日本結縷草ZjNAC3基因在鹽脅迫中的功能
2021-11-03 02:42:10姜紅巖范希峰溫海峰滕文軍尹淑霞
草業科學 2021年9期
姜紅巖,范希峰,溫海峰,韓 朝,滕文軍,滕 珂,尹淑霞
(1.北京市農林科學院,北京 100097;2.北京林業大學草業與草原學院,北京 100083)
土壤鹽堿化是影響植物生長的主要非生物脅迫,嚴重影響作物產量[1]。鹽脅迫會使植物產生離子毒害作用,影響植物吸收水分,破壞生理機制導致植物枯萎死亡[1-2]。草坪草生長容易受到外界環境的影響,坪觀質量在園林綠化、運動場應用中是一項重要的衡量標準。鹽脅迫影響草坪草種子的萌發,草坪草的生長發育、營養物質積累及抗氧化酶的活性,從而影響草坪的使用價值[3-4]。
轉錄因子在植物的生長發育、抵抗環境脅迫中發揮著重要作用[5]。已有研究證明,轉錄因子能夠通過調控上下游基因影響植物對鹽脅迫的耐受性,其中包括AP2/ERF、NAC、WRKY、MYC 等轉錄因子家族[5]。NAC 轉錄因子在模式植物中被率先得以研究,發現其在鹽脅迫響應中發揮著重要作用[6]。NAC 轉錄因子家族成員在調控鹽脅迫中具有不同的功能,有些NAC 轉錄因子在植物的抗鹽性方面起負調控作用,而另一些成員發揮著正調控的作用。如NAC 轉錄因子NTL8能夠通過赤霉素(gibberellin acid,GA)信號傳導途徑負調節種子的發芽[7];過表達ANAC092基因能降低鹽脅迫下種子的發芽能力[8]。而PvNAC1[9]、SNAC1[10]、ONAC045[11]、OsNAC2[12]、OsNAC6[13]等轉錄因子能夠增強植物的耐鹽性。
結縷草(Zoysia japonica)是我國一種重要的鄉土草坪草,具有抗逆性強、養護成本低等優點[14]。在結縷草中鮮有NAC 轉錄因子對植物的耐鹽性影響的報道,之前研究表明日本結縷草中的ZjNAC2基因能夠受到300 mmol·L?1NaCl 的誘導表達[15],而NAC 轉錄因子在結縷草適應鹽脅迫中的作用尚不清楚。本研究通過獲得轉ZjNAC3基因的酵母菌株和擬南芥(Arabidopsis thaliana)植株,并對其進行鹽脅迫處理,旨在探究日本結縷草NAC 轉錄因子ZjNAC3 在鹽脅迫下的調控作用,以期為深入研究NAC 轉錄因子調控日本結縷草耐鹽性及其機理奠定前期工作基礎。
1 材料與方法
1.1 材料
農桿菌GV3101、擬南芥種子、pYES2 載體、YPH500 酵母菌株均為北京市農林科學院草業花卉與景觀生態研究所實驗室保存;Ura 培養基購自北京泛基諾公司;氯化鈉等分析純試劑購自北京科百奧公司;RNA 試劑盒購自OMEGA 公司;SYBR Premix購自TaKaRa 公司。本試驗所用擬南芥植株在江南人工氣候培養箱中培養,培養條件為24 ℃/22 ℃ (日/夜),16 h 光照,濕度65%。
