基于聚類及簡化基因組聯合分析的浙江洞頭栽培羊棲菜(Sargassum fusiforme)品系篩選研究*
2021-10-11 09:51:42林立東吳明江
海洋與湖沼 2021年5期
林立東 吳明江
(1. 溫州市洞頭區海洋與漁業發展研究中心博士后工作站 溫州 325700; 2. 溫州大學生命與環境科學學院 溫州 325035)
羊棲菜[Sargassum fusiforme(Harvey)]為褐藻門(Phaeophyta) 、 圓 子 綱(Cyclospreae) 、 墨 角 藻 目(Fucales)、馬尾藻科(Sargassaceae)植物(曾呈奎,2000)。美國學者 Setchell(1931)將其歸為馬尾藻屬(Sargassum)反曲葉亞屬(Bactrophycus)中, 而日本藻類學家 Okamura(1932)則將其單列為羊棲菜屬(Hizikia) (吉田忠生, 2001)。目前, Algaebase 數據庫參考了相關文獻信息(Choet al, 2012; Dixonet al, 2014;Titlyanovet al, 2016; Huanget al, 2017), 僅采用Sargassum fusiforme拉丁名。
羊棲菜是太平洋西北沿岸特有的多年生大型褐藻, 主要分布于我國(北起遼東半島南至廣東雷州半島)、日本(北海道南部經本州至九州)和朝鮮半島(東岸、南岸及西南岸)近岸海域(曾呈奎, 2000)。目前, 羊棲菜野生種群主要分布于我國遼寧省大連市長海縣獐子島, 山東省煙臺市長島列島、威海榮成市鏌铘島和蘇山島, 浙江省舟山市嵊泗列島、枸杞島、嵊山島(種群數量最多)、溫州市洞頭列島(中國最大人工栽培基地)和南麂列島, 福建省漳浦縣古雷半島、福州市馬祖島和黃岐島、漳州市東山島, 廣東省汕頭市南澳島、湛江市硇洲島等地(李生堯等, 2010; 于深輝, 2012;徐佳麗等, 2014; 南春容等, 2015)。日本羊棲菜野生種群主要分布于北海道南部近海海域, 九州島長崎野母崎、長崎壹岐島和對馬巖原町島近岸海域。韓國羊棲菜野生種群主要分布于全羅南道省南部和濟州島(韓國最大人工栽培基地)近岸海域。
羊棲菜為食藥兩用海藻, 被推崇為益壽食品, 我國的《神農本草經》和《本草綱目》中記載了羊棲菜的療效(林立東等, 2019)。日本民眾食用羊棲菜的歷史悠久, 將羊棲菜美譽為“長壽菜”(何丹等, 2016), 并將每年的9 月15 日定為羊棲菜節, 16 歲成人禮要食羊棲菜。近年來, 日本羊棲菜市場的進口需求總量約4 500 t/年。浙江洞頭于1987 年首例實現野生羊棲菜干品出口日本貿易, 于1989 年開始以野生羊棲菜為苗種的試驗栽培, 成為我國最大的羊棲菜栽培、收購、加工和出口基地(楊志林, 1993)。1989—2000 年間栽培苗種主要依賴采集各地野生苗及假根再生苗,尤以浙江、福建沿海地區野生苗長勢最好、產量最高、品質最佳(何培民等, 2018)。2000 年以后羊棲菜有性生殖培苗技術逐漸成熟, 成為生產用苗種的主要來源。早期來自不同地域野生苗的基因在世代有性生殖過程中彼此交流, 形成了浙江洞頭栽培羊棲菜品系的多樣性。本文運用“特征”大氣囊表型和簡化基因組聯合分析方法, 系統歸納了浙江洞頭栽培羊棲菜群體品系特征, 以期為開展栽培羊棲菜品系多樣性與結構, 品系間親緣關系、優良品系篩選、品系生殖與繁衍特征、遺傳穩定性和品質差異等基礎研究, 以及遺傳育種、農業新品種選育、品系純化與擴繁等應用研究, 提供方法和理論支撐。
1 材料與方法
1.1 栽培羊棲菜品系“特征”大氣囊的選取、測量與樣品差異聚類分析
