青藏高原金露梅灌叢草甸水分利用效率長期變化特征
2021-11-03 02:42:04王云英裴薇薇郭小偉杜巖功
草業科學 2021年9期
王云英,裴薇薇,辛 瑩,郭小偉,杜巖功
(1.中國科學院西北高原生物研究所,青海 西寧 810001;2.中國科學院大學,北京 100049)
金露梅(Potentilla fruticosa)灌叢草甸是青藏高原重要植被類型之一,面積約為0.106 × 106km2[1],僅次于高山嵩草草甸[2],廣泛分布于青藏高原東部海拔2 700~4 500 m 的山地陰坡、土壤濕度較高的平緩灘地以及地下水位較高的河谷階地[3],具有明顯的地帶分布性規律[4],是青藏高原主要的夏季牧場[5-6]。其生態功能對氣候變化和人類活動的響應比高寒草甸更加敏感[7]。
水分利用效率 (water use efficiency, WUE)是表征植物耐旱性[8]、生態系統碳水循環間相互耦合關系的重要指標[9-10],能夠反映生態系統對水資源變化的響應[11],對揭示大氣-葉片碳水循環過程中相互作用及植物生存適應對策有極其重要意義[12]。水分利用效率的計算及表達方式也因測定尺度存在差異。葉片尺度上,WUE 是植物凈光合速率與蒸騰速率的比值,主要由氣孔導度來控制[13];生態系統尺度上,WUE 是總初級生產力(gross primary productivity,GPP)和蒸散量(evapotranspiration, ET)的比值[14-15]。2003 ? 2010年,在8 個中國典型陸地生態系統中,當雄高寒草甸WUE 值最低,為0.18 g·kg?1[12]。2000 ?2014年黃土高原植被生態系統WUE呈顯著增加趨勢(P< 0.05), 增速為0.02g·(kg·a)?1[12]。 2000?2010年,黃土高原呈現以0.027 g·(m2·mm)?1的速度上升趨勢[12]。2000 ? 2017年新疆天山植被WUE 呈現以?0.014 1 g·(m2·mm)?1的速度下降趨勢[12]。西南高山亞高山區2000? 2014年植被WUE 以0.011 g·(m2·mm)?1的速率增加[16]。
WUE 受內部植被調控和外界環境的共同作用[12]。從植物自身調控而言,氣孔導度是影響植物氣體交換的主要影響因子[17]。氣孔導度主要通過影響植物蒸騰和通過氣孔進入植物的CO2含量來影響植物WUE[18]。在外界環境中,降水[19-22]是影響草地生態系統WUE 的主要影響因子,另外,相對濕度[23]、氣溫[12,24]、飽和水汽壓差[25]、光合有效輻射[26]等均是影響植被WUE 變化的關鍵氣候因子。