
玉米SNAC轉錄因子的基因結構特征與逆境調控預測
2021-11-03 03:30:46羅平龐博崔進鑫于爽王曉楠程名陳勇高文偉郝轉芳
中國農業科技導報 2021年10期
羅平,龐博,崔進鑫,于爽,王曉楠,程名,陳勇,高文偉*,郝轉芳*
(1.新疆農業大學農學院,烏魯木齊 830000;2.中國農業科學院作物科學研究所,北京 100081)
隨著全球氣候變化,降雨量不均衡導致干旱等一系列逆境脅迫的發生。在非生物逆境脅迫中,干旱脅迫是制約農作物生長發育和產量的重要因素[1]。逆境會導植物一系列形態、生理、生化和分子組成的改變[2]。在細胞和分子水平,植物通過ABA途徑和非ABA途徑調控一系列耐旱基因表達來響應干旱脅迫。這些基因不僅通過產生重要的功能蛋白保護細胞避免受到傷害,還通過特定基因調控信號轉導來響應逆境脅迫,比如轉錄因子[3]。轉錄調控是基因表達的重要機制[4],轉錄因子等調控蛋白通過與基因啟動子中的順式元件相互作用,激活或抑制應激相關基因,在應激反應中發揮重要作用[5]。NAC轉錄因子家族是一類植物特有的參與耐逆響應的轉錄因子。NAC家族的命名最早源于牽牛花(Pharbitis)NAM、擬南芥(Arabidopsisthaliana)ATAF1/2和CUC2(cup-shaped cotyledon)基因[6],取三個基因的首字母命名為NAC基因[7]。NAC轉錄因子N端具有由150個氨基酸組成的保守NAC結構域,高度保守,C端的轉錄激活區則高度可變。NAC結構域主要包含5個亞域[8],其中,A、C、D亞域非常保守,而B、E在不同基因中有所差異。NAC轉錄因子C端通常作為轉錄激活物或阻遏物發揮作用,這些蛋白質通過與順式作用的啟動子元件結合而起作用,從而為啟動靶基因的轉錄提供了觸發器[9]。
植物生長和農業生產力不斷受到干旱、高鹽和極端溫度等各種環境因素的威脅。植物已經在分子和生理水平上進化出適應脅迫的有效機制,從而使它們能夠在脅迫條件下生存。近年來,已發現多個NAC轉錄因子直接參與或通過調控參與干旱、高鹽、高溫、冷害等應答基因的表達,在植物非生物逆境脅迫響應中發揮著重要作用[10]。NAC轉錄因子的過表達株系多表現為較強的非生物逆境耐受性,并且可以提高兩種或以上的耐逆境能力。例如,過表達OSNAC10提高了水稻耐旱性[11];過表達SNAC1和ONAC045提高了水稻耐旱和耐鹽性[12-13];過表達SNAC2、OsNAC5、OsNAC6提高了水稻抵御冷、旱和鹽脅迫的能力[14-16]。小麥中過表達TaNAC2L可以提高轉基因擬南芥的耐高溫能力[17];過表達TaNAC2和TaNAC8可以提高轉基因擬南芥抵御非生物的能力[18-19]。