基于作者前期授權的國家發明專利“一種羊棲菜的品系鑒別和分類方法”(ZL201511004442.4) (林立東等, 2018), 以孢子體器官(假根、莖、氣囊、生殖托等)方差貢獻率最大的“特征”大氣囊表型為標準, 2018 年4 月2 日, 挑選浙江洞頭栽培羊棲菜多態性植株21 個,置4 °C 保存; 再分別選取各樣品二級側生枝“特征”大氣囊(≥10 支) (圖1), 置于事先注入適量新鮮過濾海水的培養皿中, 拍照記錄形態(圖2); 選取羊棲菜各樣品10 支“特征”大氣囊, 參照測量單個氣囊總長、囊尖長、囊體長、囊體寬、囊柄長及鮮重, 記錄數據,計算均值與標準差, 均值數據代入SPSS16.0 統計軟件, 距離閾值T=0.9, 聚類判定樣品表型差異。
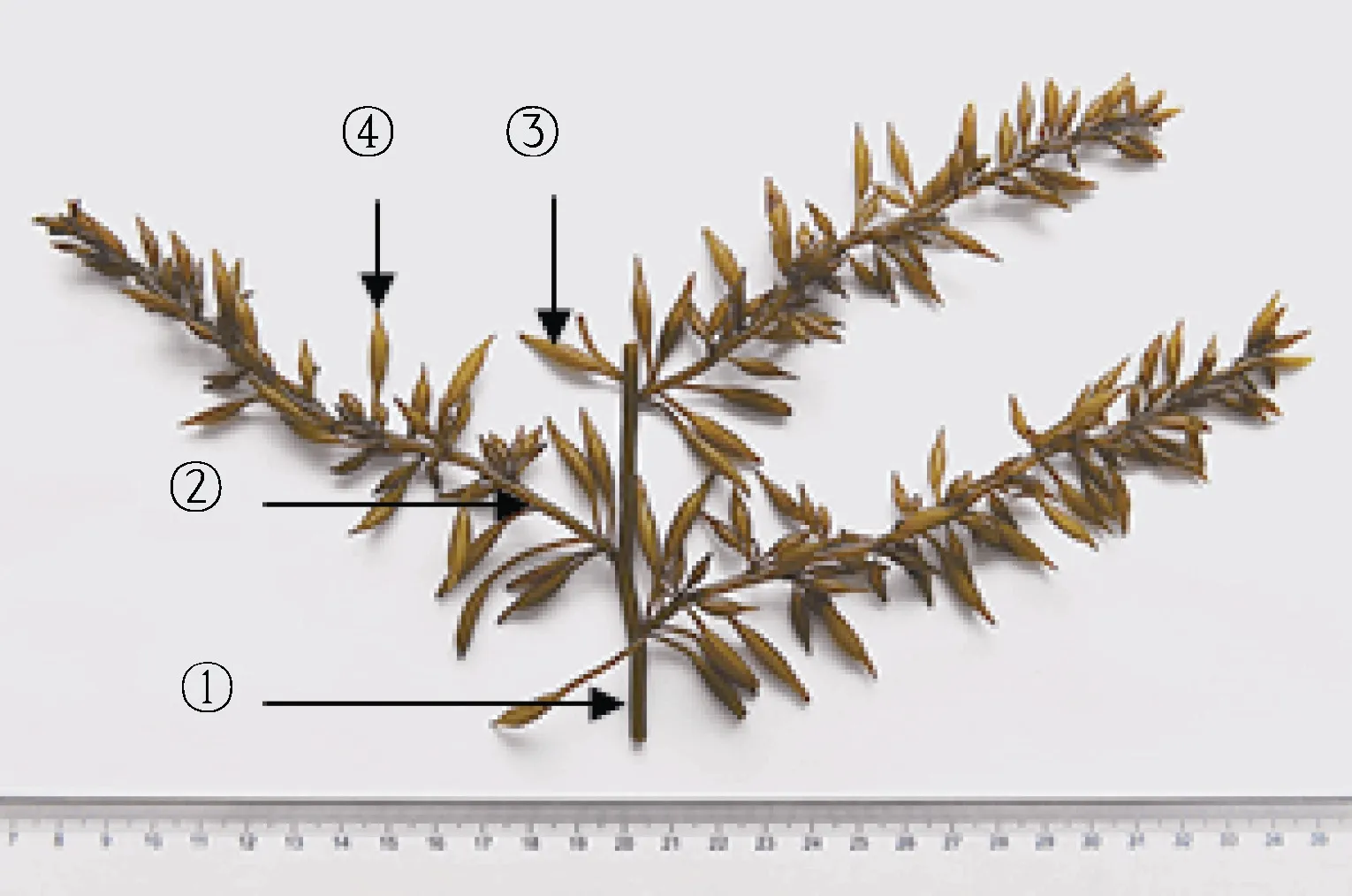
圖1 栽培羊棲菜側生莖和“特征”大氣囊Fig.1 Lateral stem and characteristic big air-bladder of cultivated S. fusiforme


圖2 栽培羊棲菜“特征”大氣囊形態Fig.2 Characteristic big air-bladder morphologys of cultivated Sargassum fusiforme
1.2 羊棲菜品系簡化基因組的測序與系統進化樹分析
1.2.1 羊棲菜樣品的采集 于2017 年11 月和2018 年4 月份兩個批次采集樣品, 共計34 個樣品(表1)。取各樣品側生枝20 g, 置封口袋封存, 置4 °C 保存待測。
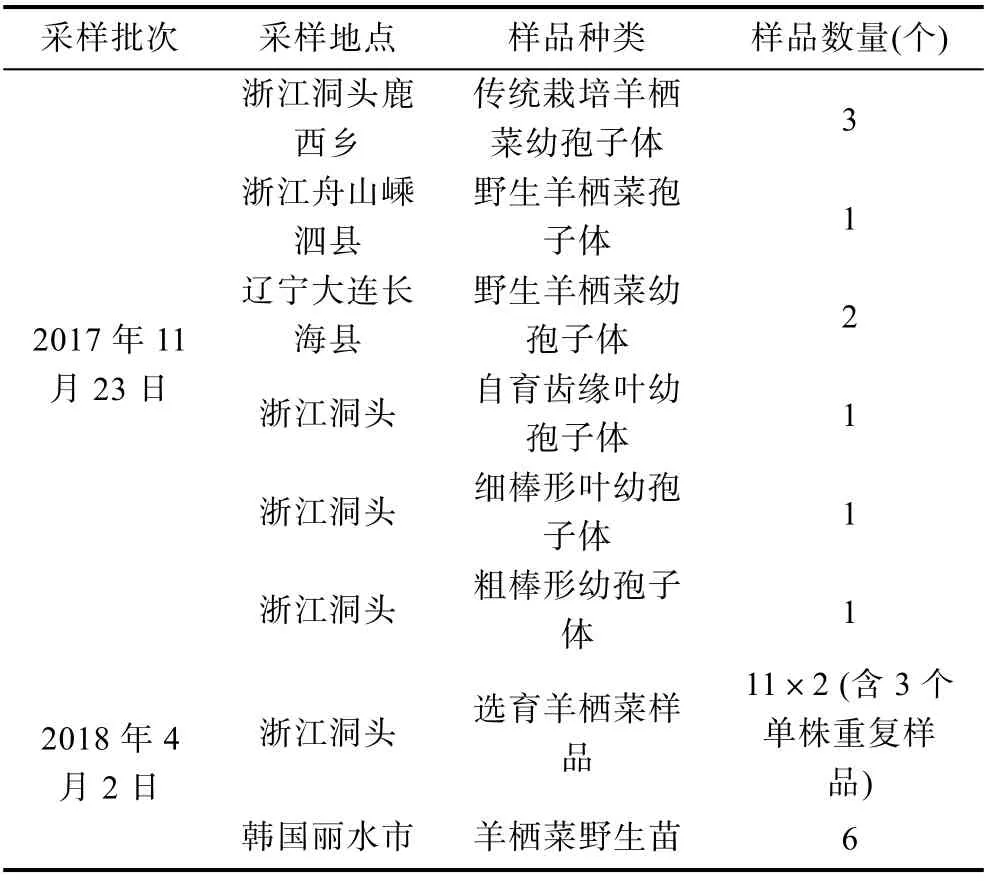
表1 栽培羊棲菜樣品的采集信息Tab.1 Information of sampling for cultivated S. fusiforme
1.2.2 羊棲菜樣品的DNA 提取 精確稱取葉片或氣囊1.0 g, 置于液氮預冷的研缽中研磨成粉末; 轉入2 mL 離心管中, 加入750 μL CTAB 溶液(用前預熱至65 °C, 并加入3%—5% β 巰基乙醇), 渦旋混勻;65 °C 恒溫水浴30 min 后, 加入700 μL 的氯仿:異戊醇(24 : 1), 搖至乳白色; 4 °C、10 000 r/min 離心10 min, 取上清液650 μL 置2 mL 離心管中, 加入預冷異丙醇700 μL, 再加3 mol/L NaAc 100 μL, 加蓋,輕輕混勻, 至白色絮狀沉淀析出; 緩慢挑出白色絮狀沉淀至1.5 mL 離心管(預先加入1 mL 70%酒精), 輕搖洗去雜質; 4 °C、12 000 r/min 離心10 min, 倒掉酒精, 將沉淀DNA 置陰涼通風處, 使酒精全部自然揮發; 加入含有0.5% RNA 消化酶的T.E 溶液, 于4 °C 溶解DNA 12 h, 將溶解好的DNA 進行SDS 電泳檢測, 待用。
1.2.3 羊棲菜樣品的簡化基因組測序與DNA 文庫構建 利用參考基因組序列模擬各種限制性內切酶的酶切位點的數量與分布, 選擇合適的酶進行酶切建庫; 本實驗采用Reads1 端EcoRI (G^AATTC)和Read2 端NlaIII (Hin1II, CATG^)進行雙酶切。質檢合格的DNA 樣品, 采用ddRAD 建庫方式構建長度范圍在300—500 bp 的pair-end 文庫。具體實驗內容包括:(1) 取DNA 500 ng, 加入0.6 UEcoRI (NEB)、T4DNA連接酶(NEB)、ATP (NEB)和EcoRI 接頭(含區分樣品的Index 序列), 37 °C 反應3 h, 65 °C 退火1 h; 然后加限制性內切酶NlaIII (NEB)和NlaIII 接頭在37 °C 下反應3 h, 反應結束后在65 °C PCR 儀中放置30 min失活內切酶。(2) 使用瓊脂糖凝膠電泳對連接產物進行片段選擇, 選擇400—600 bp 回收酶切產物。(3) 使用Qubit3.0 (Life Technology)對回收產物進行DNA 定量, 等量混合34 個樣品。(4) 使用Illumina TruSeq 試劑盒對混合產物進行DNA 文庫構建。
1.2.4 羊棲菜樣品的簡化基因組系統進化樹分析
