海洋沉積物中金屬依賴型甲烷厭氧氧化作用研究進展及展望
2021-11-05 02:13:32辛友志孫治雷王紅梅陳燁徐翠玲耿威曹紅張喜林張現榮李鑫閆大偉吳能友
海洋地質與第四紀地質 2021年5期
關鍵詞:研究
辛友志,孫治雷,王紅梅,陳燁,徐翠玲,耿威,曹紅,張喜林,張現榮,李鑫,閆大偉,吳能友
1. 中國地質大學(武漢)地球科學學院,武漢 430074
2. 自然資源部天然氣水合物重點實驗室,中國地質調查局青島海洋地質研究所,青島 266237
3. 青島海洋科學與技術試點國家實驗室海洋礦產資源評價與探測技術功能實驗室,青島 266237
4. 中國地質大學(武漢)生物地質與環境地質國家重點實驗室,武漢 430074
從20世紀70年代后期開始,隨著海洋生態學的研究由生物的種群、群落結構等靜態研究向生態功能和生態系統的物質轉化與能量循環等動態研究的轉變,越來越多的海洋微生物生態學的研究集中到微生物介導的元素循環上。海洋微生物不僅種類豐富、數量龐大,而且還具有多樣化的代謝途徑,在海洋碳、氮、硫等元素的生物地球化學循環中具有重要地位[1-2]。甲烷(CH4)是能夠引起溫室效應的主要氣體之一,它的循環對于氣候變化有重要調節作用[3]。據估計,全球海洋沉積物中約90%的甲烷氣體會通過AOM消耗[4-5],從而使甲烷從沉積物向海水和大氣的排放量保持在相當低的水平。
海洋環境是地球表面最大的生態系統之一,包括洋殼、海底沉積物以及冷泉、熱液、海山等多種地質結構[2]。海洋沉積物是由海水中的顆粒物質和生物殘骸不斷沉降并在海底聚積形成的特殊生態環境[3]。經過漫長地質時期的積累,海洋沉積物平均厚度達500 m,約覆蓋地球面積的48.6%[4],這為微生物的生存提供了巨大的場所。海洋沉積物中微生物的代謝活動取決于電子供體(可被氧化的單質或化合物)和電子受體(可被還原的單質或化合物)的可利用性和價態。考慮到數量和可利用性,硫酸鹽(SO42?)是AOM主要的電子受體。因此,在厭氧的海底沉積物中,AOM主要發生于硫酸鹽-甲烷轉換帶(sulfate-methane transition zone, SMTZ)[6-7]。但是在冷泉區,甲烷的上升通量很高,在SMTZ中并不能完全被消耗氧化。因此,海洋沉積物中微生物介導的AOM過程可能會涉及不同的電子受體,除硫酸鹽以外,針對能夠與AOM耦合的其他電子受體的研究也越來越多。目前,研究發現的類型包括硫酸鹽依賴型AOM(Sulfate-AOM)、金屬(鐵、錳、鉻)依賴型AOM(Fe-, Mn-, Cr-AOM)和硝酸鹽、亞硝酸鹽依賴型AOM(NO3?-, NO2?-AOM)[8-9](表1)。
早在1980年,Zehnder和Brock就提出了金屬離子可能也會參與AOM的假設[10]。因為僅根據吉布斯自由能規律(表1),AOM與金屬氧化物還原反應耦合要比與硫酸鹽還原耦合更具優勢[10]。沉積物滲漏實驗(leakage experiments)表明[11],硫酸鹽還原反應、鐵還原反應、AOM和甲烷生成作用可同時在海洋沉積物中發生,并且鐵氧化物的存在顯著促進了細菌硫酸鹽還原反應的速率。已有研究推測metal-AOM在地球早期缺氧、富鐵、貧硫的環境中可能會十分繁盛,甚至還推動了地質歷史演化[12]。錳和鐵的固體氧化物在深海沉積物中的氧化還原地球化學中起著重要作用。同時,金屬氧化物也存在多樣化的來源。例如,海底深部流體的排放過程既發生在空間受限的快速對流體中(如沿洋中脊分布的熱液噴口),也發生在大面積的海底(包括冷泉滲流等),其溫度低,擴散程度更大[2]。如果熱液和冷泉兩種截然不同的海洋生態環境相互毗鄰,那么兩者之間的關聯互作必然會影響metal-AOM的發生。
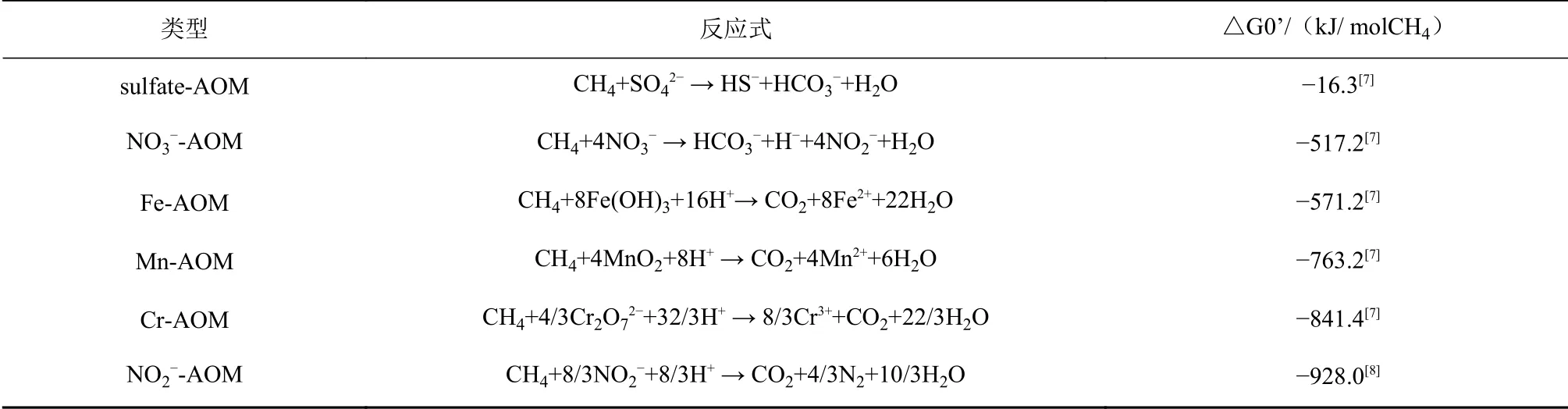