有研究在ZmNAC111基因啟動子中插入微型反轉重復轉座因子(MITE),結果顯示,MITE與玉米抗旱性的自然變異顯著相關,通過在擬南芥中異源表達,82 bp 的MITE通過RNA定向DNA甲基化和H3K9二甲基化抑制ZmNAC111的表達,增強了ZmNAC111在轉基因玉米中的表達,提高苗期的耐旱性,提高水分利用效率,誘導干旱響應基因在水分脅迫下的上調[20]。擬南芥中過表達ZmSNAC1(ZmNAC010312)、ZmNAC33(ZmNAC030295)和ZmNAC55(ZmNAC050436)導致種子萌發期對ABA和滲透脅迫的敏感性,提高了擬南芥的耐旱性[21-23]。同時,ZmNAC84在煙草中過表達可以提高轉基因植株耐旱性,減輕干旱誘導的氧化損傷,在此過程中,ABA產生的H2O2首先誘導ZmCCaMK和ZmNAC84表達并激活ZmCCaMK,隨后激活的ZmCCaMK在S113位點磷酸化ZmNAC84,從而通過激活下游基因來誘導抗氧化防御[24]。大豆GmSNAC49在擬南芥中過表達導致干旱和ABA信號通路相關基因上調,提高了干旱脅迫的耐受性[25]。葡萄VvNAC17的過表達增強了擬南芥的抗旱性,同時上調了與ABA和脅迫相關基因的表達[26]。芒屬植物MLNAC10通過調節ABA信號通路發揮抗氧化酶的作用,減輕ROS的損傷,是干旱和耐鹽脅迫的重要調節因子[27]。在耐干旱作物高粱中也有研究表明,13個SbNAC在開花后的干旱脅迫反應中起著積極或消極的轉錄調控作用[28]。
上述研究都表明,NAC基因是參與植物非生物脅迫反應的關鍵調控基因家族之一。玉米是主要的糧、經、飼兼用的作物,對我國農牧業有重要的支撐作用。因此,本研究利用生物信息學手段分析和鑒定玉米SNAC家族基因,系統地對成員的理化性質、基因結構、系統進化、磷酸化位點、啟動子及二級結構等進行預測分析。這些信息將為進一步研究SNAC基因家族的耐逆境功能提供理論參考。
1 材料與方法
1.1 供試材料
16個玉米SNAC轉錄因子蛋白序列從PlantTFDB(http://PlantTFDB.cbi)獲取,命名及分類參照Li等[29]方法。水稻、擬南芥、大豆等已知的與非生物脅迫相關的NAC基因組序列從NCBI數據庫(https://www.ncbi.nlm.nih.gov/genome)下載。
1.2 玉米SNAC的系統發育分析
將SNAC序列通過ClustalX 1.83軟件進行比對。然后使用PhyML軟件(版本2.4.3)[30]中最大似然法對所有對齊的NAC序列進行系統發育分析。通過FigTree1.4.0對進化樹進行可視化[31]。