(1) 信息分析: 對測序得到的原始reads (雙端序列)進行數據評估, 得到各個樣品的原始reads, 將reads 進行比對聚類, 獲得ddRAD 片段后進行SNP 檢測。
(2) 系統進化分析: 基于SNP, 通過FastTree 軟件,做樣品間的系統進化分析, 計算得到群體進化樹。
1.3 栽培羊棲菜品系學名命名
根據林奈(Carl von Linné)的“雙名法”, 采用羊棲菜拉丁文“Sargassum fusiforme”命名規則, 對浙江洞頭栽培羊棲菜5 個品系進行命名。“Sargassum”為屬名拉丁文, “fusiforme”為種加詞, 屬形容詞, 中文解釋為梭形的、紡錠狀的或兩端漸細的。據此, 本文采用“屬名+種加詞(adj)+命名人+新品系標識”規則予以學名命名, 并對命名品系加以“模式指定”。
2 結果
2.1 浙江洞頭栽培羊棲菜樣品的“特征”大氣囊表型指標
浙江洞頭栽培羊棲菜樣品二級側枝“特征”大氣囊表型包括氣囊總長、囊尖長、囊體長、囊體寬、囊柄長和氣囊鮮重6 個指標, 測量結果顯示不同樣品表型參數差異較大(表 2)。各樣品氣囊總長介于(22.57±2.85)—(56.65±3.09) mm, 有尖或無尖, 囊尖長度介于(2.57±1.34)—(13.47±2.11) mm, 囊體長介于(8.82±0.36)—(27.86±1.23) mm, 囊 體 寬 介 于(2.50±0.28)—(5.60±0.48) mm, 囊 柄 長 介 于(8.00±2.49)—(28.24±2.72) mm, 鮮 重 介 于(43.70±10.00)—(200.80±20.00) mg。
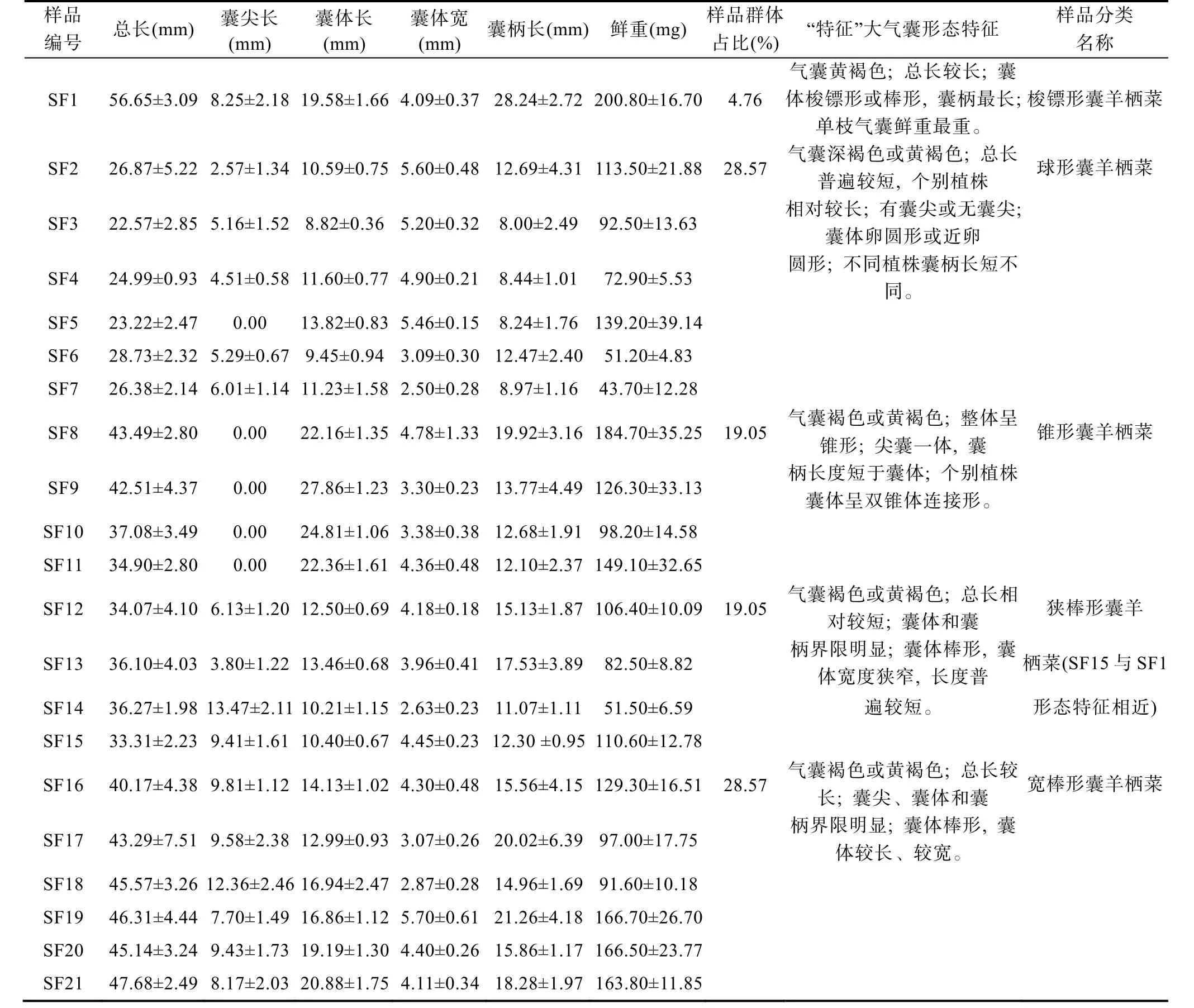
表2 浙江洞頭栽培羊棲菜樣品“特征”大氣囊表型指標和生物學基本特征Tab.2 Phenotypic parameters and biological characteristics of characteristic big air-bladders of S. fusiforme sampled from Dongtou,Zhejiang
依據氣囊顏色、總長、有無囊尖、囊體性狀和囊柄長短5 個方面的差異, 將洞頭栽培羊棲菜樣品“特征”大氣囊初步劃分為梭鏢形、卵圓形、錐形、狹棒囊形和寬棒囊形5 種“特征”大氣囊表型。梭鏢形表型樣品占比總樣品數最少, 為4.76%, 卵圓形和寬棒囊形表型樣品占比最高, 分別為28.57%, 錐形囊和狹棒囊形表型樣品占比居中, 分別為19.05%。
2.2 浙江洞頭羊棲菜樣品的“特征”大氣囊表型指標參數聚類分析
羊棲菜樣品“特征”大氣囊表型參數聚類分析結果如圖3 所示。根據系統聚類最短距離法判定分析如下:
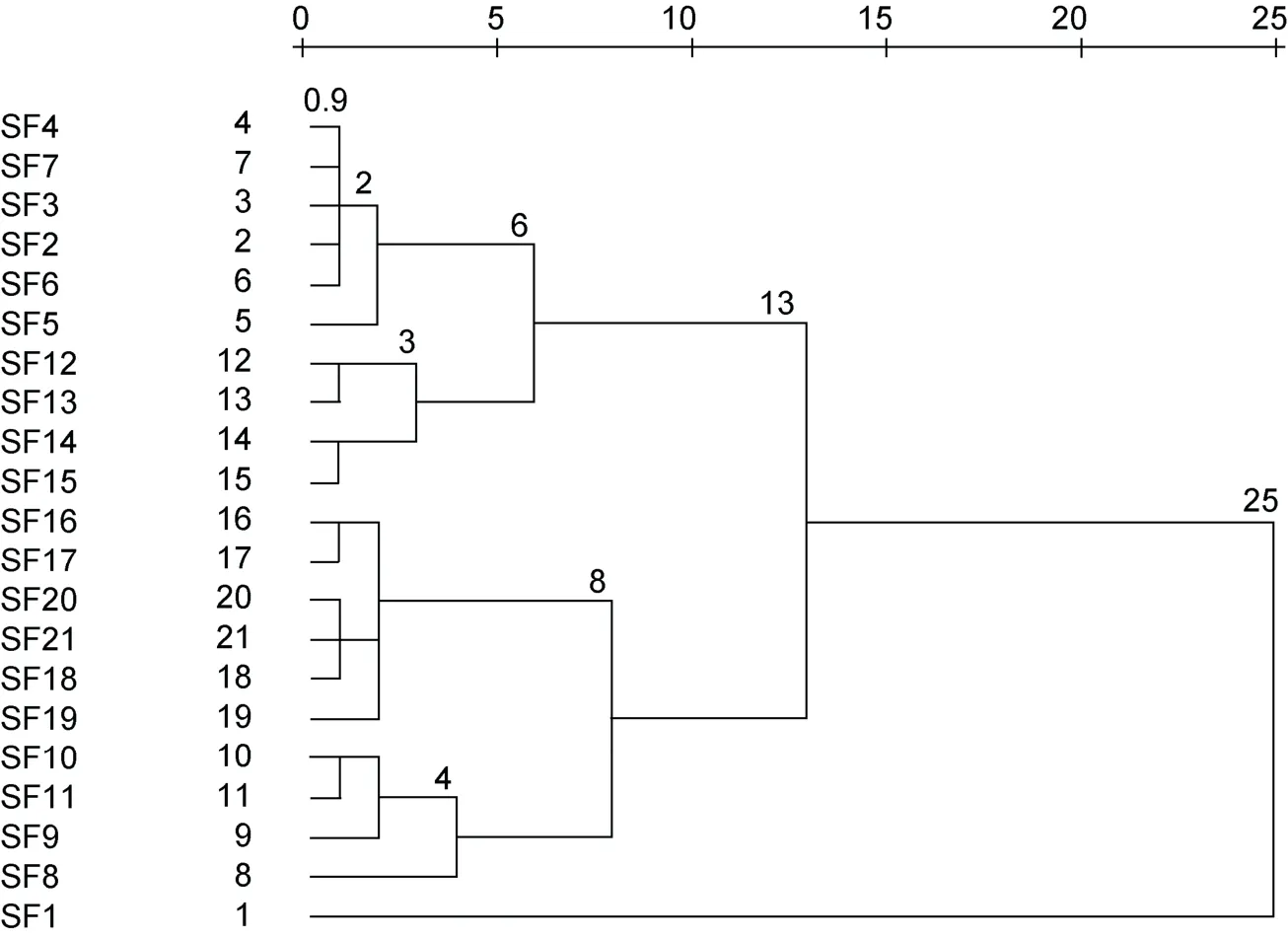
圖3 平均(不同群間)重新標度距離聚類樹狀圖Fig.3 Dendrogram using average (between groups) rescaled distance cluster combining
當T=0.9 時, 21 個樣品各成一個表型品系; 當0.9≤T<2.0 時, 21 個樣品可分成11 個表型品系, 即: 品系1 (SF2、SF3、SF4、SF6 和SF7)、品系2 (SF5)、品系3 (SF12 和SF13)、品系4 (SF14 和SF15)、品系5 (SF16 和SF17)、品系6 (SF18、SF20 和SF21)、品系7 (SF19)、品系8 (SF10 和SF11)、品系9 (SF9)、品系10 (SF8)及品系11 (SF1)。
當2.0≤T<3.0 時, 21 個樣品可分成7 個表型品系,即: 品系1 (SF2、SF3、SF4、SF5、SF6 和SF7)、品系2 (SF12 和SF13)、品系3 (SF14 和SF15)、品系4(SF16、SF17、SF18、SF19、SF20 和SF21)、品系5 (SF9、SF10 和SF11)、品系6 (SF8)和品系7 (SF1)。
