ptsG/mglB雙基因敲除對大腸桿菌發(fā)酵混合糖產(chǎn)L-乳酸的影響
2021-10-18 13:06:32劉汝婷郭西鵬王金華
中國釀造 2021年9期
關(guān)鍵詞:利用
劉汝婷,張 倩,郭西鵬,王金華,高 娃*
(湖北工業(yè)大學 生物工程與食品學院 發(fā)酵工程教育部重點實驗室,湖北 武漢 430068)
L-乳酸是一種重要的有機酸,廣泛應(yīng)用于食品、醫(yī)藥和化工等領(lǐng)域。作為可生物降解的新材料——聚乳酸(polylactic acid,PLA)的主要原料,L-乳酸已經(jīng)成為目前最重要的有機酸之一[1-2]。由于其居高不下的生產(chǎn)成本,難以在價格上與傳統(tǒng)的塑料相競爭[3-4]。
我國是農(nóng)產(chǎn)品種植大國,每年產(chǎn)生的農(nóng)作物秸稈還有其他農(nóng)作物廢棄物較多[5]。數(shù)量巨大的小麥秸稈尚未進行大規(guī)模有效的利用與開發(fā),多采用焚燒處理造成環(huán)境污染,利用這些廉價的小麥秸稈可以代替糧食生產(chǎn)乳酸[6-7]。木質(zhì)纖維素經(jīng)過各種處理后,其水解液包含大量的葡萄糖和木糖[8-11]。因碳代謝阻遏效應(yīng)(carbon catabolite repression,CRR)[12-13],即葡萄糖的存在會抑制其他糖的代謝,致使大腸桿菌(Escherichia coli)優(yōu)先利用葡萄糖,葡萄糖耗盡后才可利用其他糖類[14-15],絕大多數(shù)的微生物無法利用水解液中的木糖。
為解除在利用木糖時存在的碳代謝阻遏效應(yīng),有研究表明,可以通過基因工程技術(shù)對微生物進行改造[16],該阻遏效應(yīng)與磷酸烯醇式丙酮酸-糖磷酸轉(zhuǎn)移酶(phosphoenolpyruvate-sugarphos photransferase,PTS)系統(tǒng)密切相關(guān)[17]。PTS系統(tǒng)主要負責特異性地轉(zhuǎn)運葡萄糖,同時將葡萄糖轉(zhuǎn)化為葡萄糖-6-磷酸,進入糖酵解過程[18]。有研究報道,敲除PTS系統(tǒng)中的葡萄糖轉(zhuǎn)運酶基因ptsG來降低碳代謝阻遏效應(yīng),進而提高木糖的利用程度,提高糖酸轉(zhuǎn)化效率;半乳糖的相關(guān)轉(zhuǎn)運系統(tǒng)也可以轉(zhuǎn)運葡萄糖,通過敲除半乳糖轉(zhuǎn)運基因mglB,也可降低碳代謝阻遏效應(yīng),使重組大腸桿菌能夠同時利用葡萄糖和木糖[19],如丁小云等[20]構(gòu)建的葡萄糖轉(zhuǎn)運酶基因ptsG缺失乳酸工程菌可同時利用葡萄糖和木糖發(fā)酵產(chǎn)D-乳酸,產(chǎn)量為83.04 