年均氣溫和降水量是影響黑龍江西部農田WUE 的主要氣象因子[27]。西南高山亞高山區89.56%區域的WUE 與氣溫正相關,92.54%區域的WUE 與降水量負相關,針對不同生態系統植被類型,草地WUE 與氣溫相關性最高[16]。也有研究表明,高寒草甸WUE 與氣溫無顯著相關關系[28]。不同時間尺度影響WUE 的因子亦存在差異,光合有效輻射僅在短時間尺度內對WUE 影響效果顯著,而氣溫和相對濕度無論時間尺度長短都是影響WUE 的重要氣象因子[26]。
現階段,對于金露梅灌叢草甸的研究主要集中在植物群落特征[2,29-30]、土壤理化性質[31-34]、碳通量[3, 35-37]等方面。但對于金露梅灌叢草甸水分利用效率年際及季節變化特征的研究,以及氣象因子對水分利用效率的潛在控制過程方面的研究存在欠缺。因此,本研究基于2003 ? 2010年青海海北金露梅灌叢草甸GPP 及蒸散量ET 計算水分利用效率值,以期回答以下兩個科學問題:1) 連續8年間青海海北金露梅灌叢草甸WUE年份變化及月份變化特征;2)氣象因子對金露梅灌叢草甸WUE 的調控。旨在為區域生態建設、草甸的合理利用以及明晰金露梅灌叢草甸對氣象因子的適應調控機制提供理論基礎。
1 研究區概況
研究區位于中國科學院海北高寒草甸生態系統定位站干柴灘地區(37°39′56″ N,101°20′39″ E),地形為祁連山北支冷龍嶺東段南麓的洪積扇。受高原大陸性氣候影響,該區無明顯四季之分,一年只有冷暖兩季,冬季寒冷干燥,夏季濕潤多雨,年均氣溫為?1.7 ℃,年降水量為580 mm 左右(圖1),生長季降雨約占全年降水量的80%,且主要集中于植物生長季(5月 ? 9月)。土壤類型為高寒草甸土,高寒灌叢渦度相關觀測系統(37°40′ N,101°20′ E,3 400 m)位于海北站北部約8 km 的高寒金露梅灌叢生態系統內。金露梅灌叢渦度相關系統建于2002年8月,由10 m × 10 m 的圍欄保護,碳水熱通量的觀測高度為2.5 m。該區域群落結構比較簡單,一般分為灌木片層和草本植物片層。金露梅株高30~50 cm,生長較密集,群落總覆蓋度達70%~80%,以金露梅為建群種,伴生種有山生柳(Salix oritrepha)、高山繡線菊(Spiraca alpina)等,草本層植物生長發育較好,蓋度為50%~70%,優勢種包括線葉嵩草(Kobresiacapilliforlia)、喜馬拉雅嵩草(K.royleana)、青藏苔草(Carex moorcroftii)等。