當3.0≤T<4.0 時, 21 個樣品可分成6 個表型品系,即: 品系1 (SF2、SF3、SF4、SF5、SF6 和SF7)、品系2 (SF12、SF13、SF14 和SF15)、品系3 (SF16、SF17、SF18、SF19、SF20 和SF21)、品系4 (SF9、SF10 和SF11)、品系5 (SF8)和品系6 (SF1)。
當4.0≤T<6.0 時, 21 個樣品可分成5 個表型品系,即: 品系1 (SF2、SF3、SF4、SF5、SF6 和SF7)、品系2 (SF12、SF13、SF14 和SF15)、品系3 (SF16、SF17、SF18、SF19、SF20 和SF21)、品系4 (SF8、SF9、SF10和SF11)和品系5 (SF1)。
當6.0≤T<8.0 時, 21 個樣品可分成4 個表型品系,即: 品系1 (SF2、SF3、SF4、SF5、SF6、SF7、SF12、SF13、SF14 和SF15)、品系2 (SF16、SF17、SF18、SF19、SF20 和SF21)、品系3 (SF8、SF9、SF10 和SF11)和品系4 (SF1)。
當8.0≤T<13.0 時, 21 個樣品可分成3 個表型品系,即: 品系1 (SF2、SF3、SF4、SF5、SF6、SF7、SF12、SF13、SF14 和SF15)、品系2 (SF16、SF17、SF18、SF19、SF20、SF21、SF8、SF9、SF10 和SF11)和品系3 (SF1)。
當13.0≤T<25 時, 21 個樣品可分為2 個表型品系,即: 品系1 (SF2、SF3、SF4、SF5、SF6、SF7、SF12、SF13、SF14、SF15、SF16、SF17、SF18、SF19、SF20、SF21、SF8、SF9、SF10 和SF11)、品系2 (SF1)。
當T≥25 時, 21 個樣品僅為1 個表型品系。
以閾值T=0.9 為基數,T值距離2.0、3.0、4.0、6.0、8.0、13.0 和25.0 之間的相似度分別為45%、30%、22.5%、15%、11.25%、6.9%和3.6%, 差異貢獻率(KMO,Kaiser-Meyer-Olkin)分別為55%、70%、77.5%、85%、88.75%、93.1%、96.4%。參照Kaiser 的KMO 檢驗統計變量是否適合因子分析的5 級判定標準(本組設定KMO>90%, 非常適合分類; 85%≤KMO≤90%, 適合分類; 80%≤KMO<85%, 一般適合分類; 75%≤KMO<80%, 不太適合分類, 比較勉強; KMO<75%,不適合分類), 作為栽培羊棲菜“特征”大氣囊表型品系聚類分類判定標準, 即: 4.0≤T<6.0 時, 差異貢獻率為85%, 符合“適合分類”標準。
依據以上聚類分析結果和Kaiser 的判定標準, 本文將浙江洞頭栽培羊棲菜“特征”大氣囊表型品系分為5 類, 即: 表型品系1-球囊形羊棲菜(SF2、SF3、SF4、SF5、SF6 和SF7)、表型品系2-狹棒囊羊棲菜(SF12、SF13、SF14 和SF15)、表型品系3-寬棒囊羊棲菜(SF16、SF17、SF18、SF19、SF20 和SF21)、表型品系4-錐形囊羊棲菜(SF8、SF9、SF10 和SF11)和表型品系5-梭鏢形羊棲菜(SF1)。
2.3 羊棲菜樣品的簡化基因組測序和系統進化樹分析
2.3.1 ddRAD 標記多態性統計 羊棲菜樣品間的Tag 標簽進行比對聚類(blat 軟件), 聚類時將identity在95%以上的Tag 標簽聚類成Marker(標記), 且將小于2 個Tag 標簽的標記定義為非多態性標記, 其余為多態性標記。羊棲菜34 個樣品ddRAD 標記多態性結果(表3)表明, 非多態性標記(nonPoly)數量為225 328,總測序深度為11 712 620, 平均深度為52; 多態性標記(Poly)數量為167 777, 總測序深度為30 773 677,平均深度為183。
