新疆某地區(qū)犬流感病毒血清學(xué)及病原學(xué)調(diào)查
2021-10-20 06:26:04米曉云盛肖剛吳建勇汪萍楊學(xué)云魏玉榮
塔里木大學(xué)學(xué)報(bào) 2021年3期
米曉云,盛肖剛,吳建勇,汪萍,楊學(xué)云,魏玉榮*
(1新疆畜牧科學(xué)院獸醫(yī)研究所,新疆 烏魯木齊,830013)
(2新疆動(dòng)物疫病研究重點(diǎn)實(shí)驗(yàn)室,新疆 烏魯木齊 830013)
(3新疆維吾爾自治區(qū)動(dòng)物衛(wèi)生監(jiān)督所,新疆 烏魯木齊830011)
犬流感(canine Influenza,CI)是一種由犬流感病毒(Canine Influenza Virus,CIV)引起的急性接觸性呼吸道傳染病。犬是流感病毒生態(tài)循環(huán)中的重要中間宿主,馬流感病毒(H3N8亞型)、禽流感病毒(H3N2亞型、H5N1亞型、H5N2亞型和H10N8亞型)和人流感病毒(H1N1亞型)等均可感染犬[1-6]。犬H3N2亞型流感毒株2006年由中國(guó)分離并報(bào)道,其基因序列與韓國(guó)水鳥的禽源H3N2基因序列高度同源[7-8]。隨后研究報(bào)道,犬H3N2亞型流感在中國(guó)、韓國(guó)、泰國(guó)及美國(guó)等地流行[9-12]。盡管目前尚未有CIV跨種感染人的報(bào)道,但犬的流感病毒中間宿主作用和“病毒混合容器”角色,注定無法忽視它在流感病毒生態(tài)循環(huán)中的重要作用,及其對(duì)人類公共衛(wèi)生安全造成潛在的威脅。為了解烏魯木齊地區(qū)流浪犬和寵物犬群體中流感現(xiàn)狀,評(píng)估其對(duì)公共衛(wèi)生危害性,本研究對(duì)部分流浪犬和寵物犬進(jìn)行了犬流感病毒血清學(xué)和病原學(xué)調(diào)查。
1 材料與方法
1.1 被檢樣本
流浪犬基地流浪犬血清樣本和鼻拭子樣本各100份采集于2019年1月;寵物店就診寵物犬血清樣本150份和鼻拭子樣本79份收集于2019年1月至2019年9月。鼻拭子樣本放置于含500 μL PBS緩沖液(pH 7.2)的2 mL離心管中。所有被檢樣本-20℃保存?zhèn)溆茫y(tǒng)一檢測(cè)。
1.2 主要試劑
DL 2000 DNA Marker和TakaRa PrimeScriptTM One Stemp RT-PCR Kit Ver.2購(gòu)自購(gòu)自 TakaRa;ID Sreen?influenza A Antibody Comptition Multi-species試劑盒購(gòu)自ID VET;TIANamp Virus RNA Kit購(gòu)自天根生化科技(北京)有限公司。
1.3 引物
根據(jù)NY/T-772-2013合成流感檢測(cè)通用引物M229[13],根據(jù)文獻(xiàn)合成擴(kuò)增 CIV 檢測(cè)引物 cH3N2-602F和cH3N2-1145R[14]。參照GenBank序列(登錄號(hào):MK212413、MK212411)設(shè)計(jì)HA和NA基因引物,見表1。引物由新疆昆泰銳生物技術(shù)有限公司合成。

表1 引物序列
1.4 CIV血清學(xué)檢測(cè)