1.2 方法
1.2.1 酵母轉化及轉基因酵母的鹽脅迫處理
以pYES2-ZjNAC3-F/R 為 引 物(表1),pMD19-ZjNAC3 克隆載體為模板擴增,將擴增產物連接到pYES2 載體上構建重組質粒pYES2-ZjNAC3。按照Clontech 的說明書(貨號: 630439) 轉化酵母,分別將構建的pYES2-ZjNAC3 和pYES2 空載質粒轉化YPH500 酵母菌株。挑取SD/-Ura 培養基平板上生長的單菌落于Ura 液體培養基中培養至OD600值為0.2,以pYES2-F/R (表1)為引物進行菌落PCR 鑒定后,按照 1 ? 10、1 ? 100、1 ? 1 000、1 ? 10 000 的比例進行稀釋。將稀釋后的菌液接種到含有0、200、300 mmol·L?1NaCl 的Ura 固體培養基上培養2~3 d,觀察酵母菌落的生長情況。
1.2.2 擬南芥轉基因
以3302Y3-ZjNAC3-F/R 為引物(表1),以pMD19-ZjNAC3 克隆載體為模板進行PCR 擴增,將擴增產物連接到3302Y3 載體[15]上,構建3302Y3-ZjNAC3。取5 μL 構建好的質粒于50 μL 農桿菌感受態中,冰浴30 min,液氮中速凍1 min,37 ℃下水浴5 min,冰浴5 min,之后28 ℃暗培養4 h 后涂布于培養基上培養,2~3 d 后挑取單克隆進行PCR 鑒定。對鑒定正確的菌液用50%甘油進行保菌,于?80 ℃冰箱保存以備用。然后用獲得的轉化目的質粒的GV3101農桿菌菌液浸染擬南芥花序,選擇生長3~4 周大小的擬南芥植株,將其花序浸到農桿菌菌液中,浸染時間不超過30 s,暗培養12 h 后恢復正常的光照條件。待植株生長成熟后收獲的種子備用。

表1 引物列表Table 1 List of primers used in this study
1.2.3 轉基因植株的鑒定
將收獲的T0代擬南芥種子放于冰箱中4 ℃下春化2~3 d,播種于草炭 ? 蛭石 ? 珍珠巖配比為3 ? 1 ? 1的基質中。播種后1~2 周,對幼苗噴施60 mg·L?1的草銨膦溶液一次進行初步篩選。存活下來的擬南芥植株生長至3~4 周后,用CTAB 法提取葉片DNA,并以3302Y3-F/R (表1) 為引物進行轉基因植株鑒定。擴增產物經凝膠電泳檢測后,選擇條帶大小符合的擬南芥植株繼續收獲種子,直至篩選至T3代后用于后續試驗。