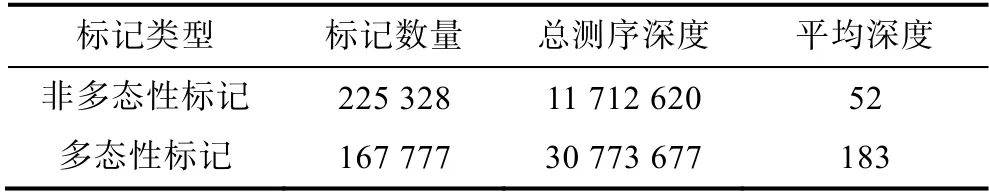
表3 多態性標記統計Tab.3 Statistics of polymorphic markers
2.3.2 羊棲菜Marker 數量統計分析 羊棲菜34個樣品Marker 數量均不相同(圖4), 且差異顯著, 其中較多Marker 數量樣品包括R16、R17、R18、R19和R24 (隨機編組, 詳見圖4), 數量分別為140 962、151 815、145 605、139 360 和115 548; 較少Marker數量樣品包括R01、R02、R03、R04 和R06, 數量分別為17 852、17 945、23 654、20 724 和28 170, 其他樣品Marker 數量介于二者之間。
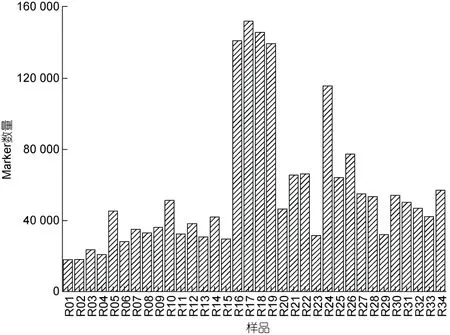
圖4 羊棲菜樣品間的Marker 深度分布Fig.4 Marker depth distribution between S. fusiforme samples
2.3.3 羊棲菜樣品簡化基因組群體系統進化分析浙江洞頭栽培羊棲菜、野生羊棲菜(韓國麗水、遼寧大連和浙江舟山)及栽培羊棲菜幼孢子體簡化基因組系統進化樹特征表明(圖5), 浙江洞頭栽培羊棲菜樣品與不同地域野生羊棲菜之間具有不同的親緣關系;根據簡化基因組進化樹最遠距離特征, 全部樣品具備5 個典型基因型品系聚類特征, 即: 基因型品系1(韓國野生羊棲菜1 和5, 鹿西培育幼孢子體1、2 和3, SF1、SF4、SF6 和SF7)、基因型品系2 (舟山野生幼孢子體, 自育齒緣葉、細棒葉和粗棒葉幼孢子體, SF2、SF17、SF20 和SF21)、基因型品系3 (SF8、SF9、SF10、SF11 和SF18)、基因型品系4 (韓國野生羊棲菜2、3 和4, SF15)和基因型品系5 (大連野生羊棲菜1 和2, SF14)。
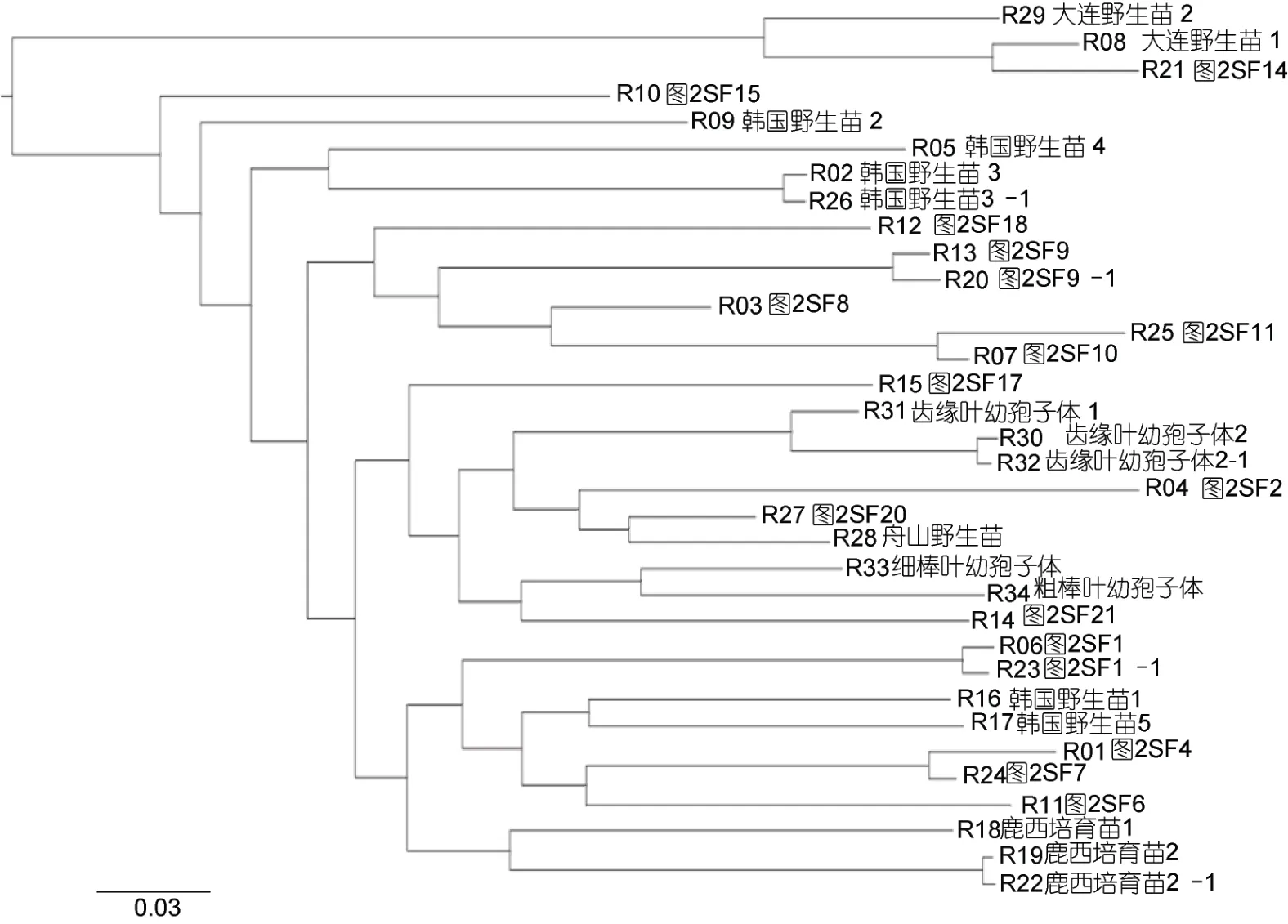
圖5 羊棲菜樣品的簡化基因組系統進化樹Fig.5 A simplified genome phylogenetic tree of S. fusiforme samples
2.3.4 羊棲菜樣品簡化基因組重復測序驗證 通過對韓國野生羊棲菜樣品3、栽培羊棲菜樣品1 與樣品9、洞頭栽培羊棲菜齒緣葉幼孢子體和鹿西培育羊棲菜幼孢子體等5 個同株樣品簡化基因組重復測序,結果表明不同樣品簡化基因組遺傳距離差異顯著,而同株不同部位的簡化基因組無差異(圖5)。
2.4 浙江洞頭栽培羊棲菜的基因型品系與表型品系比較分析
栽培羊棲菜基因型品系1 中 R01 (SF4)、R11(SF6)和R24 (SF7)樣品的基因型與表型品系1 相符,R06 (SF1)樣品的基因型與表型品系1 不符, 四者基因遺傳距離不同, 但親緣關系較近, 且四者均與韓國野生羊棲菜和鹿西培育羊棲菜具有較近的親緣關系。
栽培羊棲菜基因型品系2 中R15 (SF17)、R27(SF20)和R14 (SF21)樣品的基因型與表型品系3 分類相符, R04 (SF2)樣品的基因型與表型品系3 不符, 四者基因遺傳距離不同, 但親緣關系較近, 且四者均與舟山野生羊棲菜及自育齒緣葉、細棒葉、粗棒葉羊棲菜幼孢子體親緣關系較近。
栽培羊棲菜基因型品系3 中R03 (SF8)、R13 (SF9)、R07 (SF10)和R25 (SF11)樣品的基因型與表型品系4 相符, R12 (SF18)樣品的基因型與表型品系4 不符, 5 者基因遺傳距離不同, 但親緣關系較近, 且5 者均與韓國、大連和舟山野生羊棲菜之間具有較遠的親緣關系。
