養基上,比較生長抑制率。結果表明,突變體菌株ΔCfvps26對DTT耐受性顯著高于野生型菌株和回補菌株(圖4C、D),說明CfVPS26基因負調控果生刺盤孢應答內質網壓力脅迫過程。
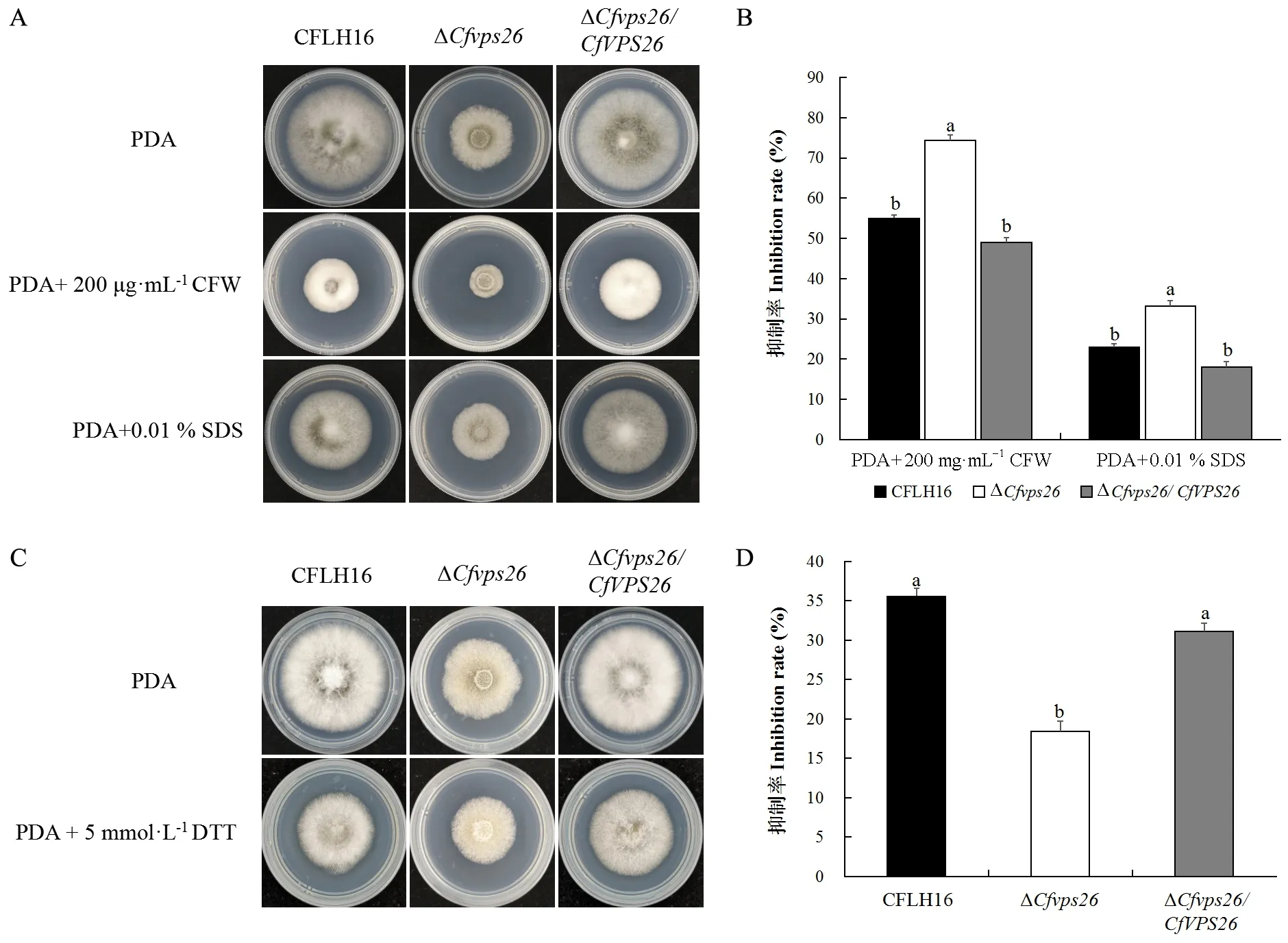
圖4 突變體ΔCfvps26參與外界環境脅迫應答Fig. 4 Growth assays of the ΔCfvps26 mutant on PDA with environmental stressorsA: 突變體ΔCfvps26在細胞壁脅迫下的生長情況; B: 抑制率統計; C:突變體ΔCfvps26在DTT脅迫下的生長情況; D: 抑制率統計。A: Growth of ΔCfvps26 under cell wall stress; B: Inhibition rates; C: Growth of ΔCfvps26 under DTT stress; D: Inhibition rates.