1.3 玉米SANC蛋白結構分析
通過GSDS工具(http://GSDS.cbi.pku.edu.cn/)對SNAC基因的內含子和外顯子結構圖譜進行可視化。利用ExPASy(https://web.expasy.org/cgi-bin/protparam/protparam)數據庫統計SNAS蛋白的氨基酸數目、理論等電點、分子量、不穩定參數、脂肪族氨基酸指數等理化性質信息,SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsaautomat.pl?page=npsa_sopma.html)軟件對SNAC家族成員的氨基酸序列進行二級結構預測。DNAMAN 軟件對SNAC家族的16個成員進行氨基酸多序列比對分析。利用MEME(http://MEME.nbcr.net/meme3/MEME.html)鑒定SNAC蛋白中存在的保守序列模體。
1.4 玉米SANC基因家族生物信息學預測分析
通過TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM-2.0/)對SNAC家族成員的氨基酸序列進行分析,預測其有無跨膜結構;利用 NetPhos 3.1 Server (http://www.cbs.dtu.dk/services/NetPhos/)軟件對SNAC家族成員的氨基酸序列進行潛在磷酸化位點分析;利用植物順式元件數據庫PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)分析SNAC基因啟動子區的順式作用元件。
2 結果與分析
2.1 玉米SNAC基因家族的系統發育分析
利用PhyML軟件對玉米SNAC以及其他物種中典型的耐脅迫NAC基因進行系統分析,以探討SNAC基因之間的進化關系,結果(圖1)表明,ZmNAC050302與SNAC1、ZmSNAC1(ZmNAC010312)和ZmNAC55(ZmNAC050436)同屬一個亞群,SNAC1、ZmSNAC1和ZmNAC55三個基因已有被證實具有耐脅迫功能。ZmNAC030295與TaNAC6關系較近,屬同一亞群。CarNAC3亞群中ZmNAC100475和ZmNAC080398關系較近,有研究發現,ZmNAC111(ZmNAC100475)基因在玉米中與耐旱相關,與其同一類群的ZmNAC080398可能具有相似的耐逆境機制。
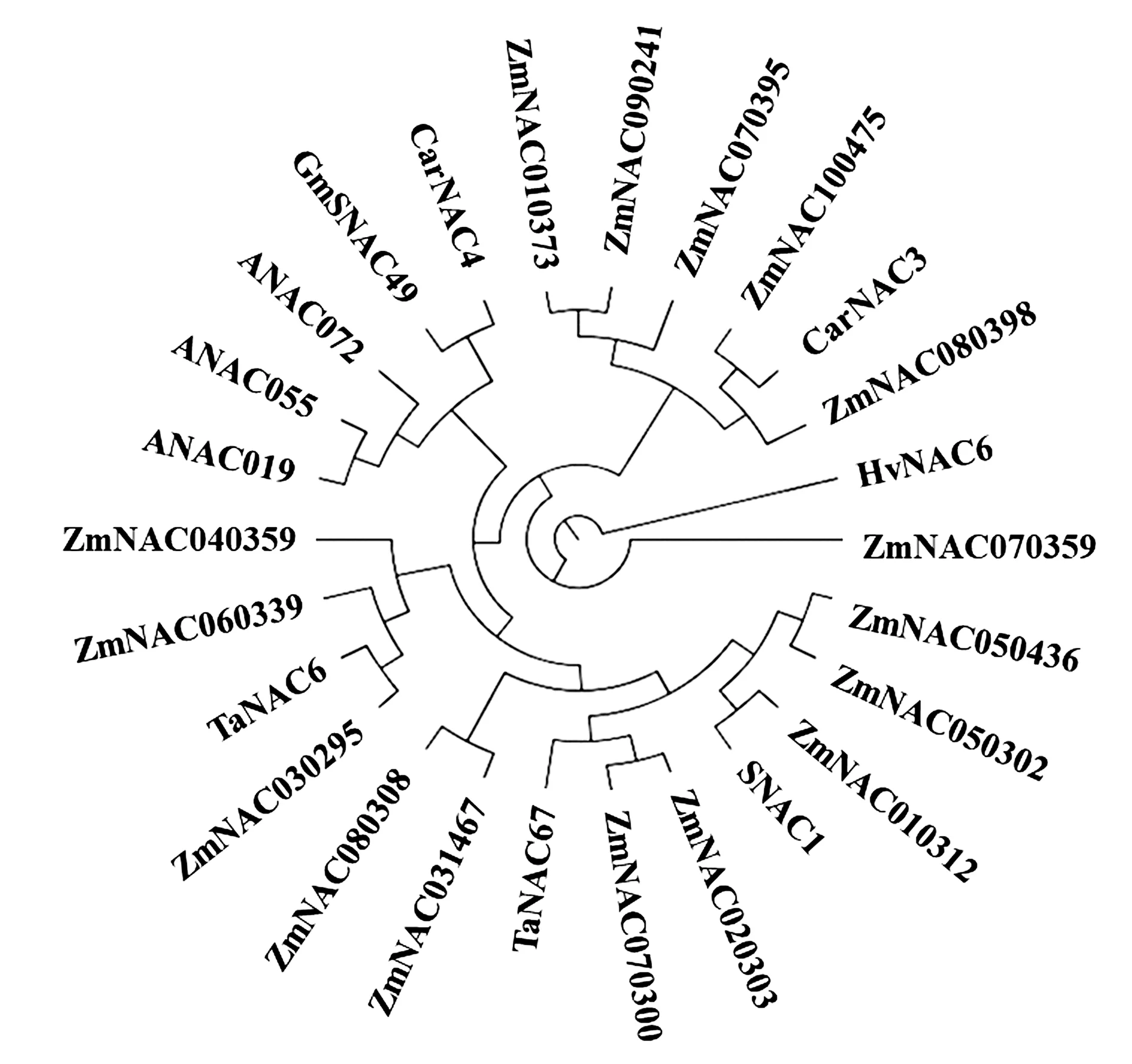