栽培羊棲菜基因型品系4 中R10 (SF15)樣品的基因型與表型品系5 相符, 它與韓國野生羊棲菜2、3和4 樣品的親緣關系較近, 它們與韓國野生羊棲菜本1 和5 樣品相比, 呈同源產地、不同樣品間基因遺傳距離不同的特征。
栽培羊棲菜基因型品系5 中R21 (SF14)樣品的基因型與表型品系2 相符, 它的基因遺傳距離最大, 基因型與大連野生羊棲菜基因型接近。
樣品基因型與樣品表型品系間重合度結果表明,基因型品系1 與樣品表型品系1、基因型品系2 與樣品表型品系3、基因型品系3 與樣品表型品系4、基因型品系4 與樣品表型品系5 及基因型品系5 與樣品表型品系2 之間的重合率分別為75%、100%、75%、80%、100%, 樣品平均總重合率為86%。
2.5 浙江洞頭栽培羊棲菜的5 個典型品系學名命名
綜合上述研究結果, 本文將浙江洞頭栽培羊棲菜劃分為球囊羊棲菜、錐囊羊棲菜、寬棒囊羊棲菜、窄棒囊羊棲菜和梭鏢囊羊棲菜5 個品系(表4); 根據生物學分類及等級關系、林奈的動植物“雙名法”及品系“模式指定”要求(Singh, 2004), 對5 個新劃分的栽培羊棲菜品系并加以學名命名和模式指定。

表4 栽培羊棲菜品系分類、學名命名和模式指定Tab.4 Classification, scientific nomenclature and pattern specification of cultivated S. fusiforme strains
3 討論
3.1 浙江洞頭栽培羊棲菜群體的親本來源與“特征”大氣囊表型多樣性
浙江洞頭于1989 年起以野生羊棲菜幼孢子體為苗種, 率先在國內探索性栽培羊棲菜(李生堯, 2001)。1989—1994 年間, 在出口市場需求驅動下, 羊棲菜栽培面積逐年擴大, 羊棲菜苗種的需求量也越來越大,羊棲菜苗種的采集地也從溫州本土拓展到中國、日本和韓國等沿海地區, 形成了洞頭栽培羊棲菜群體親本的多樣性來源, 使得現有栽培羊棲菜群體“特征”大氣囊表型呈現出了多樣性特征。
關于馬尾藻屬海藻的形態描述, 尚無統一標準。單體鋒等(2009)采用羊棲菜藻體鮮重、藻體長度、側枝密度、側枝長度和側枝鮮重等5 個藻體性狀指標,描述不同品系差異(單體鋒等, 2009)。李生堯等(2010)報道了“鹿豐1 號”羊棲菜, 詳細描述了枝葉粗壯繁茂、氣囊產生早、顆粒大、產量高等性狀特征, 且與舟山東福和嵊山野生種源羊棲菜比對了“特征”大氣囊、主枝和側枝的直徑、長度和總量等基礎數據, 但未采用科學方法進行主成分分析, 無法進行“特征”大氣囊表型辨別和驗證, 致使“鹿豐1 號”羊棲菜品系信息至今停留在基礎數據層面。Yao 等(2019)運用RAPD和ISSR 標記技術對洞頭栽培羊棲菜品系進行了遺傳分析, 但未對所用的兩個鹿豐1 號植株樣品及另外兩個羊棲菜樣品的表型特征作出明確描述(Yaoet al,2019)。現有洞頭栽培羊棲菜群體多樣性研究報道表明, 洞頭栽培羊棲菜群體“特征”大氣囊表型多樣性客觀存在, 但僅有Li 等(2018)在關于不同形態氣囊褐藻羊棲菜生物活性物質及營養成分的比較研究中,列舉了兩個樣品的枝狀體, 其他研究報道均未明確標識羊棲菜表型主成分“特征”大氣囊的形態特征。
本文以“同種生物同一性狀的不同表現形式稱為相對性狀”為理論依據(林立東等, 2018), 在多年跟蹤監測栽培羊棲菜樣品季節性單株鮮重、干重、長度、側生枝數、莖寬、假根長度, 以及“特征”大氣囊的總長度、寬度、囊尖長度、囊柄長度和鮮重等11 項性狀指標基礎上, 通過羊棲菜成熟孢子體性狀指標相關性數學模型、主成分數學模型、主成分聚類數學模型等綜合分析, 確定羊棲菜性狀指標中“特征”大氣囊表型為性狀主成分, 貢獻率最高。上述判定符合栽培羊棲菜成熟藻體周身氣囊, 氣囊簇生, 每簇著生1—2支“特征”大氣囊的特征描述(林立東等, 2020)。本研究以栽培羊棲菜“特征”大氣囊表型和指標數據作為品系劃分依據, 首創了定性+定量聯合判定栽培羊棲菜品系表型方法, 使表型品系分類更加符合現代植物學、植物分類學和生物統計學的交叉學科特征, 補充了傳統植物分類學僅依據直根或須根、單主莖或多主莖、葉對生或互生、花萼三基數或五基數和果實單心皮或兩心皮等過于簡單的定性定量分類的不足。