g/L;江吉雄[21]通過敲除半乳糖轉(zhuǎn)運基因mglB,降低了混合糖發(fā)酵D-乳酸中的葡萄糖效應(yīng),使發(fā)酵周期縮短了約40%,轉(zhuǎn)化率提高了2.3%;許瓊丹等[18]通過敲除mglB基因,降低了混合糖發(fā)酵乙醇中的葡萄糖效應(yīng),使發(fā)酵周期縮短了約36%,轉(zhuǎn)化率提高了5.8%。
本研究以可高效利用葡萄糖產(chǎn)L-乳酸的大腸桿菌(E.coli)JH16為出發(fā)菌株,通過Red同源重組技術(shù)構(gòu)建ptsG/mglB雙基因缺陷菌株JH2705,以減弱葡萄糖效應(yīng),提高混合糖中木糖的利用率,為基于木質(zhì)纖維素等可再生原料高效生產(chǎn)L-乳酸提供技術(shù)參考。
1 材料與方法
1.1 材料與試劑
1.1.1 菌株與質(zhì)粒
本研究所用的菌株和質(zhì)粒見表1。

表1 本研究所用的菌株和質(zhì)粒Table 1 Stains and plasmids used in this study
1.1.2 試劑
CaCl2·2H2O、L-阿拉伯糖(均為分析純):國藥集團化學試劑有限公司;DL5 000脫氧核糖核酸(deoxyribonucleic acid,DNA)Marker、聚合酶鏈式反應(yīng)(polymerase chain reaction,PCR)Master Mix:美國Fermentas公司;10×TE溶液、氨芐青霉素、卡那霉素(純度均為99%):法國Mersco公司;PCR引物:上海生工生物工程技術(shù)服務(wù)有限公司。
1.1.3 培養(yǎng)基
LB液體培養(yǎng)基[16]:1%胰蛋白胨,0.5%酵母粉,0.5%NaCl。LB固體培養(yǎng)基在LB液體培養(yǎng)基中添加1.5%~2%瓊脂粉。
抗性篩選培養(yǎng)基[16]:LB固體培養(yǎng)基中添加50 μg/mL氨芐青霉素或卡那霉素。
種子培養(yǎng)基[16]:LB液體培養(yǎng)基中加入20 g/L葡萄糖或木糖。
發(fā)酵培養(yǎng)基[16]:LB液體培養(yǎng)基加入56 g/L葡萄糖和24 g/L木糖。
上述培養(yǎng)基均在121 ℃條件下滅菌20 min。
1.2 儀器與設(shè)備
MicroPluser電轉(zhuǎn)儀、My Cycler PCR儀:美國Bio-Red公司;Waters e2695型高效液相色譜(high performance liquid chromatography,HPLC)儀:美國Waters公司;Sartorius BB-8846880發(fā)酵罐:德國Sartorius Stedim Biotech公司。