-20℃保存的鼻拭子樣本56℃滅活處理30 min后加入直徑3 mm鋼珠3粒,將離心管置于Tissuelyser-LT破碎儀中進(jìn)行震蕩5次,50 Hz,30 s/次。震蕩結(jié)束后離心管至4℃離心機(jī)中8000 rmp離心5 min,取離心管中上清液提取病毒RNA,保存于-80℃?zhèn)溆谩IV RNA提取操作按TIANamp Virus RNA Kit說明書于生物安全柜中進(jìn)行。
1.5 CIV基因擴(kuò)增
以提取的病毒RNA為模板,先擴(kuò)增M基因,檢測(cè)是否流感病原學(xué)陽性。具體按TakaRa Prime-ScriptTM One Stemp RT-PCR Kit Ver.2說明書操作,在0.2 ml PCR管中加反應(yīng)體系共50 μL:2×Onestep RT-PCR buffer 25 μL,酶混合液4 μL,M-229 U/M-229 L(20 pmol/μL)4 μL,無RNA水14 μL,RNA模板3 μL;其中陰性對(duì)照模板為3 μL無RNA水。反應(yīng)參數(shù)為:42℃ 30 min;94℃ 5 min;94 ℃ 45 s,55℃ 45 s,72 ℃ 30 s,30 cycles,72 ℃保溫10 min。PCR擴(kuò)增結(jié)束后,取產(chǎn)物10 μL瓊脂糖凝膠電泳鑒定。有特異性條帶的擴(kuò)增產(chǎn)物送生工生物工程(上海)股份有限公司測(cè)序。測(cè)序?yàn)镃IV M基因的樣本,按上述方法,用引物cH3N2-602F/cH3N2-1145R擴(kuò)增,反應(yīng)參數(shù)為:42 ℃ 30 min;94 ℃ 5min;94 ℃ 30 s,57 ℃ 30 s,72℃ 35 s,30 cycles,72℃保溫10 min。PCR擴(kuò)增結(jié)束后,取產(chǎn)物10 μL瓊脂糖凝膠電泳鑒定。有特異性條帶的樣品,分別再用引物cH3N2-HA-l/cH3N2-HA-1701R和cH3N2-NA-l/cH3N2-NA-1403R擴(kuò)增流感HA基因和NA基因,反應(yīng)參數(shù)為:42℃30 min;94 ℃ 5 min;94 ℃ 30 s,55 ℃ 30 s,72 ℃ 90 s,30 cycles,72 ℃保溫10 min。PCR擴(kuò)增結(jié)束后,產(chǎn)物10 μL經(jīng)瓊脂糖凝膠電泳鑒定。擴(kuò)增產(chǎn)物再次送生工生物工程(上海)股份有限公司測(cè)序。
1.6 序列比較分析
測(cè)序序列經(jīng)NCBI中BLASTn比對(duì)確認(rèn),用Lasergenev7.1 DNAStar軟件進(jìn)行序列整理、校對(duì)和拼接,利用MEGA5.0軟件繪制系統(tǒng)進(jìn)化樹。
2 結(jié)果
2.1 犬流感血清學(xué)結(jié)果
采集的100份流浪犬血清樣本和150份寵物犬血清樣本進(jìn)行ELISA檢測(cè),結(jié)果顯示,流浪犬CIV抗體陽性率58%(58/100),寵物犬CIV抗體陽性率5.33%(8/150),見表2。
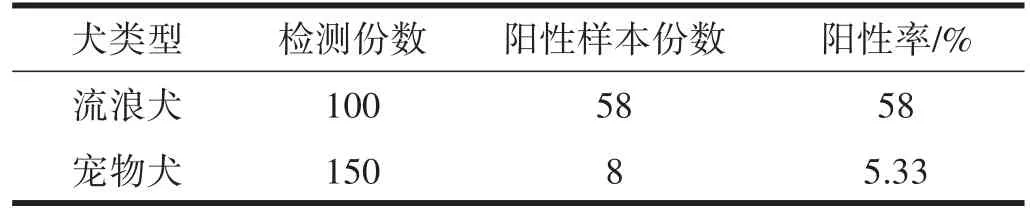
表2 犬血清流感抗體ELISA方法檢測(cè)結(jié)果
2.2 CIV病原學(xué)結(jié)果
2.2.1 CIV基因擴(kuò)增結(jié)果