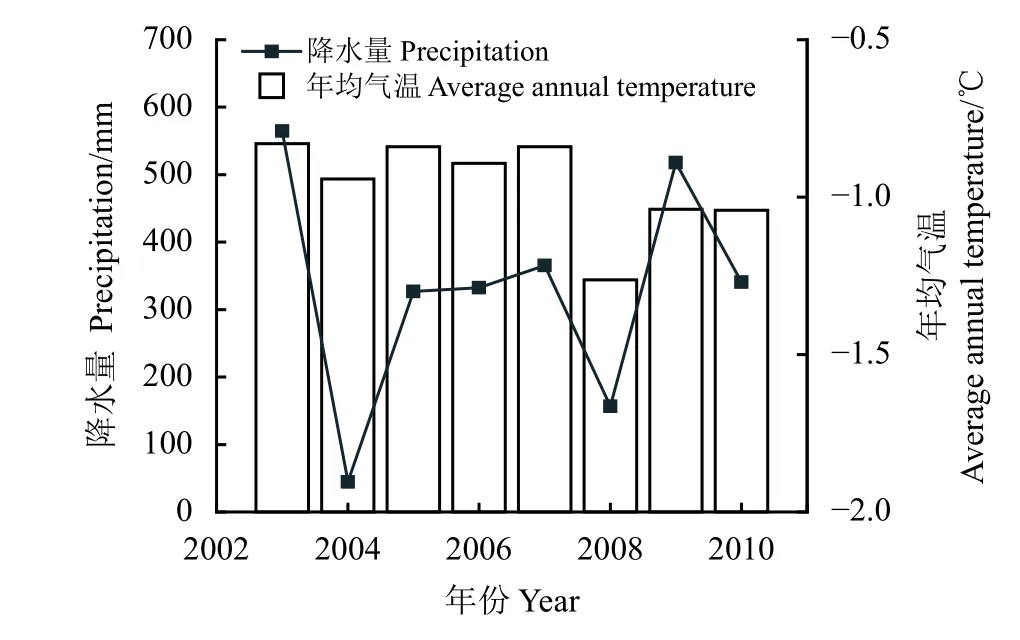
圖1年降水量和年均氣溫的變化Figure 1 Changes in annual precipitation and annual temperature between 2003 and 2010
2 研究方法
2.1 野外采樣及室內分析
金露梅灌叢草甸草地生物量于2003 ? 2010年植物生長季5月 ? 9月每月25日左右進行監測,地上生物量采用標準收獲法,樣方面積50 cm × 50 cm,重復5~8 次。地下生物量現存量測定采用根鉆法,按 0 ? 10、10 ? 20、20 ? 30 和30 ? 40 cm 分層采集,重復5 次,隨機設置于地上生物量調查樣方中,以水洗法進行根系的洗滌,65 ℃下烘干至恒重稱重。
土壤有機質(SOM):2003 – 2006年采用重鉻酸鉀氧化外加熱法測定;2006 – 2010年采用總有機碳分析儀(島津5000A)測定;有效氮(AN):氯化鉀浸提–蒸餾法測定;速效鉀[7]:乙酸銨浸提–火焰光度法測定;速效磷(AP):擴散吸收法測定;全氮[10]:半微量開氏法測定。土壤理化性質含量見表1。
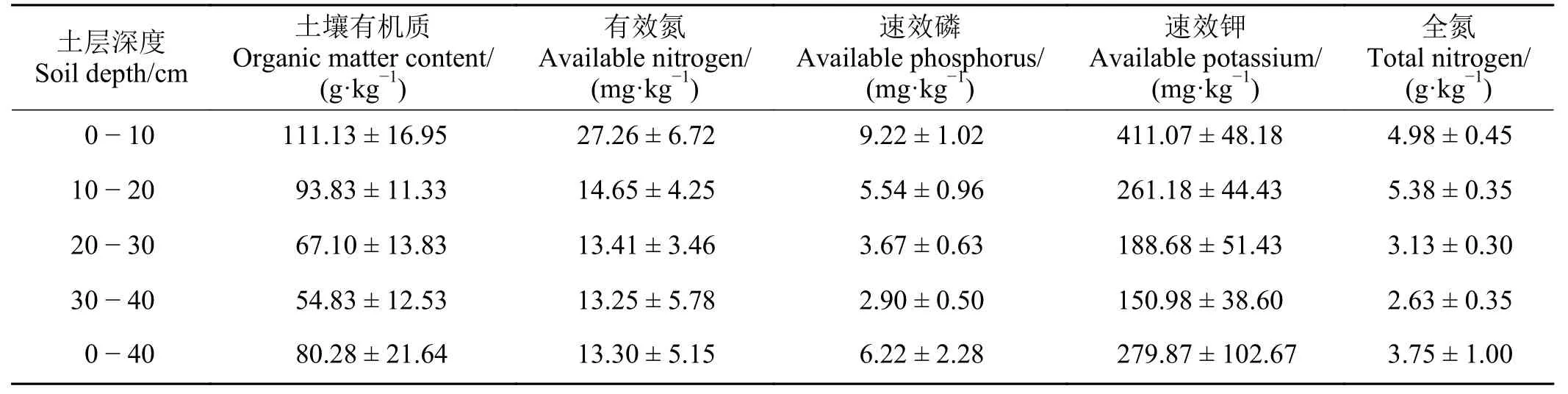
表1 金露梅灌叢草甸不同土層深度土壤養分特征Table 1 Nutrient profile of different soil layers found interacting with Potentilla fruticosa shrubs
2.2 氣象數據
本研究中氣象數據均來自于青海海北高寒草地生態系統國家野外科學觀測研究站氣象觀測站。
2.3 數據分析
2.3.1 蒸散量(ET)的計算
由渦度觀測中的潛熱通量LE (W·m?2)換算得到的[38]。計算公式如下:
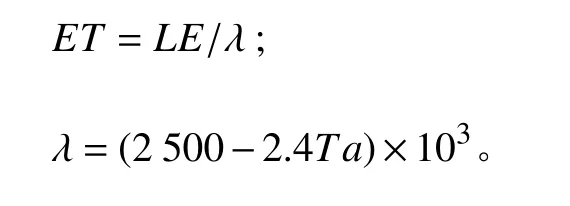
式中:λ為蒸發潛熱(J·kg?1);Ta為空氣溫度(℃)。
2.3.2 總初級生產力(GPP)的計算
站點通量數據來自國家生態系統觀測研究網絡科技資源服務系統(http://rs.cern.ac.cn/),數據時間尺度為2003 ? 2010年。通量數據包括碳凈交換量[8]和生態系統呼吸(Re)。通過獲取海北站日通量數據計算GPP,計算方法如下式:
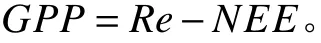