1.3 方法
1.3.1 PCR引物設(shè)計
根據(jù)ptsG和mglB基因序列設(shè)計敲除引物分別為△ptsGP1、△ptsG-P2、△mglB-P1、△mglB-P2,結(jié)果見表2。該對引物5'端堿基長度為45 bp的基因片段與ptsG和mglB基因序列同源,以18~20 bp(表中下劃線序列)與質(zhì)粒pkD4上FRT-kan-FRT閱讀框序列同源。
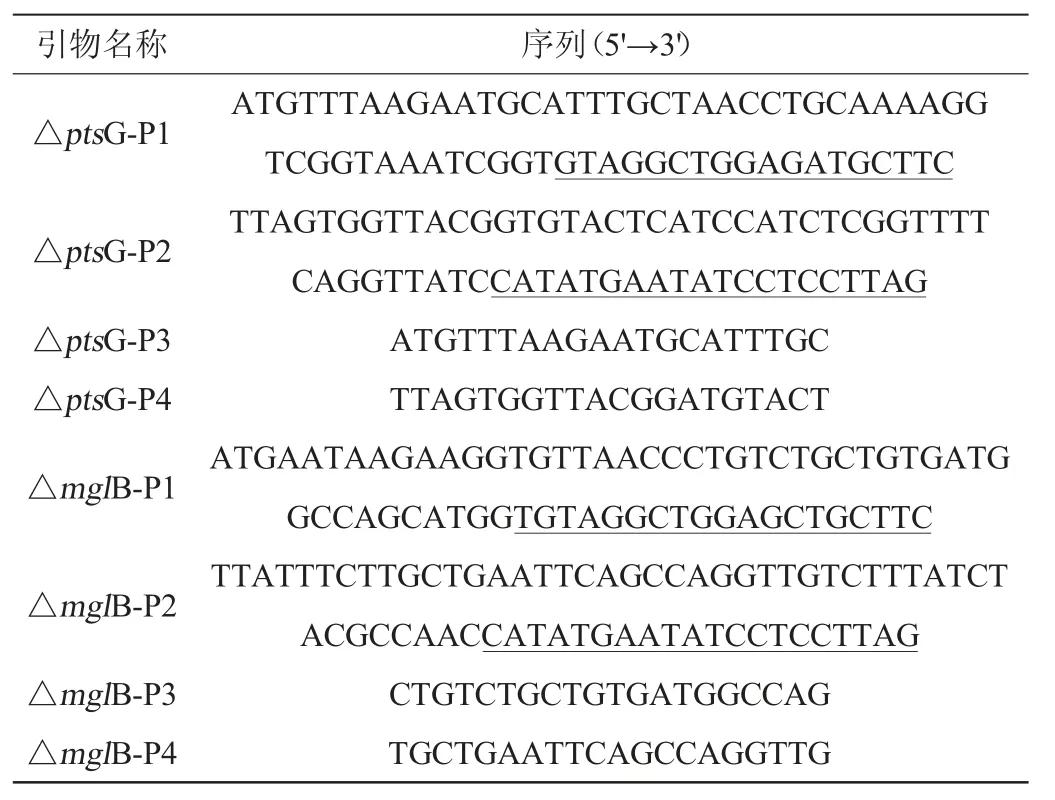
表2 本研究所用引物Table 2 Primes used in the study
1.3.2ptsG/mglB雙基因敲除大腸桿菌的構(gòu)建
以質(zhì)粒pkD4為模板,△ptsG-P1、△ptsG-P1為引物進行PCR擴增得到帶有卡那霉素抗性基因的PCR擴增產(chǎn)物。PCR擴增條件:95 ℃預(yù)變性3 min;95 ℃變性30 s,55 ℃退火30 s,72 ℃延伸2 min,循環(huán)30次。
采用CaCl2法[22]將質(zhì)粒pkD46轉(zhuǎn)化入E.coliJH16的細胞中,通過氨芐青霉素抗性篩選培養(yǎng)基上篩選得到陽性單菌落。將陽性單菌落接種于含有1.6%L-阿拉伯糖的LB培養(yǎng)基中,30 ℃條件下培養(yǎng)至OD600nm值=0.3~0.6,冰水浴30 min后,用去離子水洗滌4次后得到感受態(tài)細胞E.coliJH16/pkD46。
采用電轉(zhuǎn)法[19]將PCR擴增產(chǎn)物轉(zhuǎn)化到感受態(tài)細胞E.coliJH16/pkD46中,涂布到卡那霉素抗性篩選培養(yǎng)基上。在37 ℃培養(yǎng)箱中培養(yǎng)24 h,挑選生長良好的陽性單克隆在卡那霉素抗性篩選培養(yǎng)基上轉(zhuǎn)接1~2代,并用鑒定引物△ptsG-P3、△ptsG-P4進行PCR驗證。將成功敲除ptsG基因的E.coliJH16命名為E.coliJH17。
敲除mglB基因方法與敲除ptsG基因方法相同。將成功敲除mglB基因的E.coliJH17命名為E.coliJH2705。
1.3.3 發(fā)酵搖瓶培養(yǎng)
挑取E.coliJH2705單菌落接種于LB液體培養(yǎng)基中,裝液量為100 mL/250 mL,37 ℃、200 r/min條件下培養(yǎng)至OD600nm值至1.0~1.2左右,作為種子液。按2%(V/V)的接種量將種子液接種于發(fā)酵培養(yǎng)基中,裝液量為100 mL/250 mL,37 ℃、200 r/min條件下培養(yǎng),定時取樣,測定菌體濃度光密度值(OD600nm值)、葡萄糖、木糖殘留量及乳酸產(chǎn)量,每個處理作三個平行。