2.5 CfVPS26基因參與調控果生刺盤孢的致病力
對突變體ΔCfvps26、野生型和回補菌株進行了致病力測定,結果顯示,在離體接種3天后,與野生型和回補菌株相比突變體ΔCfvps26沒有產生病斑,喪失了對油茶葉片的致病力(圖5A、B)。以上結果說明CfVPS26基因參與調控果生刺盤孢的致病力。
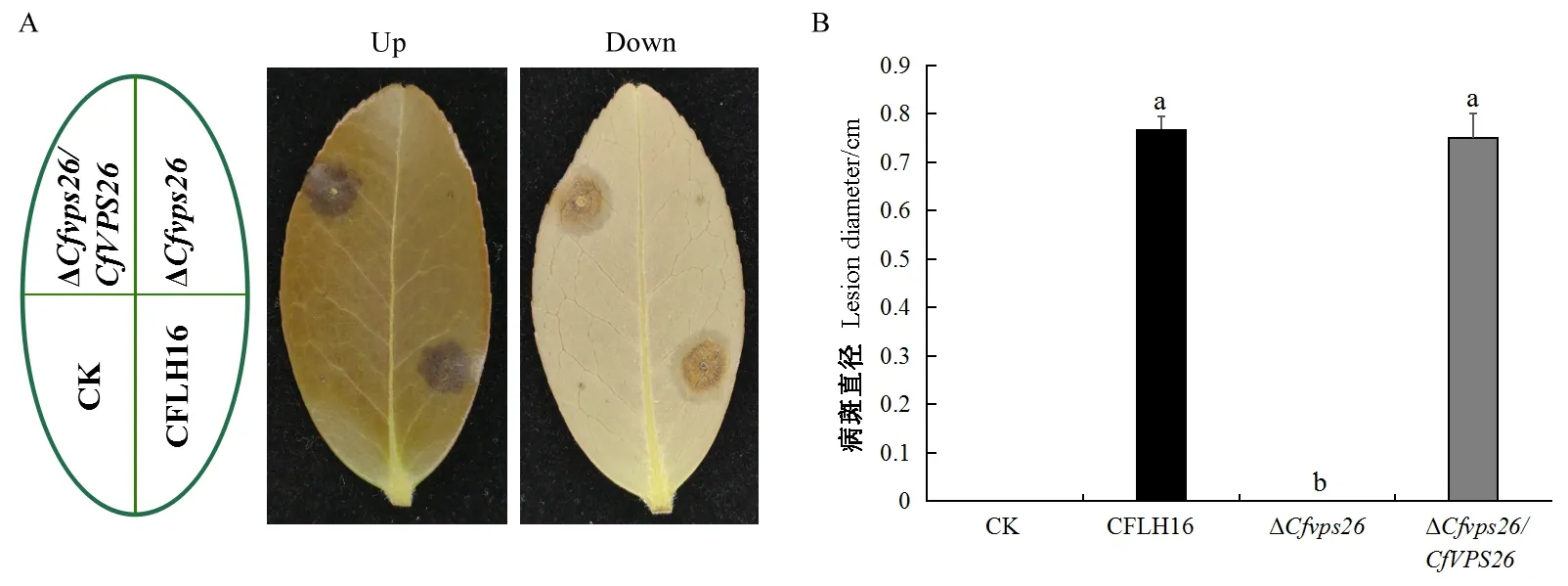
圖5 突變體ΔCfvps26對油茶葉片的致病力測定Fig. 5 Pathogenicity of ΔCfvps26 to oil-tea leavesA: 野生型、突變體ΔCfvps26和回補菌株接種在有傷葉片上; B: 病斑統計。A: CFLH16, ΔCfvps26 and ΔCfvps26/CfVPS26 were inoculated onto wounded leaves; B: Lesion diameter.