洞頭栽培羊棲菜“特征”大氣囊表型差異表明, 來源于同一地點的野生羊棲菜, 經馴化栽培, “特征”大氣囊表型不盡相同。本研究通過栽培羊棲菜群體簡化基因組分析表明, 洞頭栽培羊棲菜球囊形、粗棒囊形和狹棒囊形品系分別與韓國麗水、浙江舟山和遼寧大連等地野生羊棲菜的親緣關系較近; 聯合分析結果表明, 羊棲菜基因型品系樣品與表型品系樣品之間的總重合率為86%, 說明當年采自上述三地的野生苗種在洞頭雖歷經三十年的雜交及有性生殖繁衍,仍保持遺傳的相對穩定性。然而, 控制氣囊表型的基因定位, 以及不同“特征”大氣囊表型羊棲菜間經有性生殖產生的子代基因型屬“父系遺傳”, 還是屬“母系遺傳”等問題尚未見文獻報道, 還有待深入研究。
3.2 浙江洞頭栽培羊棲菜群體的遺傳多樣性
浙江洞頭栽培羊棲菜品系遺傳多樣性問題一直深受國內研究者們的密切關注。單體鋒等(2009)運用AFLP 法研究了浙江洞頭羊棲菜1 個代表性栽培群體的遺傳多態性, 多態位點比率為93.8%; 呂慧等(2009)運用RAPD 技術研究了浙江洞頭羊棲菜栽培中常見的3 個品系的遺傳多樣性, 多態位點比率為84.4%;徐佳麗等(2014)研究了5 種不同品系羊棲菜(原始采集地分別是韓國、浙江洞頭、浙江南麂列島、浙江東極島及廣東汕頭)的遺傳多樣性, 多態性比率為58.9%; 于深輝(2012)運用ISSR 技術研究了我國北起遼寧南至廣東沿海的9 個野生羊棲菜種群遺傳多樣性, 多態性比率為 99.61%; 南春容等(2015)運用ISSR 技術對浙江洞頭羊棲菜野生與選育種群的研究表明, 浙江南麂島和浙江竹嶼島野生羊棲菜遺傳相似度最大, 二者與洞頭栽培羊棲菜三個品系間遺傳距離較遠。
上述研究結果表明, 地理隔離限制了群體間基因流動, 形成了自然種群獨特的遺傳特征, 因而野生羊棲菜群體的遺傳變異性和遺傳多樣性普遍高于栽培羊棲菜, 種群間的遺傳分化大于種群內。上述2009年的研究數據顯示, 無論采用哪種分子標記技術和哪種樣品采集方式, 浙江洞頭栽培羊棲菜品系較之野生型羊棲菜的遺傳多態性比例降低約15%, 2014 年降低了40%, 說明早期采自各地的野生羊棲菜在有性生殖繁育過程中發生了基因流動, 且隨著時間推移基因流動性會越來越大, 栽培群體的遺傳多態性越來越低。
上述研究報道中, 樣本的選擇要么都是栽培樣本, 要么都是野生樣本; 即便是存在選用兩類樣本對比分析的情況, 樣本采集量卻不夠全面。為了全視野地分析現階段浙江洞頭栽培羊棲菜群體遺傳多態性,深層次地挖掘栽培羊棲菜群體的親本來源及遺傳分化與變異規律, 本文全面選取浙江洞頭栽培羊棲菜、野生羊棲菜(韓國麗水、遼寧大連和浙江舟山)及栽培羊棲菜幼孢子體等樣品, 構建了大樣本簡化基因組系統進化樹, 確立了5 個基因型品系。韓國麗水、遼寧大連和浙江舟山野生羊棲菜遺傳多樣性差異較大,體現了地理隔離對群體間基因流動的限制, 從而形成了自然種群獨特的遺傳特征; 韓國野生羊棲菜樣品與本地選育的羊棲菜優良品系遺傳關系較近, 說明早期來自韓國的羊棲菜苗種在人工有性生殖培育進程中得以基因延續, 該特征既體現了韓國羊棲菜遺傳的相對穩定性, 又體現了在韓國羊棲菜深受國際市場青睞背景下, 科研單位和育種企業對韓國羊棲菜的長期性定向選育和示范推廣。
3.3 浙江洞頭栽培羊棲菜樣品、系、種和新品種的關系與學名命名