1.3.4 發(fā)酵罐發(fā)酵培養(yǎng)
一級種子液培養(yǎng):挑取E.coliJH2705單菌落接種于LB液體培養(yǎng)基中,裝液量為50 mL/250 mL,37 ℃、100 r/min條件下培養(yǎng)12 h。二級種子液培養(yǎng):取20 mL一級種子液接種于LB液體培養(yǎng)基中,裝液量為400 mL/1 L,37 ℃、150 r/min條件下培養(yǎng)到OD600nm值為1.0左右。將培養(yǎng)好的二級種子液按10%(V/V)的接種量接種至發(fā)酵培養(yǎng)基中,裝液量為4 L/7 L發(fā)酵罐,37 ℃、200 r/min條件下發(fā)酵72 h,發(fā)酵過程中采用3 mol/L的Ca(OH)2作為中和劑控制發(fā)酵液pH值為7.0。定時取樣,測定菌體濃度OD600nm值、葡萄糖、木糖殘留量及L-乳酸產(chǎn)量。
1.3.5 分析檢測
菌體濃度OD600nm值:采用紫外分光光度計在波長600 nm處測定菌液的吸光度值;葡萄糖、L-乳酸含量:采用生物傳感儀檢測法測定[23];木糖含量:采用高效液相色譜分析[19]。
2 結(jié)果與分析
2.1 重組菌大腸桿菌JH2705的構(gòu)建結(jié)果
以鑒定引物△ptsG-P3和△ptsG-P4、△mglB-P3、△mglBP4對磷酸轉(zhuǎn)移酶系統(tǒng)中的葡萄糖轉(zhuǎn)運酶基因ptsG(堿基長度為950 bp)和半乳糖轉(zhuǎn)運基因mglB(堿基長度為910 bp)的敲除結(jié)果進行驗證,結(jié)果見圖1。由圖1可知,E.coliJH16經(jīng)過菌落PCR擴增后,出現(xiàn)堿基長度分別為950 bp和910 bp的基因片段,而敲除這兩個基因后的重組菌E.coliJH2705經(jīng)菌落PCR擴增后,無ptsG、mglB基因片段,說明重組菌E.coliJH2705中的ptsG、mglB基因已經(jīng)成功被敲除。
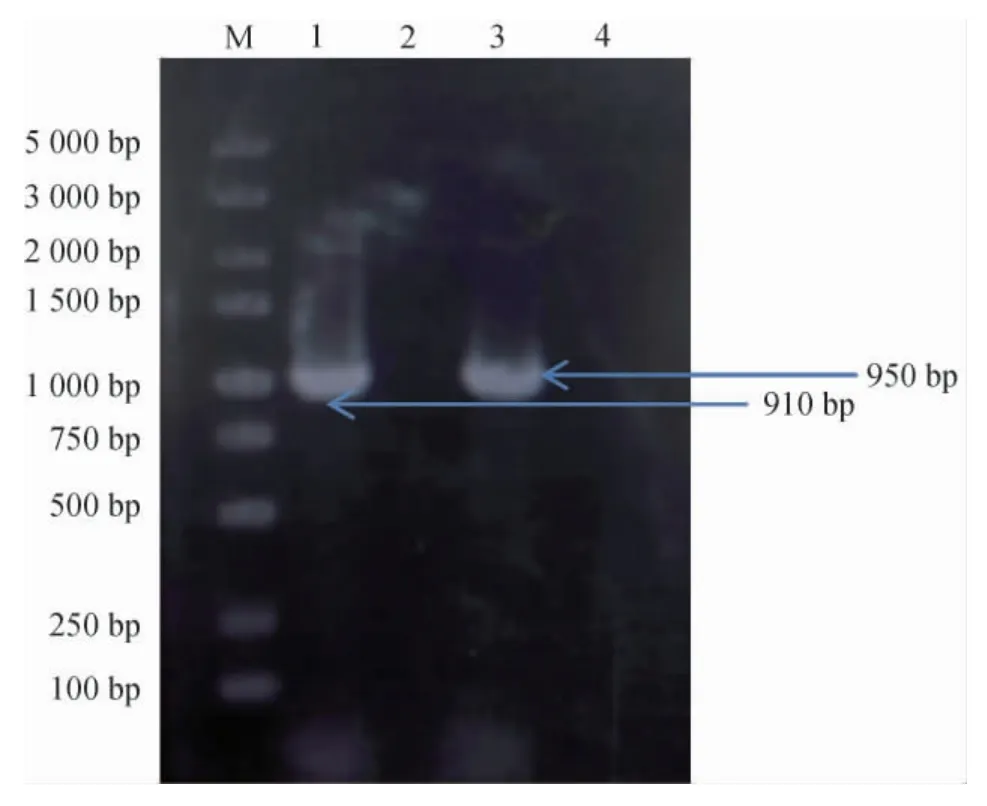
圖1 ptsG/mglB基因缺陷菌株Escherichia coli JH2705的PCR鑒定結(jié)果Fig.1 PCR identification result of ptsG and mglB genes deficient Escherichia coli JH2705
2.2 搖瓶發(fā)酵試驗結(jié)果