式中:GPP為植被總初級生產力(g·m?2);Re為白天生態系統呼吸量(g·m?2);NEE為白天生態系統CO2凈交換量(g·m?2)[39]。
2.3.3 水分利用效率的計算
本研究水分利用效率的計算采用以下公式:
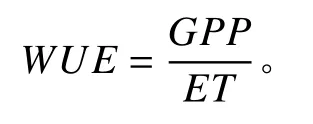
式中:WUE為水分利用效率[g·(m2·mm)?1];GPP和ET分別為生態系統總初級生產力(g·m?2)和實際蒸散量(mm)。
2.4 數據處理
數據整理采用Excel 2016,顯著性檢驗(單因素方差分析)、相對貢獻率的計算借助于增強回歸樹分析方法。增強回歸樹(BRT)是基于分類回歸樹算法(CART)的一種自學習方法,該方法通過隨機選擇和自學習方法產生多重回歸樹,能夠提高模型的穩定性和預測精度。在運算過程中多次隨機抽取一定量的數據,分析自變量對因變量的影響程度,剩余數據用來對擬合結果進行檢驗,最后對生成的多重回歸取均值并輸出。BRT 方法提高了計算結果的穩定性和精度,得出自變量對因變量的影響載荷,以及其他自變量取均值或不變的情況下,該自變量與因變量的相互關系[40]。在R 語言中(4.0.3 版本) 利用Dismo 軟件包實現。設置學習速率(learning rate)為0.001,樹的復雜度(tree complexity)為兩層,每次抽取50% 的數據進行分析,50% 用于訓練,并進行10 次交叉驗證。相關性分析的計算采用R 語言(4.0.3版本)軟件處理。運用origin 2019 進行圖件的繪制。
3 結果與分析
3.1 金露梅灌叢草甸生態系統水分利用效率年際變化特征
年際尺度上,金露梅灌叢草甸2003 – 2010年GPP、WUE 呈增加趨勢,且均達到極顯著性水平(P<0.001) (圖2),E呈微弱增加趨勢,8年間GPP 與ET、WUE均值分別 為326.66g·m?2、810.15mm 和756.03 g·(m2·mm)?1。2010年相較于2003年GPP 上升了1.14 倍;ET 下降了3.44%;WUE 上升了2.49 倍。GPP、ET 和WUE 分別以每年98.55 g·m?2、40.15 mm和138.70 g ·(m2·mm)?1增加趨勢增加(表2)。
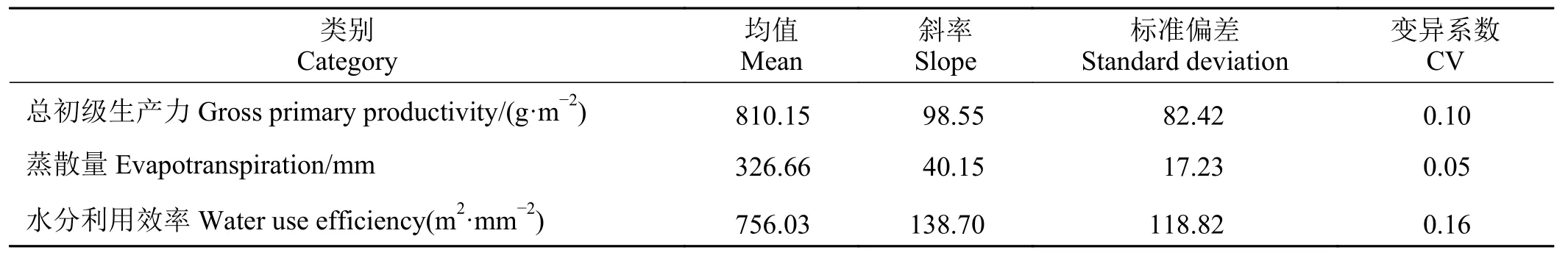
表2 高寒草甸總初級生產力、蒸散量與水分利用效率年變化統計特征值Table 2 Characteristic values of annual changes in gross primary productivity,evapotranspiration and water use efficiency in alpine meadows