表 1 甲烷厭氧氧化作用類型及其標準吉布斯自由能(△G0’)Table 1 Standard Gibbs free energies(△G0’)of different AOMs
總而言之,在海洋極端環境中,控制微生物群落的環境梯度存在于從微米到千米的空間尺度,也存在于從毫秒到千年的時間尺度。目前對能夠介導metal-AOM的微生物類群依然認識不清,其介導的metal-AOM發生機制和重要意義還需要更加深入的研究。因此,本文從微生物生態學角度,綜述了海洋沉積物metal-AOM的研究進展,總結了具有介導metal-AOM潛能的微生物類群,概括了metal-AOM發生的地球化學證據,并以當前已經展開較多相關研究的西太平洋沖繩海槽為例,探討metal-AOM的發生機制模型,以期為未來研究提供方向和思路。
1 海洋沉積物中具有介導metal-AOM潛能的微生物
如果大量甲烷從海洋沉積物釋放到大氣,將會對全球氣候產生巨大溫室效應[13]。微生物既是海洋沉積物中甲烷的生產者同時又是甲烷的利用者。在缺氧的沉積環境,地球化學證據表明甲烷的消耗與硫酸鹽的消耗存在正向關聯[14]。同時,對微生物遺傳物質和脂類的研究,表明該互養關系(syntrophic relationship)是由硫酸鹽還原菌(sulfate-reducing bacteria,SRB)和甲烷厭氧氧化古菌(anaerobic methane-oxidizing archaea, ANME)介 導 的[15],并 且ANMEs(ANME-1,ANME - 2, ANME -3)和多個SRB類群都參與該過程[16]。海洋微生物催化AOM可將海底產生的75%的甲烷轉化為碳酸鹽沉淀,從而大大降低了向海洋水體釋放[17-18]。然而通過對墨西哥灣北部冷泉沉積物孔隙水的地球化學定量研究發現,現代海洋沉積物中AOM和硫酸鹽還原(sulfate reduction, SR)之間存在很大的不平衡[19]。實際上,盡管AOM與SR耦合已得到充分的證明,但近期的研究表明,在海洋熱液沉積物[20]等一系列自然環境中,可能存在由硝酸鹽、亞硝酸鹽和金屬氧化物(鐵、錳、鉻)還原驅動的AOM[21-28]。但是,迄今為止,在自然海洋環境中與metal-AOM相關聯的信息還十分匱乏。
目前已知的能夠執行AOM的微生物大多數屬于古菌域的ANMEs。基于16S rRNA基因系統發育研究表明,所有ANMEs都屬于廣古菌門(Euryarchaeota),并且與可純培養的產甲烷菌在進化上十分相近[29]。研究發現,甲烷八疊球菌目(Methanosarcinales)以及甲基桿菌屬(Methylobacter)微生物可能參與Kinneret湖泊沉積物Fe-AOM[27]。淡水生態系統NO3?-AOM主要是ANME-2d類群參與[29-30],后來發現該類群也能夠催化Fe-AOM[31]。近期,通過宏基因組高通量測序技術在ANME-2d類群中鑒定了一個新的微生物種屬(Candidatus Methanoperedems ferrireducens),該類微生物也可以生活在淡水沉積物,并參與Fe-AOM[32]。同樣,在對低硫、富鐵的湖泊沉積物進行富集培養和RNA穩定同位素探針(RNA stable isotope probing, RNA-SIP)研究中,再次表明ANME-2d類群是淡水生態系統Fe-AOM的主要參與者[33]。而在海洋沉積物中,ANMEs的生態位也有分化,ANME-2a / b通常在上部和中部占主導地位,而ANME-2c或ANME-1占據較深區域[34],這表明了ANMEs代謝能力的多樣性和特異性,也為海洋沉積物metal-AOM的發生提供了先決條件。