1.2.4 對轉基因擬南芥植株的鹽脅迫處理
選用穩定遺傳的T3代轉基因擬南芥種子,先用1%次氯酸鈉溶液消毒5 min,70%酒精消毒30 s,無菌水清洗4~5 次,然后播種在MS 培養基上,生長3 周后轉移到基質中生長。待植株生長1 個月時,提取整株轉基因擬南芥的RNA,以ZjNAC3-F/R 為引物(表1)對ZjNAC3進行表達量分析。選擇ZjNAC3基因表達量相對較高的株系和野生型(WT)進行鹽脅迫處理,每個株系設置4 個重復。每盆第1 天澆灌50 mL 50 mmol·L?1的NaCl,第2天澆灌50 mL 100 mmol·L?1NaCl,第3 天澆灌50 mL 150 mmol·L?1NaCl,對照澆灌相同量的蒸餾水,處理7 d后拍照并剪取葉片測定相關指標。
采用蒽酮顯色法[16]、茚三酮比色法[16]、浸提法[17]、硫代巴比妥酸(TBA)比色法[18]分別測定可溶性糖、脯氨酸、葉綠素、丙二醛含量,同時參照陳愛葵等[19]的方法測定細胞膜透性。利用RNA 試劑盒提取RNA,反轉錄為cDNA。通過qRT-PCR 技術,以AtUBQ 為內參,參照Teng 等[14]設 計AtNHX1、AtAPX1、AtPOD 和AtUBQ 引 物(表1),采 用2?ΔΔCT法測定AtNHX1、AtAPX1、AtPOD基因的表達情況。
2 結果與分析
2.1 ZjNAC3 基因對鹽脅迫下酵母菌株生長的影響
本研究成功構建了載體pYES2-ZjNAC3 (圖1),同時將pYES2-ZjNAC3 載體轉到酵母YPH500 菌株中進行鹽敏感性測定(圖2)。
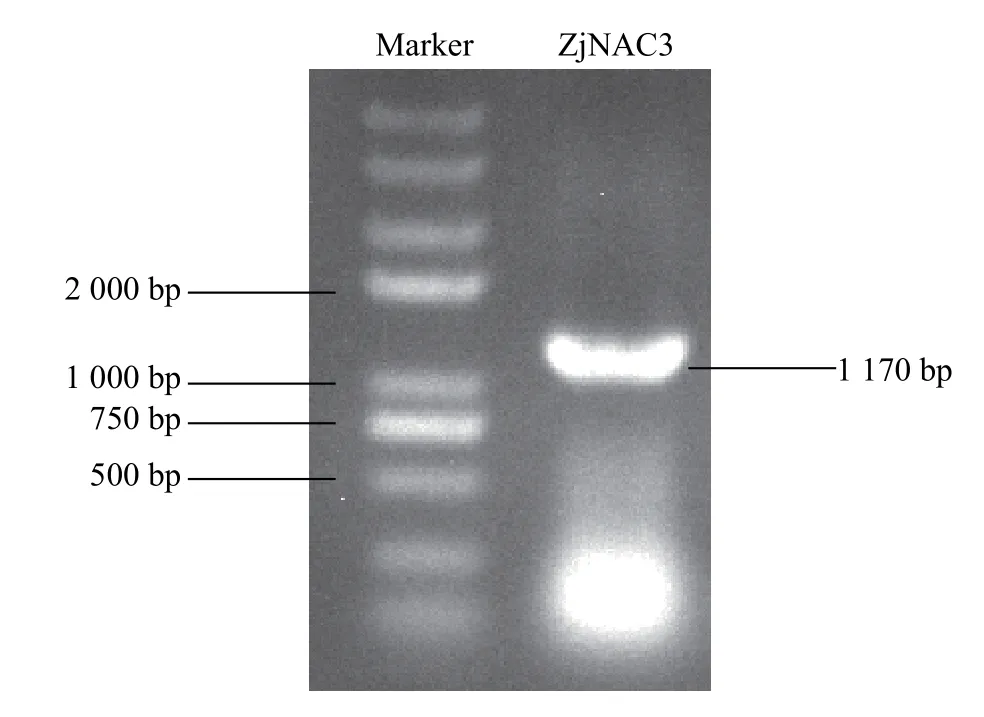
圖1 pYES2-ZjNAC3 載體構建的PCR 檢測Figure 1 PCR verification of the constructed pYES2-ZjNAC3 vector

圖2 ZjNAC3 對轉基因酵母的影響Figure 2 Effect of ZjNAC3 on the growth of transgenic yeast cells under salt stress
結果表明,在Ura 固體培養基上轉空載與重組質粒pYES2-ZjNAC3 的酵母菌株在相同的培養時間內長勢基本一致。而在鹽脅迫下,含有重組質粒的酵母菌株在濃度稀釋104倍后其長勢明顯弱于對照;同時,隨著Ura 固體培養基上鹽濃度的增加,酵母菌株的長勢呈逐漸減弱的趨勢。
2.2 轉基因植株的獲取
噴施草銨膦后,大部分T1代植株幼苗枯萎死亡,只有少部分幼苗存活下來(圖3)。對存活的植株提取DNA,通過PCR 檢測進一步篩選抗性植株,將含有目的基因條帶的擬南芥株系繼續篩選培養,獲得穩定遺傳的T3代植株(圖4)。