圖2 總初級生產力、蒸散量和水分利用效率年際變化Figure 2 Interannual variation of gross primary productivity, evapotranspiration, and water use efficiency
3.2 金露梅灌叢草甸生態系統水分利用效率季節尺度對比分析
季節尺度上,GPP、ET 和WUE 均呈倒V 型變化(圖3)。ET 在生長季盛期7月或8月份達到最大值,7月 和8月份均值分別為57.06 和57.18mm;1月份降到最小值,均值為4.99 mm。相較于其余月份,6月、7月和8月3 個月份蒸散量的變化相對較小,1月 ? 7月或8月呈快速上升趨勢,之后呈快速下降趨勢。GPP 含量在7月份達到最大值,均值為221.77 g·m?2,1月 ? 7月呈上升趨勢,且從生長季初期5月份開始呈快速上升,7月? 12月呈下降趨勢。整體而言,WUE 在1月?7月呈上升趨勢,5月 ?7月呈快速上升,7月份達到最大值,7月 ? 12月呈下降趨勢,7月份WUE 均值為3.76 g·(m2·mm)?1,在凍土凍結時期12月WUE 最低,全年和生長季WUE差異顯著性對比結果發現生長季WUE 顯著高于全年WUE (P< 0.05)。
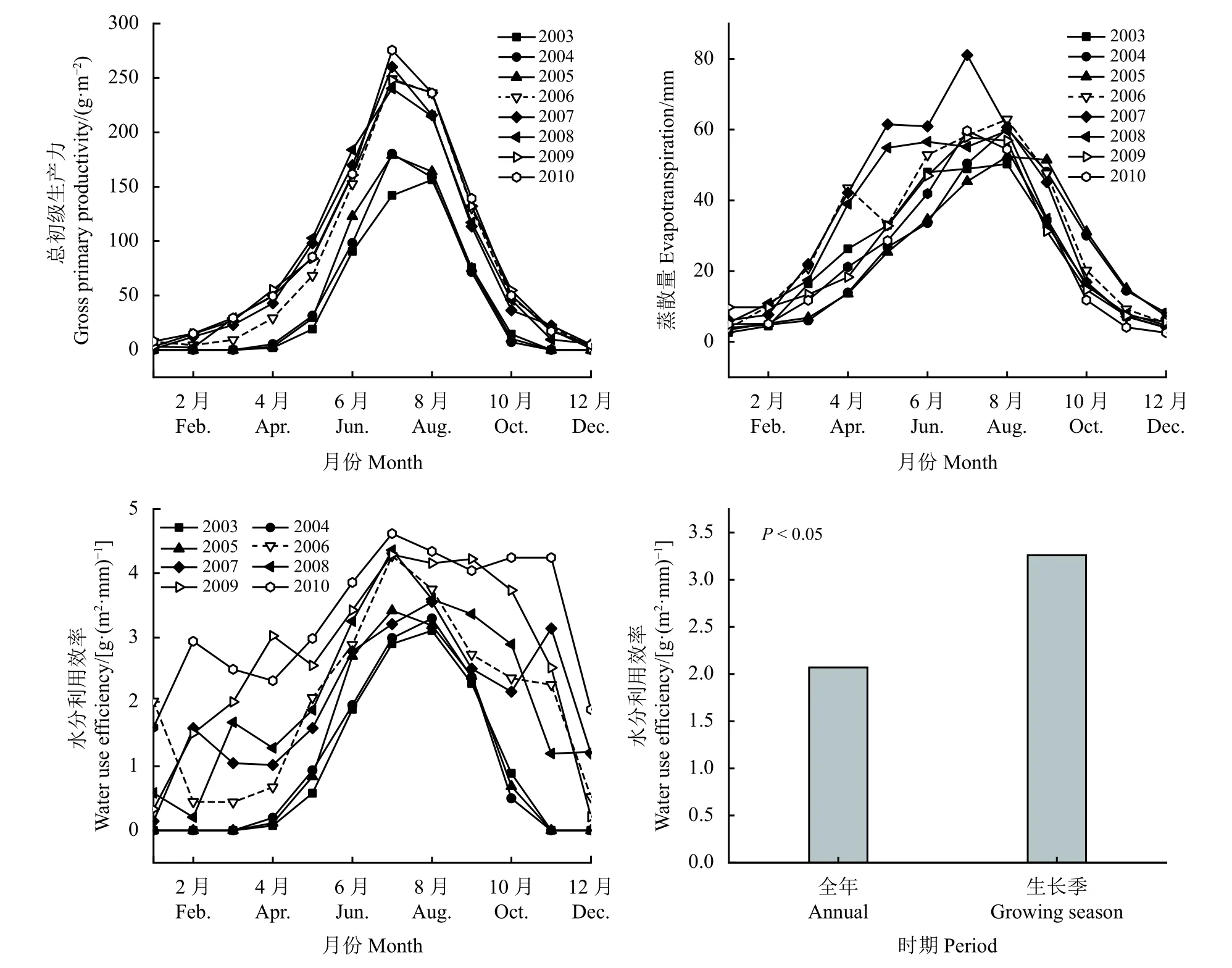
圖3 總初級生產力、蒸散量和水分利用效率生長季變化Figure 3 Seasonal changes in gross primary productivity, evapotranspiration and water use efficiency in alpine meadows
3.3 金露梅灌叢草甸水分利用效率對氣象因子的響應
增強回歸樹的模擬結果顯示,在全年尺度上,年均氣溫、相對濕度和凈輻射3 種氣象因子對WUE 的相對貢獻率最大,三者能解釋73.50%的全年水分利用效率變化。生長季尺度上,光合有效輻射、空氣溫度和相對濕度3 種氣象因子相對貢獻率最大,三者能解釋74.17%的生長季水分利用效率變化。年均氣溫和光合有效輻射分別是全年尺度和生長季尺度上對金露梅灌叢草甸WUE 貢獻率最大的環境因子,值分別為43.90%和30.79%。其與環境因子相對貢獻率的大小順序為飽和水汽壓差 > 風速 >凈輻射 > 降水量。其余環境因子相對貢獻率的變化順序為光合有效輻射 > 飽和水汽壓差 > 風速 > 降水量(圖4)。