羊棲菜基因與環境之間存在著復雜的相互作用,精細地調控著枝狀體的性狀, 即: 表型=基因型+環境因素。浙江洞頭栽培羊棲菜“特征”大氣囊表型多樣性說明, 不同地源野生羊棲菜對洞頭海洋環境均有適應性, 在長期繁衍過程中均表現出了多樣性的氣囊表型, 而不同“特征”大氣囊表型植株間的基因組親緣關系有近有遠, 存在與野生羊棲菜較近的栽培樣品。羊棲菜植株的生長發育呈非同步性, 簇生氣囊的生長發育呈漸變性, 使得相同培育批次、相同生長環境、相同生長時間的羊棲菜植株之間“特征”大氣囊表型不盡相同。因此, 洞頭栽培羊棲菜群體的表型和遺傳特征極具復雜性。有關基礎研究若無科學的表型和基因型判定依據, 所采集的植株僅能定義為生物統計學中的樣品或樣本。生物學分類單位及層次關系包括: 界(Kingdom)、門(Phylum 或Division)、綱(Class)、目(Order)、科(Family)、族(Tribe)、屬(Genus)、組(Section)、系(Series)、種(Species)、變種(Variety)、變型(Form)、栽培品種(Cultivar) (陸寶樹, 1990)。羊棲菜分類為馬尾藻屬, 為單支屬海藻。本文通過表型和基因型雙重判定法, 將洞頭羊棲菜栽培群體劃分為“系”或“品系”。被確認的“系”經多年選育、純化、擴繁和示范推廣, 獲得了穩定表型、基因型、產量和營養品質等特征后, 可申請認定為生物學分類中的“種”;若再具備穩定的純化與繁育基地, 多態性(多地區)栽培、規模化栽培及獲得顯著的經濟效益等條件, 經國家良種審定委員會審定, 可認定為新品種。
現有涉及栽培羊棲菜“品系”的相關文獻報道, 所使用的栽培羊棲菜樣品均來自于浙江洞頭, 未出現其他地區栽培樣品信息, 這一研究樣品信息, 符合生物學分類中“系”的生境限定要求。本文在氣囊表型和基因型雙重判定基礎上, 將浙江洞頭栽培羊棲菜群體系統劃分了5 個品系(表4), 在林奈的“雙名法”規則基礎上, 分別予以學名命名, 為栽培羊棲菜的生物學研究及育種實踐應用提供了更加準確的品系分類信息。
4 結論
羊棲菜性狀差異的主成分為簇生氣囊中的“特征”大氣囊。浙江洞頭栽培羊棲菜群體“特征”大氣囊表型多樣性源于多樣性地源種的混雜栽培; 不同地源野生羊棲菜對洞頭海洋環境均有適應性, 在長期繁衍過程中均表現出了多樣性的“特征”大氣囊表型;不同“特征”大氣囊表型植株簡化基因組之間的親緣關系有近有遠, 群體中存在與野生羊棲菜較近的栽培樣品。浙江洞頭栽培羊棲菜表型品系和基因型品系的總重合率為86%, 適合品系多樣性分類條件。浙江洞頭栽培羊棲菜群體可分球囊(Sargassum sphaer-bladderL. D. Lin et M. J. Wu, spp.nov.)、錐囊(Sargassum turgidum-bladderL. D. Lin et M. J. Wu,spp.nov.)、寬棒囊(Sargassum wide-cylindricalL. D.Lin et M. J. Wu, spp.nov.)、狹棒囊(Sargassum strictures-cylindricalL. D. Lin et M. J. Wu, spp.nov.)和梭鏢囊羊棲菜(Sargassum dart-shaped bladderL. D.Lin et M. J. Wu, spp.nov.) 5個品系。浙江洞頭栽培羊棲菜5個品系均具有穩定的表型、生活史、產量和生化組成等特征, 若長期純化、擴繁和示范推廣, 均可培育為新品種。
猜你喜歡
數學小靈通·3-4年級(2024年2期)2024-05-15 02:02:28
考試與評價·高二版(2021年5期)2021-09-10 07:22:44
非公有制企業黨建(2020年9期)2020-09-26 13:22:18
世界科學技術-中醫藥現代化(2020年2期)2020-07-25 02:05:36
瘋狂英語·新策略(2019年10期)2019-12-13 08:43:28
當代陜西(2019年10期)2019-06-03 10:12:04
瘋狂英語·新策略(2018年1期)2018-08-20 07:50:38
數學小靈通·3-4年級(2017年9期)2017-10-13 08:10:54
領導決策信息(2017年12期)2017-05-17 04:49:18
中國衛生(2016年7期)2016-11-13 01:06:44