圖1 玉米、擬南芥、水稻、大豆等SNAC基因聚類分析Fig.1 Cluster analysis of SNAC genes in maize,Arabidopsis,rice and soybean
2.2 玉米SNAC基因家族氨基酸序列對比分析
SNAC轉錄因子在逆境脅迫中能提高植物抵抗非生物脅迫的能力,從而增加產量。利用DNAMAN軟件對16 個玉米SNAC 蛋白序列進行多重比對發現,這些 SNAC 蛋白序列的N端均含有高度保守的NAC域,長度約為 150~163 個氨基酸,可以分為5個子域(A~E),其中ZmNAC070395不含有子域A。
2.3 玉米SNAC家族成員理化性質分析
SNAC家族成員的理化性質如表1所示。可以看出,理論等電點最高的是ZmNAC060339,為9.35,而ZmNAC060339的酸性氨基酸的總數比堿性氨基酸的總數少,為堿性蛋白。ZmNAC010373、ZmNAC030295、ZmNAC050436、ZmNAC060339、ZmNAC070359、ZmNAC080308、ZmNAC090241都為堿性蛋白。脂肪族氨基酸指數反映了蛋白質的熱穩定性,分布在57.51(ZmNAC010373)~88.37(ZmNAC031467)之間,總體表明蛋白質間熱穩定性差異較小。其中ZmNAC030295和ZmNAC080398的不穩定參數較高,分別為55.48和53.50。
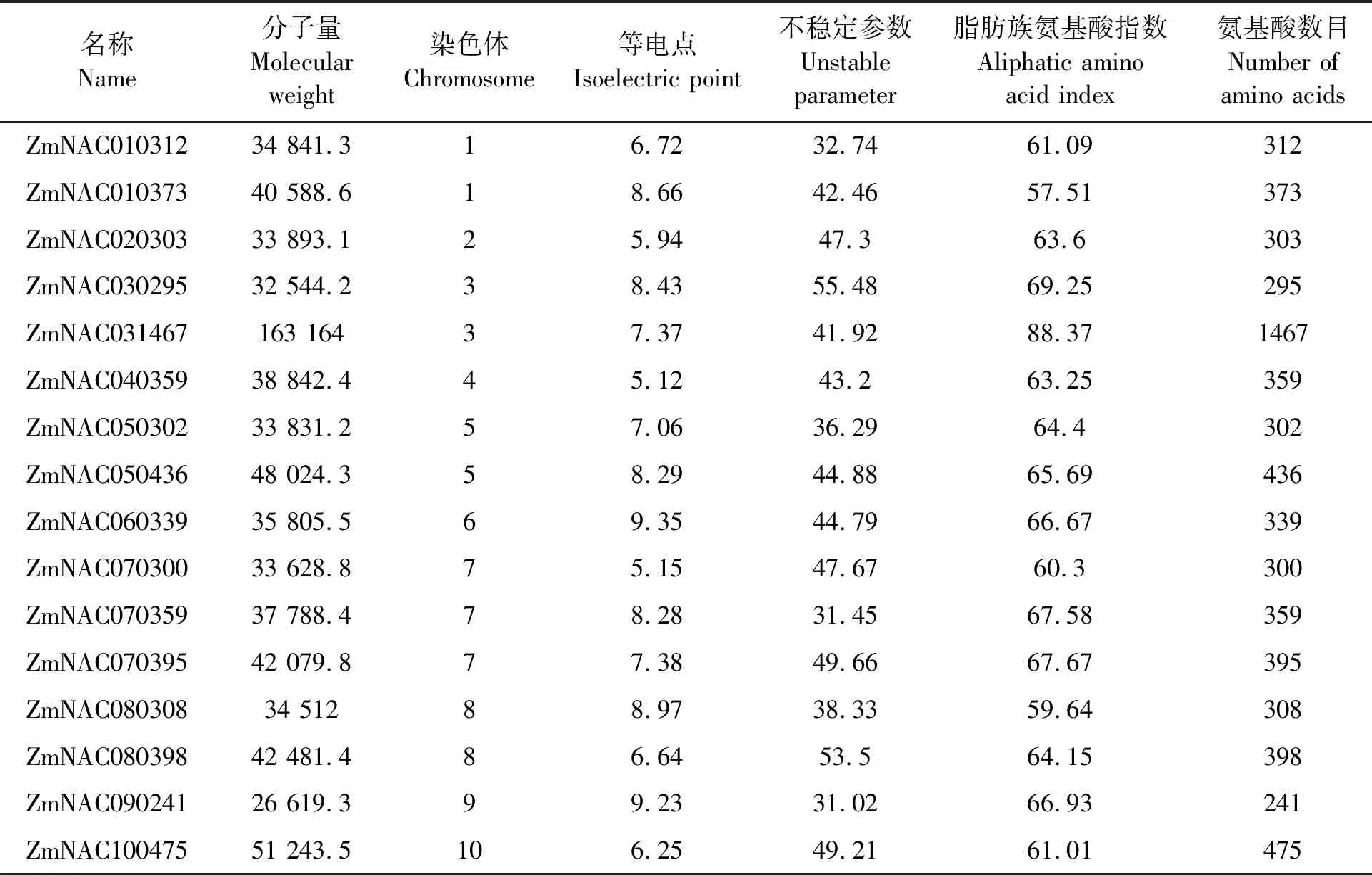
表1 SNAC家族成員的理化性質Table 1 Physico-chemical properties of SNAC family members
2.4 SNAC氨基酸序列跨膜結構預測
TMHMM 綜合了跨膜區疏水性、螺旋長度和膜蛋白拓撲學限制等性質[32]。用 TMHMM-2.0對16個玉米SNAC基因家族成員的氨基酸序列進行跨膜結構域分析,結果表明,這些蛋白不存在跨膜結構域,位于膜外區的可能性較大,為非跨膜蛋白類,推測均在細胞內表達和調控。
2.5 SANC基因家族成員的潛在磷酸化修飾位點預測
磷酸化和去磷酸化在多種真核細胞中的新陳代謝、細胞分裂以及信號轉導等方面都發揮著至關重要的作用[33]。通過NetPhos2.0 Server 軟件對玉米SNAC家族16個成員的氨基酸序列進行潛在磷酸化位點的分析,結果如表2所示。可以看出,玉米SNAC家族16個成員的氨基酸序列均有潛在的磷酸化位點,這些潛在的磷酸化位點分別位于絲氨酸Ser、蘇氨酸Thr和酪氨酸Tyr上,該結果表明,SNAC家族的成員能夠被絲氨酸激酶、蘇氨酸激酶和酪氨酸激酶所磷酸化,從而實現其功能的調控。玉米SNAC家族存在著大量的磷酸化位點,推測SNAC基因家族可能參與著大量的翻譯后修飾。由表中可以看出,SNAC家族成員中ZmNAC031467潛在的磷酸化位點數最多,其次是ZmNAC100475和ZmNAC080398,最少的是ZmNAC020303,只有13個潛在的磷酸化位點。