相關性分析結果顯示,在全年尺度上,與水分利用效率相關性大小順序為年均氣溫 > 飽和水汽壓差 > 相對濕度 > 降水量 > 風速 > 凈輻射 >光合有效輻射,除光合有效輻射外,其余因子均達到極顯著性水平(P< 0.01) (表3)。生長季尺度上相關性大小順序為光合有效輻射 > 相對濕度 > 凈輻射 >年均氣溫 > 風速 > 降水量 > 飽和水汽壓差,除飽和水氣壓以外,其余因子均達到極顯著水平(P<0.01) (表4)。
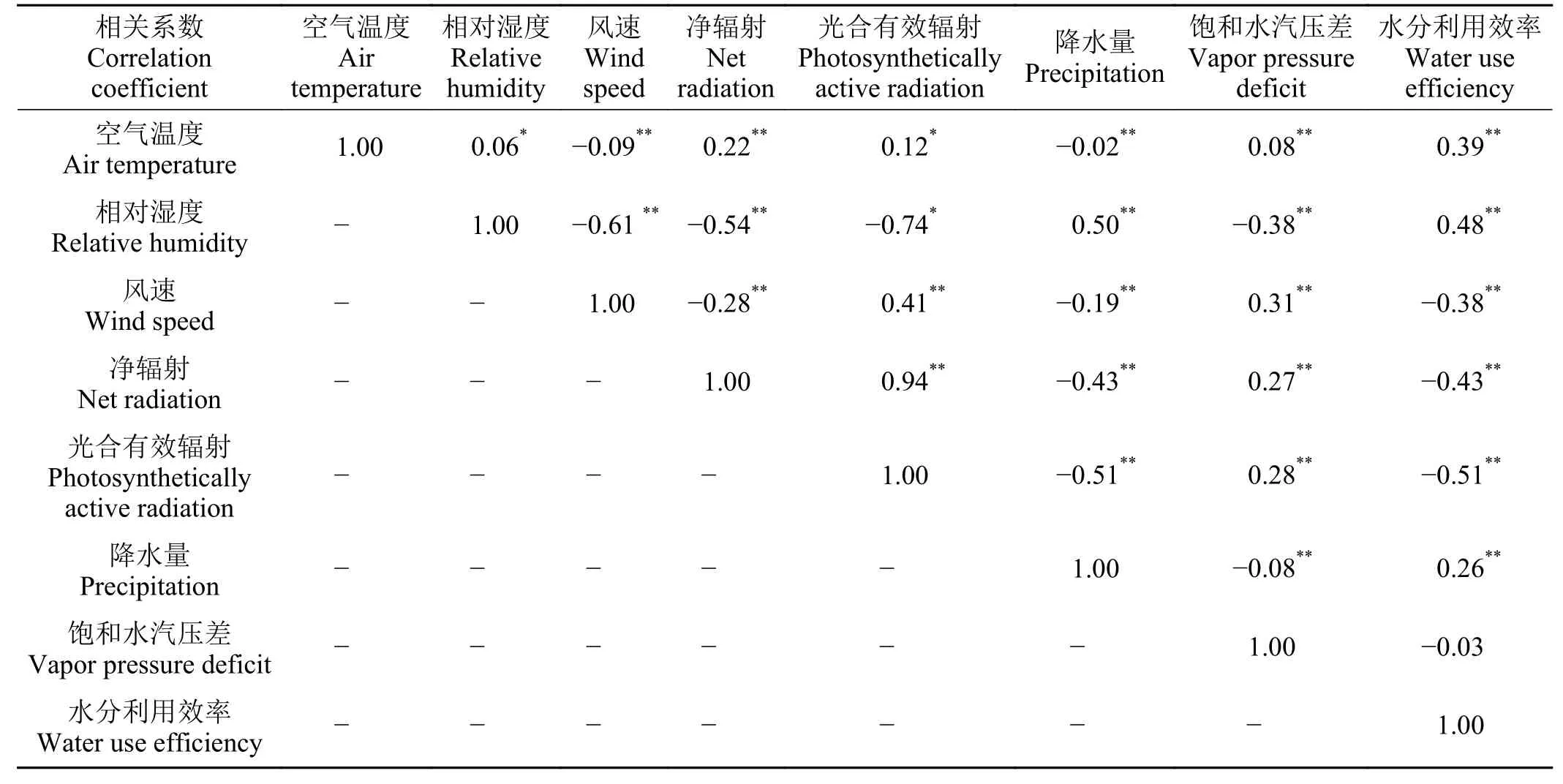
表4 高寒草甸生長季水分利用效率與氣象因子之間的關系Table 4 The relationship between water use efficiency and meteorological factors as a function of the growing season of alpine meadows

圖4 氣象因子對高寒草甸水分利用效率的相對貢獻率Figure 4 Relative contribution of various meteorologicalfactors on the water use efficiency of alpine meadows
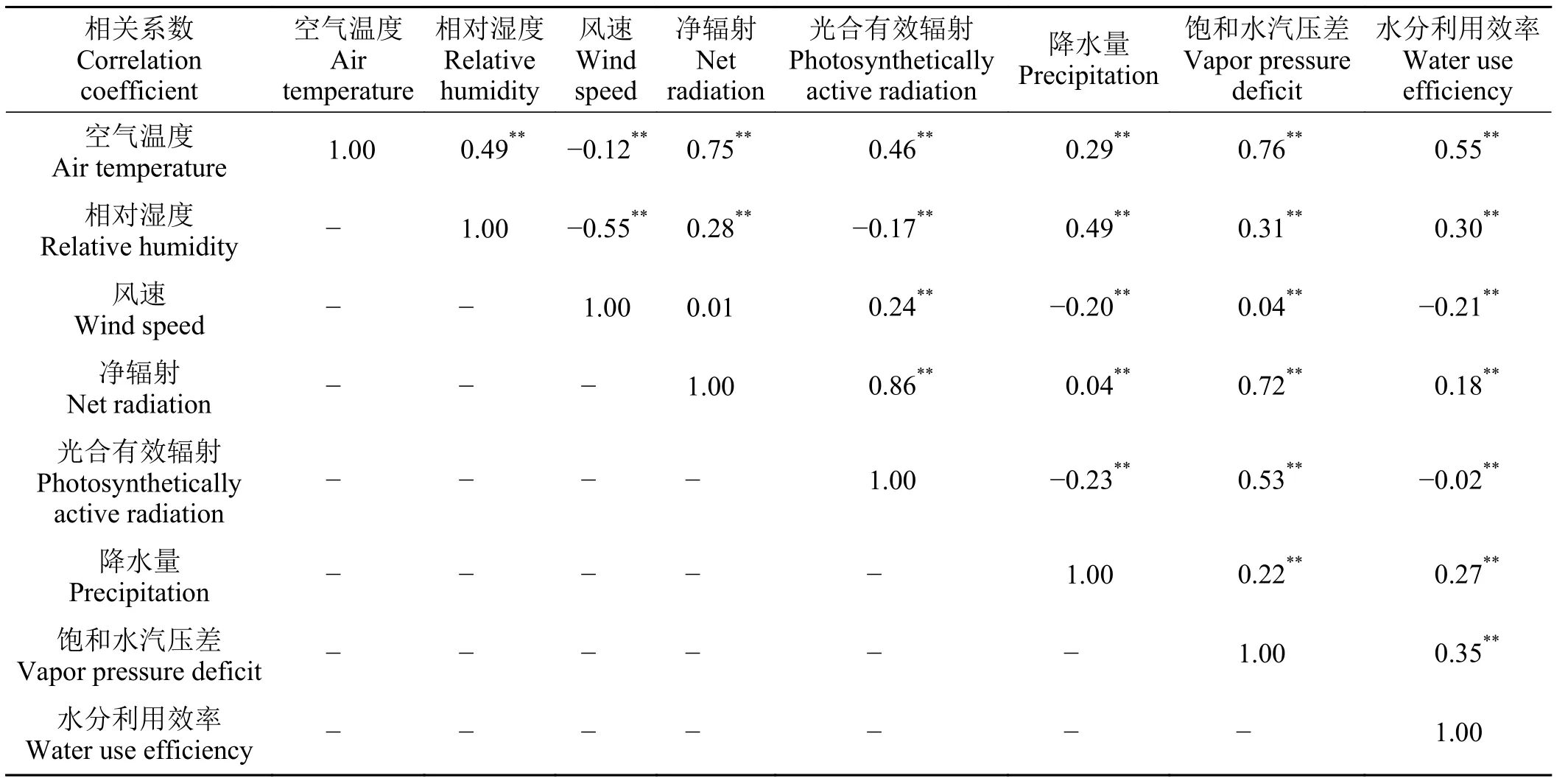
表3 高寒草甸水分利用效率與氣象因子之間的關系Table 3 The relationship between water use efficiency and various meteorological factors in alpine meadows
4 討論
由于植被WUE 主要受到GPP 和ET 兩種因素影響,故本研究亦分析了青海海北金露梅灌叢草甸GPP 和ET 的變化,結果發現2004 – 2009年GPP 和ET 呈上升趨勢,WUE 呈微弱下降趨勢。2000 – 2014年三江平原植被ET、GPP 和WUE 的變化與本研究結果一致[19]。新疆天山地區2000 – 2017年植被WUE亦呈減小趨勢,究其原因是因為這17年間ET 隨年份增加的趨勢高于GPP,且通過對水田和旱田的研究發現此地區水分利用效率變化主要受到人類不合利用等非氣候因素影響,故WUE 呈減小趨勢[12]。基于PT-JPL 模型研究退耕還林后的黃土高原于2001 – 2015年WUE 呈增加趨勢是因為退耕還林后植被生產力顯著增加[11]。2008 – 2016年青海海北金露梅灌叢草甸總初級生產力呈上升趨勢[2]。本研究中2004 – 2009年水分利用效率呈下降趨勢亦是因為連續6年間蒸散量的上升趨勢高于總初級生產力,故水分利用效率呈下降趨勢。