以小麥秸稈為代表的木質(zhì)纖維素水解液中,由纖維素水解得到葡萄糖約為60%,且半纖維素與木糖比大約為7∶3[24],因此,本研究模擬小麥秸稈水解液組成,設(shè)定5.6%葡萄糖與2.4%木糖為發(fā)酵碳源,不同菌株對混合糖的發(fā)酵效果見表3。由表3可知,E.coliJH16、E.coliJH17、E.coliJH2705三菌株在72 h內(nèi)可發(fā)酵完畢,與出發(fā)菌E.coliJH16相比,E.coliJH2705的葡萄糖殘?zhí)橇浚?8.7 g/L)增加,葡萄糖消耗速率(0.52 g/(L·h))降低33.3%,無木糖殘留,木糖消耗速率(0.36 g/(L·h))升高38.5%,乳酸生產(chǎn)強度(0.56 g/(L·h))提高33.3%。說明敲除ptsG和mglB基因后,碳代謝阻遏效應(yīng)減弱,提高木糖消耗速率。

表3 Escherichia coli JH16、JH17、JH2705對混合糖發(fā)酵結(jié)果Table 3 Mixed sugar fermentation results of Escherichia coli JH16,JH17 and JH2705
2.3 發(fā)酵罐發(fā)酵試驗結(jié)果
2.3.1 生長曲線
E.coliJH16、E.coliJH17、E.coliJH2705在7 L發(fā)酵罐中發(fā)酵利用混合糖時的生長情況見圖2。由圖2可知,E.coliJH16、E.coliJH17及E.coliJH2705分別在發(fā)酵24 h、28 h、32 h時,OD600nm值達到最大(7.1、7.4、8.4)后進入穩(wěn)定期。與E.coliJH16相比,E.coliJH17和E.coliJH2705的菌體生長更為明顯,表明敲除基因后的兩菌株可以攝取更多的碳源用于菌體生長[25]。
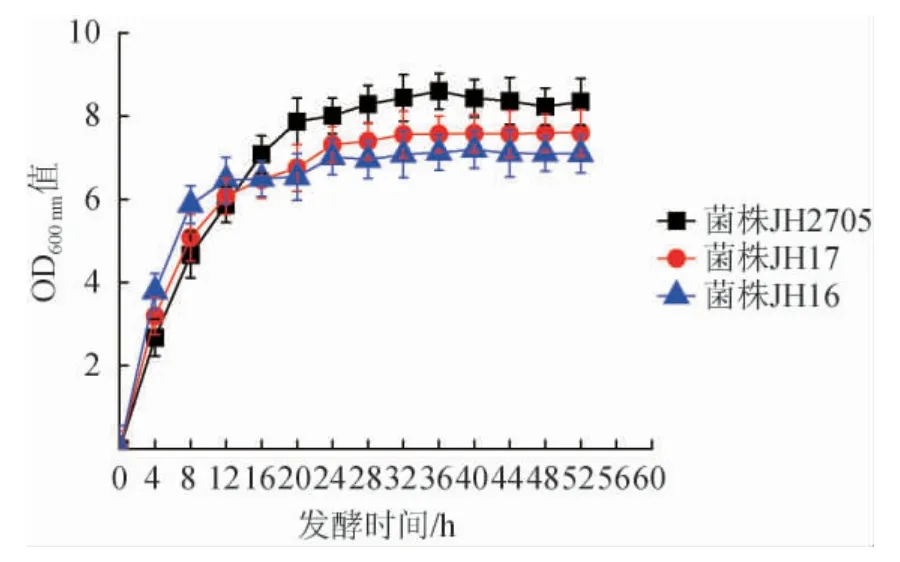
圖2 Escherichia coli JH16、JH17及JH2705發(fā)酵利用混合糖時的生長曲線Fig.2 Growth curves of Escherichia coli JH16,JH17 and JH2705 in fermentation with mixed sugars
2.2.2 葡萄糖和木糖消耗曲線
E.coliJH16、E.coliJH17及E.coliJH2705在7 L發(fā)酵罐中發(fā)酵利用混合糖時葡萄糖及木糖的消耗情況見圖3。由圖3可知,E.coliJH2705同時開始利用葡萄糖和木糖,發(fā)酵40 h時木糖消耗完畢,發(fā)酵至44 h時葡萄糖消耗完畢,而E.coliJH16優(yōu)先利用葡萄糖,發(fā)酵28 h時葡萄糖利用完畢,發(fā)酵30 h時開始利用木糖,至52 h還剩余7.1 g/L未被利用;E.coliJH17同時利用葡萄糖和木糖,發(fā)酵44 h時葡萄糖利用完畢,發(fā)酵至52 h時木糖剩余量為3.1 g/L。