2.6 SNAC家族成員蛋白質二級結構
采用SOPMA 對SNAC家族成員的氨基酸序列進行蛋白質二級結構的預測,結果如表2和圖2所示。可以看出,組成SNAC家族成員的多肽鏈的二級結構主要有4種類型,分別為α-螺旋(alpha-helix)、β-轉角(beta-turn)、延伸鏈結構(etended strand)以及無規則卷曲(rndom coil),以α-螺旋和無規卷曲為主,并且無規則卷曲占的比例最高,而β-轉角和延伸鏈結構所占的比例則相對較少。對SNAC家族成員二級結構的預測和分析有助于認識蛋白的空間結構。
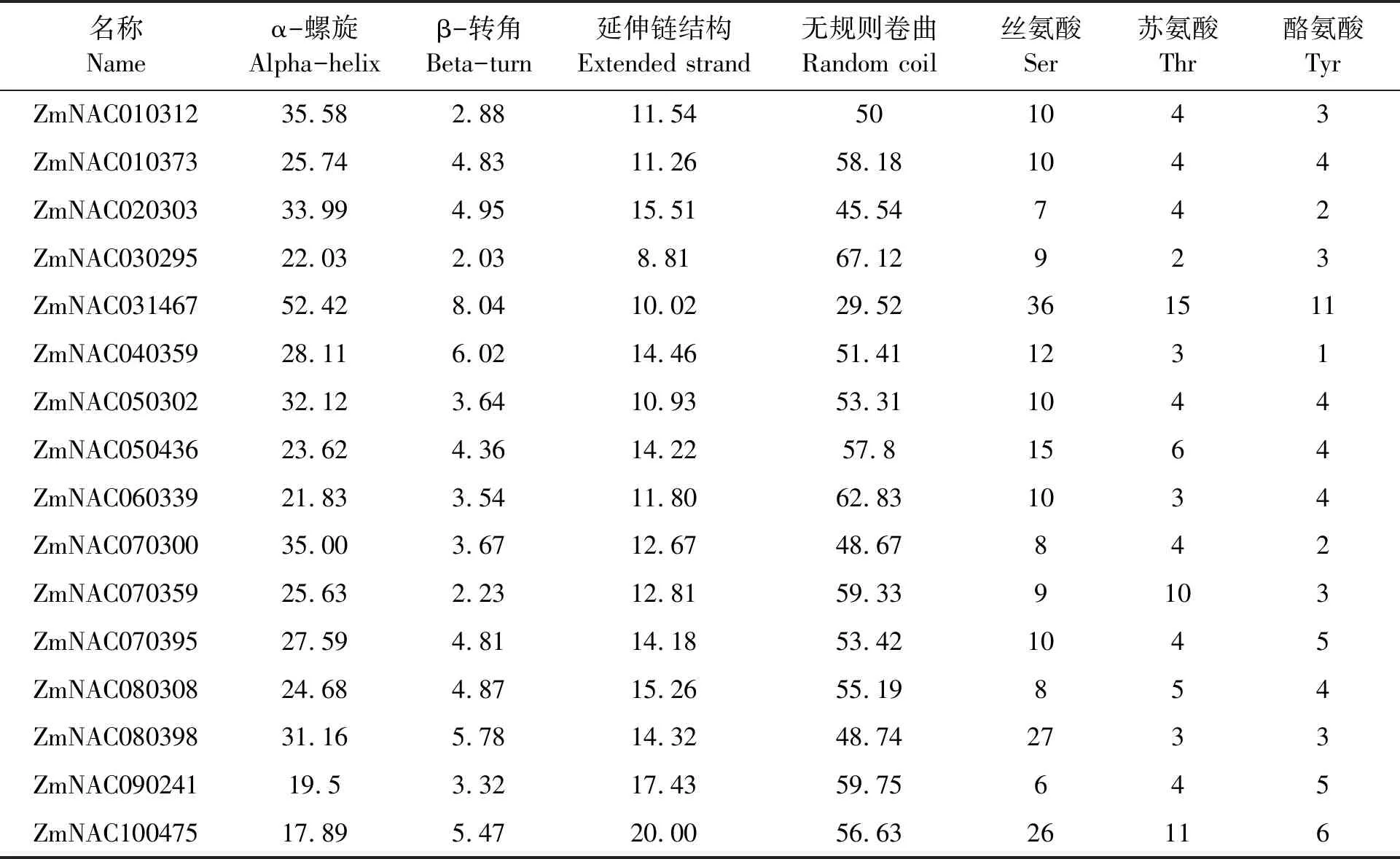
表2 SNAC家族成員的二級結構 4 種類型所占的比例及磷酸化位點預測Table 2 Proportion of 4 types of secondary structure of SNAC family members and prediction of phosphorylation sites

圖2 SNAC家族成員的二級結構Fig.2 Secondary structure of SNAC family members
2.7 玉米SNAC蛋白的保守結構域分析
利用MEME工具來識別玉米SNAC蛋白中10個不同的保守模體(圖3)。根據10個預測基序的分布,16個SNAC成員與系統發育分析的分類一致。NAC轉錄因子家族成員N端都具有高度保守的結構域,由大約150個高度保守的氨基酸殘基組成,可以形成螺旋-轉角-螺旋的三維結構,并能特異結合目標DNA和蛋白[34]。圖3中16個SNAC蛋白都具有基序2和基序3,而ZmNAC070359不具有基序1,在氨基酸序列分析中ZmNAC070359不具有子域A,但還是與剩余15個SNAC蛋白共享子域B和C。此外,盡管轉錄激活區相對不同,但在一些SNAC亞群中,轉錄激活區還是存在一些保守的子域。基序6在ZmNAC010312、ZmNAC050436和ZmNAC050302中都存在。同時,ZmNAC010312存在的基序5也與ZmNAC020303、ZmNAC050302、ZmNAC050436、ZmNAC070300共享。基序8也存在于多個基因的轉錄激活區域,如ZmNAC010373、ZmNAC031467、ZmNAC070395、ZmNAC080308、ZmNAC080398和ZmNAC100475。ZmNAC020303、ZmNAC031467、ZmNAC070300和ZmNAC080308都具有基序9。耐旱因子ZmNAC100475上的基序9也存在于ZmNAC031467、ZmNAC040359和ZmNAC080308中。