M基因擴(kuò)增與測(cè)序鑒定,以CIV鼻拭子樣本RNA為模板,One-step RT-PCR方法擴(kuò)增目的基因,電泳結(jié)果顯示100份流浪犬中有4份樣本的PCR產(chǎn)物可見約229 bp條帶,寵物犬79份樣本中有1份PCR產(chǎn)物大小與預(yù)期大小相符,見圖1。有特異性條帶的PCR產(chǎn)物純化后送基因公司測(cè)序,經(jīng)BLASTn比對(duì),所測(cè)序列均為CIV基因。
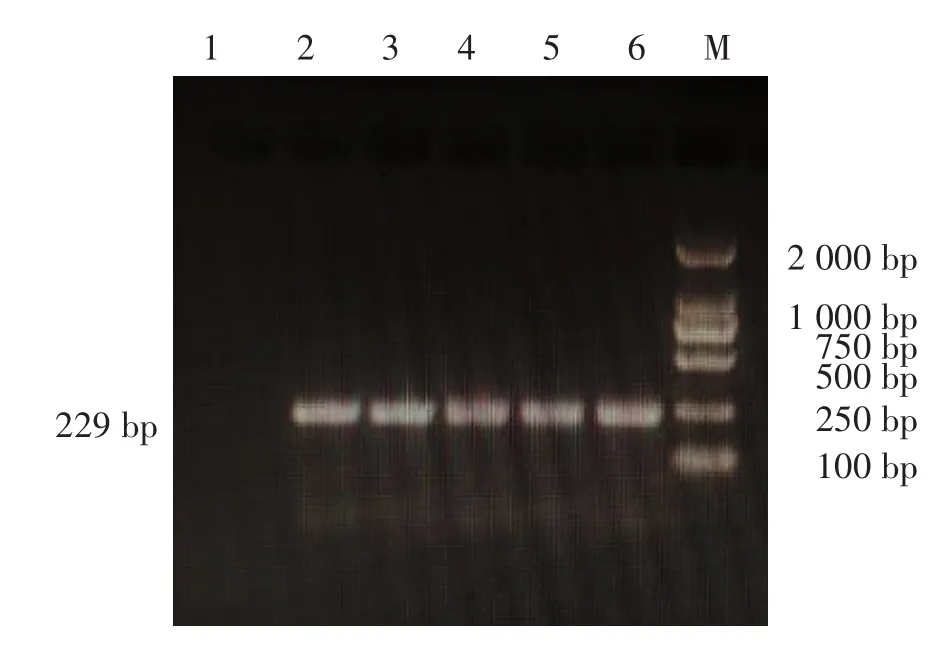
圖1 鼻拭子樣本M基因擴(kuò)增結(jié)果
H3N2亞型CIV的初步鑒定,將測(cè)序鑒定為CIV的RNA樣本,再次用引物cH3N2-602F/cH3N2-1145R擴(kuò)增,均可見預(yù)期540 bp的特異性條帶,見圖2。

圖2 鼻拭子樣本H3N2亞型初步鑒定結(jié)果
HA和NA基因擴(kuò)增與測(cè)序鑒定,經(jīng)引物cH3N2-602F/cH3N2-1145初步鑒定為H3N2亞型的2份流浪犬RNA樣本和1份寵物犬RNA樣本再次擴(kuò)增HA和NA基因,電泳結(jié)果顯示3份樣本均有1701 bp和1403 bp的預(yù)期大小特異性條帶,見圖3和圖4。

圖3 鼻拭子樣本HA基因擴(kuò)增結(jié)果
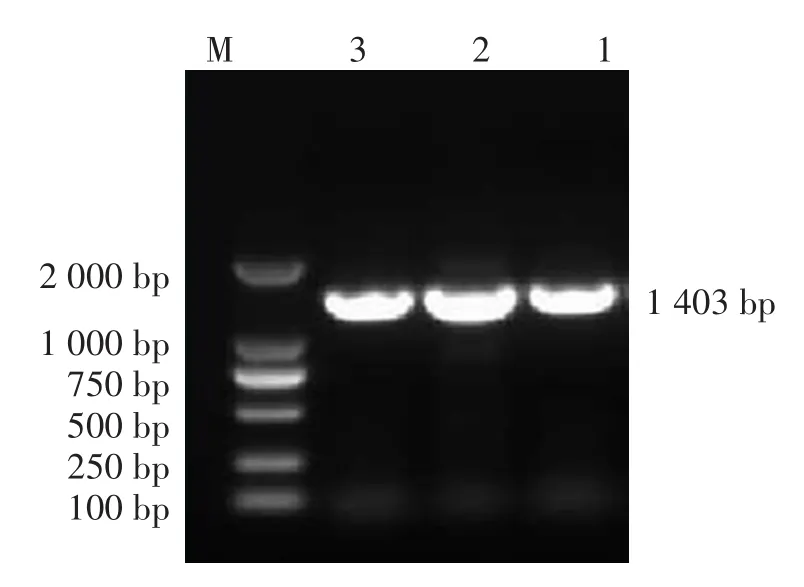
圖4 鼻拭子樣本NA基因擴(kuò)增結(jié)果
2.2.2 HA和NA基因測(cè)序分析
測(cè)序序列經(jīng)BLASTn比對(duì)確認(rèn),用Lasergenev 7.1 DNAStar軟件進(jìn)行序列整理、校對(duì)和拼接,拼接的核苷酸序列再次進(jìn)行BLASTn比對(duì),可見拼接序列與犬流感H3N2亞型同源性最高,將其分別命名為A/canine/Xinjiang/1/2019(H3N2)、A/canine/Xinjiang/2/2019(H3N2)和 A/canine/Xinjiang/3/2019(H3N2)。
2.2.3 HA基因遺傳進(jìn)化分析
運(yùn)用MEGA5.0軟件構(gòu)建基于HA基因的H3N2 CIV遺傳進(jìn)化樹,見圖5。從遺傳關(guān)系可見,2016年后的H3N2 CIV形成一個(gè)獨(dú)立的分支,本研究測(cè)序的3株CIV也處于此分支。分析病毒分子特征,這一分支的HA蛋白的第226位是Q(谷氨酰胺)、第228位是G(甘氨酸)、146位是S(絲氨酸)、第242位是I(異亮氨酸)。而2006年至2015年期間的CIV HA蛋白的第146位是G(甘氨酸)或S(絲氨酸)、第242位是V(頡氨酸)。

圖5 基于HA基因的H3N2亞型流感毒株遺傳進(jìn)化分析
3 討論
2012年至2017采集自北京、南京、上海和西安的犬樣本CIV H3N2抗體陽性率為13.5%(54/399),其中2017年犬H3N2抗體陽性率為6.35%,而2012年至2013年期間H3N2亞型抗體陽性率為3.5%,抗體陽性率的明顯升高表明目前流行的毒株可能更具傳染性[15]。本研究中流浪犬CIV抗體陽性率58%(58/100),寵物犬CIV抗體陽性率5.33%(8/150)。流浪犬CIV抗體陽性率高可能與采樣時(shí)間及采樣群體有關(guān)。流浪犬血清樣本是2019年1月集中采集于流浪犬收養(yǎng)基地,推測(cè)犬群中正在或曾經(jīng)有CIV流行。