就季節變化而言,5月 ? 7月GPP、ET 和WUE均呈快速上升趨勢,WUE 在7月份達到最大值,7月?12月呈下降趨勢。2000?2018年中亞五國WUE 在夏季6月 ? 8月達到最大值[41]。這可能是因為在植物生長季盛期,GPP 的增加趨勢高于ET。2000–2017年新疆天山植被WUE 在7月份達到最大值,為1.34 g·(m2·mm)?1,究其原因是因為受到氣候條件和植被類型的影響[12]。新疆草地全年WUE平均值為0.56 g·kg?1,且在夏季達到最大值。這是因為新疆屬于雨熱同期的氣候, 夏季水熱組合條件相對較好,利于牧草生長[42]。
根據本研究中WUE 表達式可知,影響GPP 和ET 兩個指標的影響因子均會影響WUE。飽和水汽壓差是溫度的函數,在一定程度上也影響著土壤蒸發[26,43],進而影響WUE。本研究中,溫度是影響金露梅灌叢草甸WUE 的主要因子,且呈正相關關系關系。有研究表明,當溫度低于植被光合作用的最適宜溫度時,氣溫的上升將提升植被光合速率,從而產生更多的干物質量,進而提高植物WUE[44]。溫度對WUE 的影響因區域、環境等既有正效應影響又存在負效應影響。南方亞熱帶地區水分利用效率與氣溫呈正相關關系[45]。長江源區WUE 因常年低溫和氣溫回暖與水分利用效率之間呈正相關關系[46]。空氣相對濕度是興安落葉松林8月份水分利用效率的驅動因子[47]。大興、大滿、密云和張掖4 個站在0.5 h 尺度上相對濕度與WUE 之間呈現正相關關系[26]。藏北高寒草甸[48]和遼寧東部陸域生態涵養區[49]WUE 與相對濕度之間呈現負相關關系,與本研究結果相反。有研究表明,年均氣溫、相對濕度、飽和水氣壓和光合有效輻射與WUE 之間正負相關關系由該站點的WUE 是由GPP 主導還是由ET 主導所決定的[26]。風速與WUE 之間呈負相關關系,其主要原因是在植被覆蓋度較低的環境中風速主要影響蒸散量來影響WUE[50]。凈輻射主要通過影響草地蒸散發來影響WUE,水分蒸散作用隨著太陽輻射增加導致的空氣溫度升高而增大,故凈輻射與WUE 之間存在負相關[51]。光合有效輻射對植物固碳能力和水分消耗存在主要影響,當GPP 在強光下達到飽和狀態時,ET 會隨著光合有效輻射的增大而增大,故WUE會隨PAR 的增大而減小[26]。科爾沁草甸WUE 與飽和水汽壓差和空氣溫度均呈增加二次曲線形式,在生長季初期,GPP 含量隨著飽和水汽壓的增大呈增加趨勢,ET 與飽和水汽壓之間呈微弱正相關關系[25]。