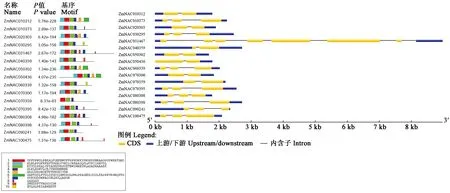
圖3 SNAC家族成員的基因結構分布及蛋白基序分布Fig.3 Distribution of gene structure and protein sequence of SNAC family members
2.8 玉米SNAC基因結構分析
對玉米SNAC基因的核苷酸結構分析可知,ZmNAC050436不存在上游調控區域,其余基因均存在上下游調控區域。在內含子方面,6個基因(ZmNAC010312、ZmNAC020303、ZmNAC050436、ZmNAC050302、ZmNAC070300、ZmNAC070359)由一個內含子構成,6個基因(ZmNAC010373、ZmNAC030295、ZmNAC060339、ZmNAC080308、ZmNAC080398、ZmNAC100475)由兩個內含子構成,3個基因(ZmNAC040359、ZmNAC070395、ZmNAC090241)由三個內含子組成,而ZmNAC031467由8個內含子構成,這可能與基因序列較長有關。玉米SNAC基因內含子較少,這可能與啟動下游基因表達有關,越少的內含子可以更快的啟動轉錄激活,從而加快相關基因的表達或抑制。ZmNAC050302、ZmNAC050436和ZmNAC070300三個基因結構相似度較高,分別與ZmNAC070300基因的相似度為50.88%和53.76%,同時ZmNAC050302和ZmNAC050436兩個基因相似度最高達93.58%。結合系統發育分析,很明顯同一亞群中最密切相關的成員具有相似的基因結構。
2.9 啟動子分析
SNAC基因的轉錄調控與啟動子區應激反應調控元件的存在有關,如DRE(dehydration-responsive elements)、ABRE(ABA-responsive elements)、茉莉酸反應元件、水楊酸反應元件、LTRE(low-temperature responsive elements)、MYB(myeloblastosis)和MYC(myelocytomatosis)結合位點[35]。