圖3 草銨膦篩選后的抗性植株Figure 3 Screening of transgenic plants using glufosinate

圖4 轉基因擬南芥T3 代植株Figure 4 Transgenic Arabidopsis thaliana (T3 generation)
2.3 鹽脅迫下過表達ZjNAC3 基因擬南芥的形態變化
對過表達ZjNAC3基因的擬南芥株系中的ZjNAC3基因進行表達分析,發現在這些轉基因株系中均有ZjNAC3基因的表達,且表達水平不一致,本研究中選用了基因表達量相對較高的40#和57#兩個株系進行鹽脅迫處理(圖5)。

圖5 轉基因株系T3 代中ZjNAC3 基因的表達水平Figure 5 Expression levels of the ZjNAC3 gene in the transgenic T3 Arabidopsis lines
為了進一步探究ZjNAC3基因是否參與鹽脅迫響應,對過表達擬南芥進行鹽敏感性測定(圖6),可以看出,鹽處理第7 天對照處理中WT 與轉基因株系的生長狀態基本一致,處理組中WT 的生長狀態明顯好于轉基因株系,這體現在WT 葉片顏色更深,覆蓋度更高。相比之下,轉基因株系中葉片明顯黃化,尤其是40#株系。

圖6 150 mmol·L?1 NaCl 處理后轉基因擬南芥株系的表型Figure 6 Phenotype of transgenic Arabidopsis thaliana lines grown in the presence of 150 mmol·L?1 NaCl
2.4 鹽脅迫下過表達ZjNAC3 基因擬南芥的生理響應
鹽脅迫處理后轉基因擬南芥與對照相比,生理指標發生了明顯變化(圖7)。在鹽處理7 d 后,兩個轉基因株系中的脯氨酸含量、丙二醛含量、電解質滲透率顯著高于WT (P< 0.05),表明在鹽脅迫下轉基因株系受到的傷害程度更高;而可溶性糖含量、葉綠素含量顯著低于WT (P< 0.05),說明轉基因株系在鹽脅迫下的葉綠素積累和滲透調節的能力受到了明顯抑制。
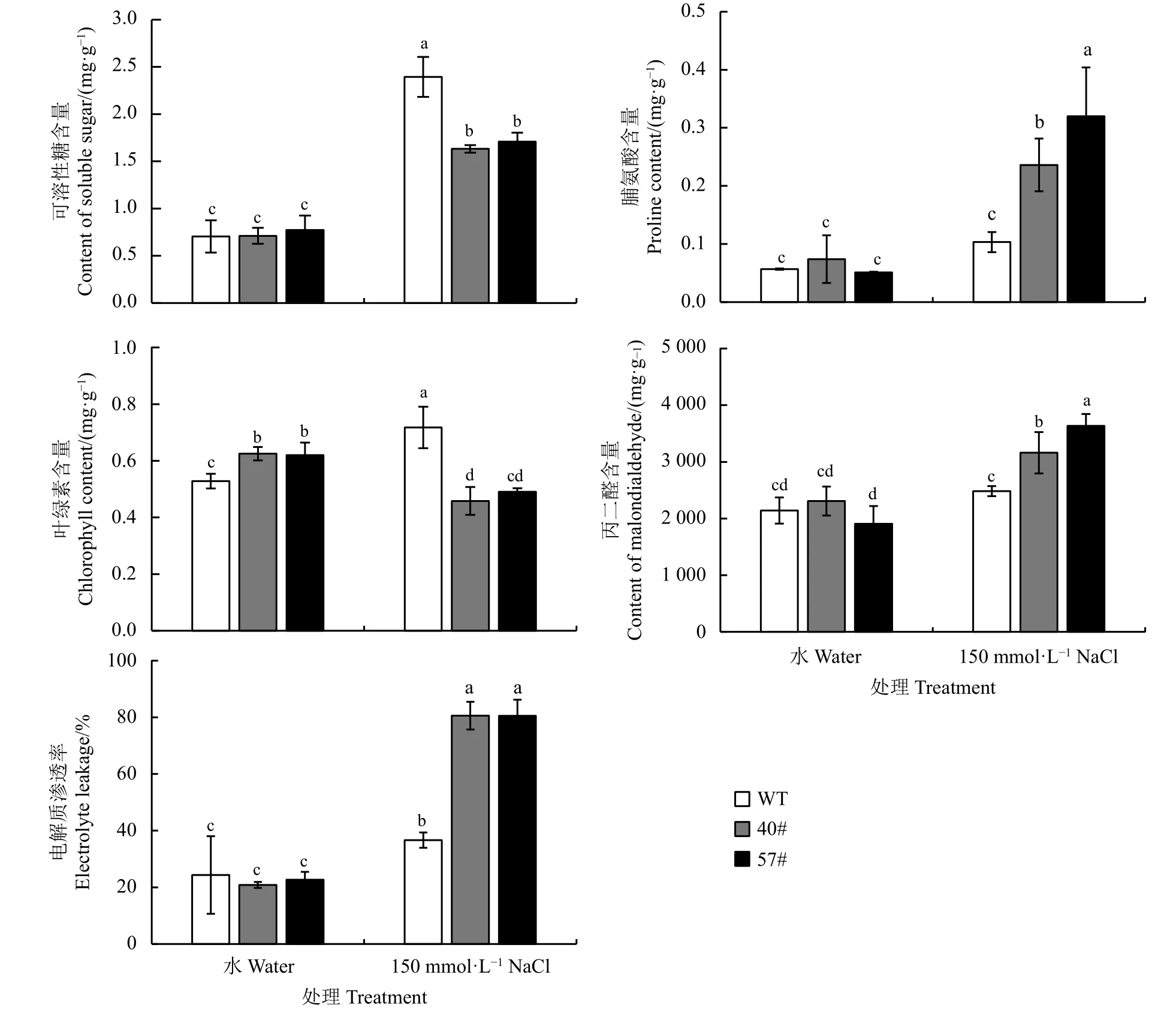
圖7 150 mmol·L?1 NaCl 處理7 d 后轉基因擬南芥葉片的生理變化Figure 7 Changes in the physiological indexes of transgenic Arabidopsis thaliana treated with 150 mmol·L?1 NaCl over a 7-day period