E.coliJH16、E.coliJH17、E.coliJH2705消耗葡萄糖的速率分別為2.01 g/(L·h)、1.28g/(L·h)、1.20g/(L·h),消耗木糖速率分別為0.33 g/(L·h)、0.40 g/(L·h)、0.60 g/(L·h)。結(jié)果表明,ptsG基因的缺失,可有效降低分解代謝阻遏效應(yīng),使得葡萄糖消耗速率降低,木糖消耗速率升高。而ptsG、mglB基因雙缺陷菌E.coliJH2705的木糖消耗速率比ptsG基因缺陷菌E.coliJH17提高50%,表明兩個基因?qū)δ咎抢眯噬呔哂携B加效應(yīng)。

圖3 Escherichia coli JH16、JH17及JH2705發(fā)酵利用混合糖時木糖和葡萄糖消耗曲線Fig.3 Curves of xylose and glucose consumption in Escherichia coli JH16,JH17 and JH2705 fermentation with mixed sugars
2.2.3L-乳酸產(chǎn)量
E.coliJH16、E.coliJH17、E.coliJH2705的L-乳酸產(chǎn)量見圖4。由圖4可知,在發(fā)酵16 h之前,E.coliJH16的L-乳酸產(chǎn)量高于E.coliJH17、E.coliJH2705,E.coliJH17、E.coliJH2705同時利用葡萄糖和木糖產(chǎn)L-乳酸,但利用木糖的速率較慢,而E.coliJH16雖然不能利用木糖,但其利用葡萄糖產(chǎn)L-乳酸的速率卻大于E.coliJH17、E.coliJH2705利用葡萄糖和木糖產(chǎn)L-乳酸的速率之和;在發(fā)酵18 h后,E.coliJH17和E.coliJH2705的L-乳酸產(chǎn)量逐漸超過E.coliJH16,E.coliJH17和E.coliJH2705利用木糖產(chǎn)L-乳酸的能力遠大于E.coliJH16。發(fā)酵至48 h時,E.coliJH16的L-乳酸產(chǎn)量為34.12 g/L,轉(zhuǎn)化率為42.0%,生產(chǎn)強度為0.71 g/(L·h);E.coliJH17的L-乳酸產(chǎn)量為53.22 g/L,轉(zhuǎn)化率為66.5%,生產(chǎn)強度為1.02 g/(L·h);E.coliJH2705的L-乳酸產(chǎn)量為56.03 g/L,轉(zhuǎn)化率為70.0%,生產(chǎn)強度為1.27 g/(L·h)。與出發(fā)菌株E.coliJH16相比,重組菌株E.coliJH17、JH2705生產(chǎn)強度分別提高了43.9%、79.1%。重組菌株E.coliJH2705已消耗完所有碳源,而E.coliJH17、JH16分別剩余3.1 g/L、7.1 g/L的木糖。結(jié)果表明,敲除ptsG和mglB基因后,菌株的生產(chǎn)強度提高。