通過生物信息學分析和預測,玉米SNAC基因家族啟動子區域均含有大量的逆境響應相關元件(表3),這些元件參與逆境脅迫應答(干旱)和脫落酸(ABA)誘導的表達調控,其中所有成員都含有DRE、MYB兩種調控元件;15個SNAC啟動子含有ABRE調控元件。ZmNAC030295、ZmNAC031467、ZmNAC090241的啟動子含有較多ABRE元件,分別含有10、14、12個,說明對脫落酸誘導的表達調控較為敏感。這三種轉錄因子與抗非生物逆境調控有關,另外還發現有MYC調控元件。此外,某些基因還具有獨特的順式作用元件,如ZmNAC030295含有多個參與光響應的順式作用元件(G-box、TCT-motif、TCCC-motif);ZmNAC050302含有赤霉素反應元件(GARE-motif);ZmNAC080398含有參LTRE順式作用元件 。以上結果表明,玉米SNAC家族基因通過這些元件的調控可能在玉米逆境響應中發揮著重要的作用。
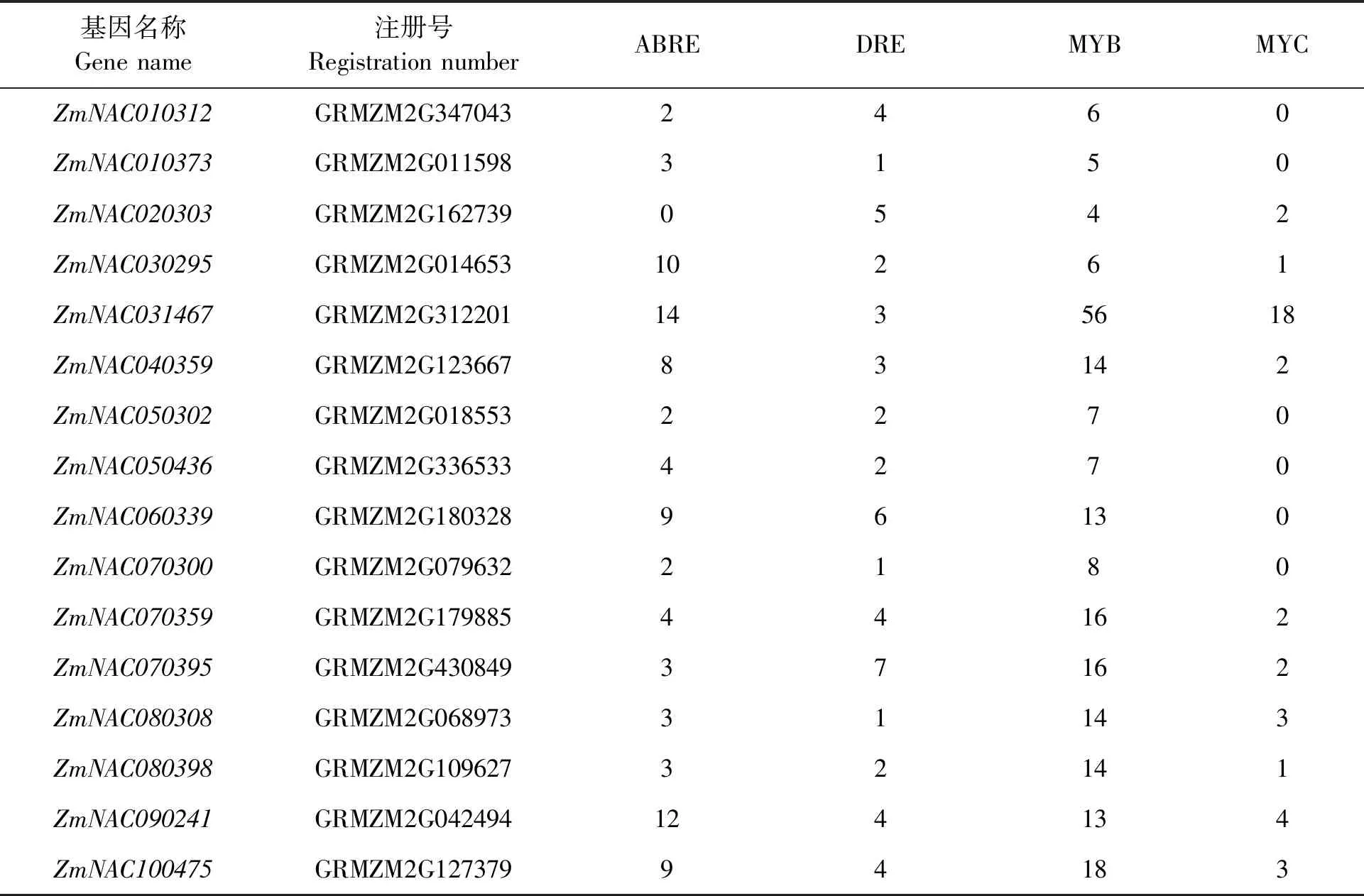
表3 玉米SNAC基因家族啟動子ABRE、DRE、MYB、MYC元件的數量分布Table 3 Distribution of ABRE,DRE,MYB,MYC components in promoters of SNAC gene family in maize
3 討論
玉米是我國三大糧食作物之一,是高效的經濟作物,對我國經濟發展有著十分重要的影響,也是調整我國農業生產結構的核心。非生物脅迫引發一系列植物反應,包括基因表達模式和細胞代謝的改變。轉錄因子調節介導多種非生物脅迫反應和生物過程的基因表達[36]。本研究對玉米SNAC家族進行了生物信息學分析,結果表明,玉米SNAC家族存在著大量的磷酸化位點,同時因不具有跨膜結構,推測均在細胞內表達和調控。此外,脂肪族氨基酸指數分布區域范圍較小,總體表明蛋白質間熱穩定性差異較小,而ZmNAC030295和ZmNAC080398的不穩定參數較高,因此推測該基因可能還具備其他功能,有助于提高玉米SNAC基因家族功能的多樣化。啟動子分析表明,玉米SNAC家族啟動子區域均含有大量的逆境響應相關元件,這些元件通過與相應的轉錄因子結合來調控下游基因的表達從而參與到植物抗逆響應。二級結構分析表明,玉米SNAC的轉錄調控區具有高度的內在混亂性,使得SNAC蛋白不能有一個單一穩定的結構,而這種靈活性使SNAC蛋白能與不同的靶蛋白相互作用,同時也使SNAC蛋白成為轉錄因子功能和結構系統分析的模式蛋白,造就了SNAC轉錄因子不同的功能作用。