2.5 抗性基因的表達分析
鹽脅迫處理后,WT 中AtAPX1、AtPOD基因的表達水平無顯著變化,而AtNHX1基因的表達水平顯著高于對照(P< 0.05),約為對照的1.5 倍;轉基因植株中AtNHX1、AtAPX1基因的表達水平在鹽處理后分別低于對照1.5 倍和2 倍,AtPOD基因的表達水平顯著高于對照4 倍(P< 0.05) (圖8)。
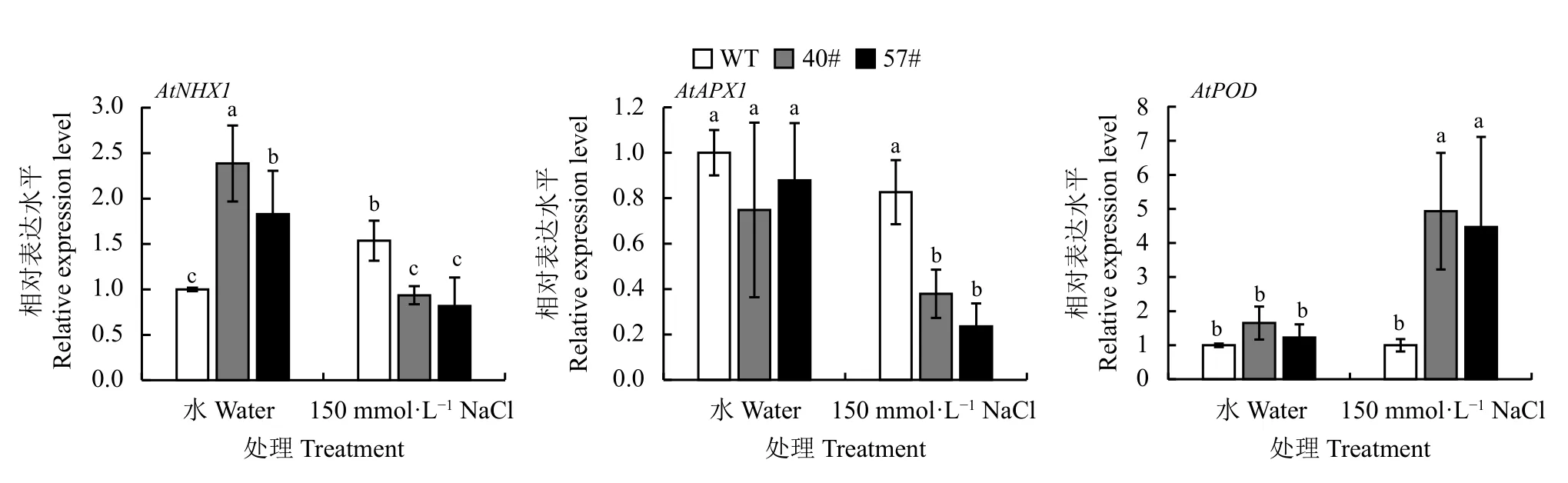
圖8 150 mmol·L?1 NaCl 處理后抗性基因的表達分析情況Figure 8 Expression of other salt resistance genes following treatment with 150 mmol·L?1 NaCl
3 討論
NAC 轉錄因子在調控植物鹽脅迫響應方面已有廣泛研究,其在鹽脅迫調控過程中發揮著重要作用。本研究探究了ZjNAC3對酵母YPH500 菌株的鹽敏感性,結果顯示,轉ZjNAC3的酵母細胞在鹽脅迫下長勢弱于對照,表明在鹽脅迫下ZjNAC3對酵母細胞的生長起負調控作用。
為進一步探究ZjNAC3基因對擬南芥植株耐鹽性的影響,本研究通過農桿菌侵染擬南芥的方法獲得了轉ZjNAC3基因擬南芥植株,并進行鹽脅迫處理。結果發現,鹽處理后轉基因植株中的葉綠素含量均顯著低于WT。葉綠素是植物光合作用所必需的色素,其含量降低影響植物的光合效率[20-21],推測轉ZjNAC3基因植株的光合作用受到鹽脅迫的明顯抑制。可溶性糖既是植物光合作用的產物[22],也常被作為滲透調節物質[23]。本研究中,鹽處理后,對照的可溶性糖含量顯著高于過表達株系,說明對照的滲透調節能力強于過表達株系。因此,滲透調節方面反映出過表達ZjNAC3基因植株的耐鹽性比WT 弱。
脯氨酸常被視為滲透調節物質,是判斷抗逆性的一種指標[24]。而近年來脯氨酸在植物抗逆中的作用存在一定爭議,如Yuan 等[25]研究發現,過表達Osa-miR396c的匍匐剪股穎(Agrostis stolonifera)的耐鹽性顯著提高,然而測定結果發現,轉基因株系中的脯氨酸含量顯著低于WT,這可能是脯氨酸的穩態比脯氨酸的積累對脅迫下植物的生長發育更重要。也有研究認為脯氨酸的含量是植物在脅迫條件下受損程度的一種指標,例如鹽敏感品種的水稻(Oryza sativa)在鹽脅迫下脯氨酸含量高于耐鹽品種,但其滲透調節能力較低[26]。本研究認為,脯氨酸是衡量脅迫受損的指標之一。鹽處理后轉基因植株中脯氨酸含量顯著高于WT,轉基因植株的受損程度高于WT。丙二醛含量和細胞膜透性也是衡量植