寵物犬血清采集于寵物醫(yī)院就診犬,時(shí)間相對(duì)分散,樣品相對(duì)較少,無法考察其流感抗體陽性率是否與性別、品種和年齡具相關(guān)性,病原學(xué)陽性率低可能與樣本保存時(shí)間相關(guān)。本研究中病原學(xué)檢測(cè)有5份樣品(5/179)可擴(kuò)增出CIV M基因序列,經(jīng)分型引物初步鑒定為H3N2亞型,有2份流浪犬樣本及1份寵物犬樣本可擴(kuò)增出完整的HA和NA基因序列,測(cè)序?yàn)镃IV H3N2亞型。測(cè)序確認(rèn)的4只CIV抗原陽性流浪犬其血清抗體也是陽性。
HA蛋白負(fù)責(zé)流感病毒與唾液酸受體的結(jié)合、入侵和膜融合,在流感病毒跨種傳播中發(fā)揮重要作用。H3亞型流感病毒,當(dāng)226位為L(zhǎng)(亮氨酸)而228位是S(絲氨酸),HA蛋白結(jié)合人α-2,6受體,而當(dāng)226位是Q(谷氨酰胺)而228位是G(甘氨酸),HA則偏好結(jié)合禽α-2,3受體[16]。LYU Y L等[15]認(rèn)為2016年以后的H3N2 CIV是有別于以前的H3N2亞型并將其取代的新進(jìn)化分支,推測(cè)起源于韓國(guó)或美國(guó)的H3N2 CIV。該分支的HA具有禽α-2,3受體結(jié)合位點(diǎn),同時(shí)在第146和第242位有氨基酸突變。2006-2015年期間的CIV H3N2亞型HA蛋白第146是G(甘氨酸)和S(絲氨酸)的頻率分別為91.89%和8.11%;第242位是V(頡氨酸)的頻率是100%[15]。而2017年至2019年期間的CIV H3N2亞型HA蛋白第146是S(絲氨酸)和第 242位是 I(異亮氨酸)的頻率均是 100%[15]。人H3N2亞型HA蛋白第146是S(絲氨酸)和G(甘氨酸)的頻率分別是99.57%和0.42%;第242位是I(異亮氨酸)和V(頡氨酸)的頻率分別是99.29%和0.39%[17]。由此可見HA第146S和242I的突變有利于CIV在哺乳動(dòng)物中流行。本研究測(cè)序的3株CIV處于2016年后的新進(jìn)化分支,發(fā)生了適應(yīng)哺乳動(dòng)物的HA第146S和242I突變。
寵物犬和流浪犬已逐漸成為流感病毒進(jìn)化和繁衍的“溫床”,在流感病毒生態(tài)循環(huán)中起到重要作用。目前雖未有強(qiáng)毒型CIV的報(bào)道,但是CIV亞型在持續(xù)增加。借鑒H1N1亞型豬流感pdm09病毒跨宿主感染人之前曾在豬體內(nèi)數(shù)十年循環(huán)適應(yīng)事件,2016年后流行于中國(guó)的進(jìn)化新分支CIV H3N2的哺乳動(dòng)物潛在適應(yīng)性及其公共衛(wèi)生安全需進(jìn)一步評(píng)估。我們應(yīng)積極監(jiān)測(cè)以期及時(shí)阻斷CIV的群體傳播和適應(yīng),同時(shí)加強(qiáng)檢疫,避免出現(xiàn)更多新亞型CIV。
4 結(jié)論
2019年新疆某地區(qū)犬樣本CIV血清學(xué)檢測(cè)流浪犬和寵物犬抗體陽性率分別為58%和5.33%;CIV病原學(xué)檢測(cè)流浪犬和寵物犬抗原陽性率分別為4%和1.27%;病原學(xué)測(cè)序分析的3株CIV屬H3N2亞型2016年后新進(jìn)化分支,HA蛋白分析可見第146S和242I突變。HA蛋白中氨基酸哺乳動(dòng)物適應(yīng)突變對(duì)公共衛(wèi)生安全具有潛在威脅。