根據在擬南芥、水稻、大豆中報道的幾個典型耐脅迫NAC基因,16個玉米SNAC家族蛋白共有4個保守基序,構成一個NAC結構域,在識別和結合特定DNA序列中起作用,而位于C末端的轉錄區域被認為是反式激活結構域[37]。據報道,SNAC1、GmSNAC49和CarNAC3所在非生物和生物脅迫耐受性中起著至關重要的作用。例如,水稻SNAC1基因導入小麥中的過表達株系與野生型相比,過表達的植株表現出對干旱和高鹽的耐受性顯著增強,正調控與非生物脅迫ABA信號轉導途徑關聯基因的表達[38]。大豆GmSNAC49的過表達導致干旱和ABA信號通路相關基因的上調,在耐旱性中發揮積極作用;鷹嘴豆CarNAC4在擬南芥中的過量表達提高了擬南芥對干旱和鹽脅迫的耐受性[39]。這些結果表明,進化關系密切的NAC基因在植物中可能具有相似的生物學功能。這些信息可用于探索植物,特別是主要作物玉米中潛在的脅迫相關SNAC基因。
系統進化分析表明,ZmNAC030295與TaNAC6基于同一亞群,同時有研究揭示,TaNAC6在提高小麥耐逆性和調控小麥生長發育方面的潛在作用[40]。同時ZmNAC050302與三個耐脅迫因子ZmNAC050436、SNAC1和ZmNAC010312在同一亞群,ZmNAC050302、ZmNAC050436和ZmNAC010312都僅有1個內含子。結構域分析表明,ZmNAC050302、ZmNAC050436、ZmNAC010312三個基因具有相同的基序,ZmNAC050302和ZmNAC010312共享基序6。這些結果預測ZmNAC050302在抗逆境方面有一定的正向調控作用。同時,ZmNAC080398與ZmNAC111(ZmNAC100475)的進化關系較近,具有相似的基因結構,在具有A~E亞域的基礎上還共享基序8,說明ZmNAC080398也可能在玉米脅迫環境中起著一定作用。同時,過量表達SNAC2和OsNAC6能提高轉基因水稻的抗旱和抗鹽能力[15-16],本研究中16個SNAC基因家族通過玉米抗旱自交系Tie 7922經過水分脅迫之后,各個基因在不同時間不同部位都發生顯著變化,比如,GRMZM2G430849_P01(ZmNAC050302)基因在脅迫后24 h表達量最高,而GRMZM2G162739_P01(ZmNAC080398)基因在1 h這個時間段表達最為明顯[43]。所以預測它們在非生物脅迫中參與了響應。
總之,本研究系統分析了玉米SNAC蛋白的系統發育關系和保守基序。在脅迫相關亞群中,一些預測的應激反應SNAC基因被篩選分離出來。生物信息學的數據加深了對玉米SNAC基因參與植物對非生物脅迫反應的理解,為進一步揭示玉米SNAC基因家族的耐逆境功能,抗逆分子育種和挖掘SNAC基因應用潛力提供參考。