株受損程度的指標,鹽脅迫條件下植物內產生大量的活性氧自由基,細胞膜膜脂被氧化產生大量丙二醛;細胞膜結構被破壞,細胞膜透性增大,細胞內的電解質、有機物等外泄[27-30]。本研究發現在鹽處理后,過細胞膜透性和丙二醛的含量在過表達株系中明顯比WT 高,說明過表達株系的損傷更強。anac092-1突變體耐鹽性的研究結果與本研究的結果一致:鹽脅迫既增強了突變體種子的發芽能力,同時又抑制了葉片衰老;而對于過表達株系中,鹽脅迫對植物的生長發育產生了阻礙[8]。
過表達柳枝稷(Panicum virgatum)PvNAC1增強了芽和根中K+的積累,而抑制了Na+的積累,并誘導相關離子轉運蛋白基因的表達[9]。AtNHX1基因編碼液泡逆向轉運蛋白,可以將細胞液中的Na+隔離到液泡中,與植物抗性密切相關[31]。本研究中,在鹽脅迫處理后,AtNHX1基因的表達水平在轉基因植株中顯著低于WT,說明轉基因植株將Na+隔離到液泡中的能力弱于WT,這與Apse 等[31]和Zhang 等[32]的研究結果一致。AtAPX1和AtPOD基因分別編碼抗壞血栓過氧化物酶(APX)、過氧化物酶(POD),能夠減弱過量活性氧對細胞造成的損傷[33-34]。本研究中,鹽處理后轉基因植株中AtAPX1和AtPOD的表達水平,與WT 相比分別顯著降低和升高,抗氧化酶基因的表達水平不一致。馬長樂等[35]研究發現鹽脅迫下鹽地堿蓬(Suaeda salsa)中SsAPX的表達量增加;而劉香娥等[36]研究發現鹽脅迫下西瓜(Citrullus lanatus) POD 和APX 的合成受阻。本研究中,鹽脅迫處理與對照的WT 中AtAPX1和AtPOD基因表達水平未顯著性變化,表明WT 在經歷7 d的150 mmol·L?1NaCl 處理后,體內活性氧已到達穩定水平,抗氧化基因的調節不明顯。而轉基因植株體內活性氧發生明顯的改變,需要激活抗氧化酶基因的轉錄來維持活性氧的平衡,其中的機制需要進一步研究。
4 結論
為研究日本結縷草ZjNAC3在鹽脅迫中的功能,首先通過對轉ZjNAC3基因酵母菌株的鹽敏感測試,發現ZjNAC3基因減弱了酵母菌株的耐鹽性。進一步利用轉基因擬南芥株系進行了驗證,發現過表達ZjNAC3基因能夠使擬南芥植株的耐鹽性降低。分析發現ZjNAC3基因可通過減弱轉基因株系的滲透調節能力、增加細胞受損程度、降低將Na+隔離到液泡的功能等途徑參與鹽脅迫調控途徑。本研究為進一步研究結縷草中NAC 轉錄因子的調控機理奠定了基礎。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