2.6 CfVPS26基因參與調控果生刺盤孢糖原代謝
附著胞內膨壓的形成來源于甘油的積累。分生孢子的萌發后,糖原物質會逐漸從分生孢子向附著胞轉移,隨后發生降解進而轉變為甘油(Wangetal., 2005)。本試驗利用糖原染色劑I2/KI對分生孢子及附著胞進行染色,結果顯示,野生型菌株分生孢子和附著胞糖原已經幾乎代謝完全,而突變體ΔCfvps26中仍然有大量的糖原存在(圖6),說明突變體中糖原代謝受阻。結果表明,CfVPS26基因參與調控果生刺盤孢的糖原代謝過程。

圖6 糖原染色Fig. 6 Staining for glycogen
3 討論
3.1 CfVPS26基因與營養生長的關系
菌絲是絲狀真菌營養生長的主要模式,侵染菌絲可以在寄主組織內擴展定殖。VPS26基因參與調控植物病原真菌菌絲生長,張瑩(2011)研究發現禾谷鐮刀菌(Fusariumgraminearum)中VPS26基因敲除突變體生長速率減慢、氣生菌絲明顯減少。本研究發現,果生刺盤孢的CfVPS26基因敲除后,突變體ΔCfvps26菌絲生長速率減慢,氣生菌絲量也顯著少于野生型和回補菌株。但黃清平(2013)研究發現,稻瘟病菌(Magnaportheoryzae)中的VPS26基因敲除后,其菌絲形態和生長速率則無明顯變化。因此VPS26基因在調控不同病原菌的菌絲生長時作用并不完全相同。
3.2 CfVPS26基因與內質網壓力的關系
內質網是蛋白質合成和分泌的重要場所,其穩態平衡在細胞生長發育中具有重要作用。當內質網負擔過重或壓力過大時,會導致蛋白質錯誤折疊,此時,細胞便會通過非折疊蛋白反應(unfolded protein response, UPR)來恢復內質網的正常功能。湯蔚(2015)研究發現,稻瘟菌的bZIP 轉錄因子Hac1和跨膜蛋白激酶IRE1均參與調控病菌內質網應激過程。筆者實驗室前期研究也表明,果生刺盤孢CfHAC1基因敲除突變體對5 mmol·L-1二硫蘇糖醇耐受性下降,說明轉錄因子CfHac1正調控病菌對內質網壓力脅迫應答過程(李司政等, 2020)。而本研究中,筆者發現果生刺盤孢的CfVPS26基因敲除突變體ΔCfvps26對5 mmol·L-1二硫蘇糖醇耐受性增強,說明該基因在應答二硫蘇糖醇造成的內質網壓力過程中發揮負調控作用,但其調控分子機制有待進一步深入研究。
3.3 CfVPS26基因與分生孢子產生的關系
植物病原真菌的分生孢子作為初侵染與再侵染的接種體,借氣流傳播從寄主表皮直接侵入致病。VPS26基因參與了植物病原真菌分生孢子的形成,對稻瘟病菌中基因VPS26進行敲除后,突變體菌株分生孢子產量顯著下降(黃清平, 2013)。而禾谷鐮刀菌中VPS26基因敲除突變體則幾乎不產孢或產生畸形孢子(張瑩, 2011)。在本研究中,筆者發現果生刺盤孢的突變體菌株ΔCfvps26的分生孢子產量也顯著下降。另外,筆者實驗室前期研究發現,果生刺盤孢中CfPMK1、CfHAC1、CfMKK1等基因均參與調控分生孢子的形成過程(李河, 2018; 姚權等, 2019; 肖宇等, 2021),說明在果生刺盤孢中存在多種基因參與調控分生孢子的形成這一過程。
3.4 CfVPS26基因與附著胞發育的關系
附著胞是侵染寄主的關鍵結構(Bechingeretal.,1999)。Kitade等(2015)研究發現玉米(Zeamays)小斑病菌(Bipolarismaydis)的STE7基因參與調控附著胞的形成。在膠胞刺盤胞(C.gloeosporioides)中的CgRGS7基因也影響了附著胞的形成過程(吳曼莉等, 2017)。前期對果生刺盤孢的研究發現,CfPMK1、CfSNF1和CfMKK1基因缺失突變體都不能形成附著胞(李河, 2018; Zhangetal., 2019; 肖宇等, 2021)。在本研究發現,突變體ΔCfvps26附著胞形成率顯著降低,這可能是導致其致病力喪失的原因之一。然而黃清平(2013)的研究發現稻瘟病菌中突變體ΔMoVps26對分生孢子的萌發和附著胞形成并無決定性作用,說明VPS26基因在不同的病原菌種類中功能存在差異。
3.5 附著胞發育與糖原代謝的關系
附著胞正常的膨壓對病原菌侵入寄主十分重要(Jongetal., 1997)。膨壓的產生需要大量甘油的累積,它來源于分生孢子內的糖原等能源物質,通過芽管運輸至附著胞降解形成甘油(Wangetal., 2005; Talbot, 2003)。在希金斯刺盤孢中,ChODC基因參與調控附著胞中糖原的轉移過程(嚴亞琴, 2020)。稻瘟病菌中基因MoEND3和MoCRN的缺失會導致糖原轉移和降解非常緩慢(李瀟, 2017)。在本研究中,果生刺盤孢野生型菌株CFLH16的糖原隨著附著胞發育成熟而逐漸代謝,而突變體ΔCfvps26分生孢子中仍有糖原滯留,附著胞內糖原代謝顯著減慢,說明糖原在突變體ΔCfvps26中的代謝受到了限制。由此推測CfPVS26基因參與了糖原的代謝過程,其敲除突變體不能形成正常的附著胞膨壓,進而喪失了對油茶葉片的致病力。
4 結論
本研究在油茶果生刺盤孢中鑒定到1個Retromer逆向囊泡轉運復合體亞基蛋白CfVps26,研究發現該蛋白參與調控病菌生長發育、產孢、附著胞形成、糖原代謝、外界脅迫應答和致病過程,研究結果有助于揭示果生刺盤孢致病的分子機制,并為開發殺菌劑提供潛在藥靶。