三江平原植被WUE 與降水量和相對濕度呈正相關關系[19],這與本研究結果一致,究其原因可能是植被三江平原區海拔越高,植被生長狀況越好,蒸散量較低,故植物WUE 呈增加趨勢[52]。年降水量是驅動年際間生態系統WUE 差異的主要因子[53],是影響伊犁河谷[20]和2005?2014年黑河流域[54]、潘陽湖流域[22]WUE 的主導因素,伊犁河谷降水量與WUE 呈正相關關系,這是因為降水減少會直接降低蒸散量,進而導致WUE 增大。黑河流域由于植被類型的不同,與降水量的關系也存在一定差異。與降水量呈正相關關系的很多區域海拔,ET 較低,植被的固碳能力較強,故WUE 增大[19]。本研究結果顯示,金露梅高寒草甸WUE 與相對濕度與年均氣溫呈極顯著正相關。這可能是因為西北地區草地WUE 與溫度呈正相關關系,且受溫度影響大[24]。而相對濕度主要通過影響GPP 來影響金露梅灌叢草甸WUE。
5 結論
總體而言,年際尺度上,GPP 和WUE 呈現極顯著上升趨勢(P< 0.01),ET 呈現微弱上升趨勢。季節尺度上,GPP、ET 和WUE 均呈倒V 形變化,并于7月份達到最大值,12月份呈現最小值。生長季的WUE 顯著高于全年WUE (P< 0.05)。
無論是年際尺度還是生長季尺度,金露梅灌叢草甸的水分利用效率主要受控于年均氣溫和相對濕度兩種氣象因子,且均呈正相關關系。全年尺度上還有凈輻射,生長季尺度上還有光合有效輻射。金露梅灌叢草甸WUE 會隨著凈輻射和光合有效輻射兩種因子的增大而減小。本研究連續8年在短時間尺度上對金露梅灌叢WUE 變化特征和適應機制進行了研究,但對于長時間尺度上WUE 變化特征及其調控機制還有待進一步研究。
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
第一財經(2021年6期)2021-06-10 13:19:08
當代陜西(2021年2期)2021-03-29 07:41:24
Coco薇(2017年9期)2017-09-07 21:23:49
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
紡織服裝流行趨勢展望(2016年2期)2016-05-04 03:47:15
通信電源技術(2016年3期)2016-03-26 07:13:38
中國衛生(2015年7期)2015-11-08 11:09:38
汽車科技(2015年1期)2015-02-28 12:14:44