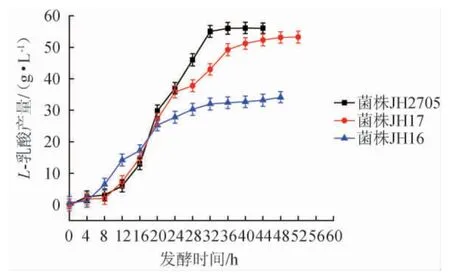
圖4 Escherichia coli JH16、JH17及JH2705發(fā)酵混合糖產(chǎn)L-乳酸含量Fig.4 Contents of L-lactic acid produced by Escherichia coli JH16,JH17 and JH2705 fermented with mixed sugars
3 結(jié)論
本研究以大腸桿菌JH16為出發(fā)菌株,通過Red同源重組技術(shù),同時敲除了磷酸轉(zhuǎn)移酶系統(tǒng)中的葡萄糖轉(zhuǎn)運酶基因ptsG和半乳糖轉(zhuǎn)運基因mglB,構(gòu)建ptsG/mglB雙基因缺陷型大腸桿菌JH2705。該菌株以8%混合糖(5.6%葡萄糖+2.4%木糖)為碳源發(fā)酵產(chǎn)L-乳酸時,能同時利用葡萄糖和木糖,大幅度減小了葡萄糖效應(yīng)所帶來的不利影響,其木糖利用速率為0.60 g/(L·h),L-乳酸生產(chǎn)強度為1.27 g/(L·h),較出發(fā)菌株E.coliJH16分別提高50%、79.1%,并且E.coliJH2705的糖酸轉(zhuǎn)化率高達70%,為木質(zhì)纖維度高效利用木糖提供一定的理論基礎(chǔ)。
猜你喜歡
中等數(shù)學(2022年2期)2022-06-05 07:10:50
中學生數(shù)理化·七年級數(shù)學人教版(2021年11期)2021-12-06 05:38:48
中學生數(shù)理化(高中版.高考數(shù)學)(2021年6期)2021-07-28 06:19:08
小學生學習指導(dǎo)(低年級)(2020年6期)2020-07-25 02:31:36
小學生學習指導(dǎo)(低年級)(2019年11期)2019-11-25 07:31:44
小學生學習指導(dǎo)(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
數(shù)學小靈通·3-4年級(2017年6期)2017-06-22 11:28:50
工業(yè)設(shè)計(2016年5期)2016-05-04 04:00:33
河北遙感(2015年4期)2015-07-18 11:05:06