在海洋生態系統,證明微生物參與metal-AOM的證據之一是來自對Eel River Basin冷泉沉積物的研究。原始樣品和對照組相比,在使用錳(水鈉錳礦)作為電子受體進行富集培養10個月后,微生物群落結構發生了變化,該研究的作者認為,ANME-1或ANME-3及各自結伴細菌可能參與Mn-AOM[35]。之后通過熒光原位雜交和次級離子質譜測定聯用(fluorescence in situ hybridization coupled to secondary ion mass spectrometry, FISH-SIMS)技術,從形態學進一步確認ANME-2類群可能參與Mn-AOM富集培養過程[36]。此外,對Helgoland Mud 地區表層沉積物的研究表明,JS1細菌、產甲烷菌和ANME-3(甲烷鹽菌屬Methanohalobium)類群的古菌與沉積物中溶解鐵的分布存在正向相關性[37]。然而最近對Helgoland Mud 地區深部沉積物的富集培養研究表明,ANME-2a也可能在metal-AOM過程發揮關鍵作用[38]。對圣塔莫尼卡盆地冷泉沉積物的研究表明,向富集培養樣品添加Fe(III)化合物(檸檬酸鐵和乙二胺四乙酸鈉鐵)后,樣品中ANME-2a和ANME-2c豐度升高,而ANME-1豐度相對降低[39]。除了在冷泉區的研究,也有對高溫下熱液沉積物AOM進行的研究,富集培養結果表明ANME-1c類群可能是在有機質貧乏、含有金屬氧化物的熱液沉積物中Fe-AOM的主要參與者[20]。目前對metal-AOM的研究較多開展于淡水生態系統和海洋生態系統。在陸地泥火山環境中,也發現了ANME-2a和脫硫單胞菌屬(Desulfuromonas)/居泥桿菌屬(Pelobacter)占主導地位的metal-AOM[40]。
2 海洋沉積物中metal-AOM機理
自從在深部沉積地層發現存在微生物以來,有關支撐微生物生命能量來源和深部生物圈代謝狀態的疑問就一直備受關注。在海洋沉積環境,電子受體的利用能力通常體現為其最大電子接受潛能,當耦合的氧化還原反應在熱力學上有利并產生足夠的能量用于三磷酸腺苷酶(ATP)生成,微生物就能獲得能量。微生物根據熱力學能量定律來選擇環境中電子受體的利用順序,針對同一電子供體,具有最高還原勢能的電子受體會首先被消耗[2]。目前對海洋沉積物中電子供體的可利用性還不十分清楚。一般認為,海洋沉積物中微生物群落的豐度與沉積物有機碳含量呈正相關,這與在其他生境中觀察到的模式一致(即富含有機質的沉積物含有較高的細胞密度)[41]。除了有機碳,其他電子供體對海洋微生物也具有重要作用[42]。
在海洋沉積物中,已證實電子受體會按次序發生還原反應,具有較高還原勢能的電子受體會在沉積物淺層被消耗,并且這種規律也適用于其他深海環境。氧(由光合作用產生,從真光層通過全球海水循環輸送)通常是沉積環境中首選末端電子受體,其后分別是硝酸鹽(主要是在有機質降解過程中通過銨鹽的硝化作用形成)、亞硝酸鹽(通常由硝酸鹽或銨鹽產生)、錳和鐵的氧化物(Mn(IV)、Fe(III);源自熱液、河流或粉塵輸入)和氧化的硫化合物(硫酸鹽,主要來自大陸侵蝕;元素硫和硫代硫酸鹽,來自H2S的再氧化)[2](圖1)。其中,鐵氧化物在海洋沉積物主要通過兩種可能的途徑被還原,即鐵氧化物還原與有機質降解(organic matter degradation,OMD)過程耦合(Fe-OMD)和鐵氧化物還原與AOM耦合(Fe-AOM)。前者可能是最古老的呼吸形式之一,由Fe3+充當電子受體[43]。然而,這種途徑在很大程度上受到沉積物剩余總有機質的可利用性低和活性差的限制[44]。例如,在沖繩海槽北部,研究發現富鐵碳酸鹽巖周圍沉積物中活性有機碳含量非常低(0.5%),表明Fe-OMD可能受到很大限制[45-46]。同樣,對南海神狐海域的沉積物柱研究也證實存在類似的過程[47]。
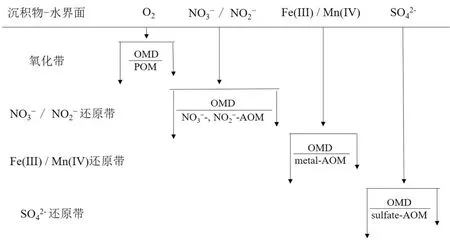
圖 1 電子受體在海洋沉積物中按序利用示意圖POM(particular aerobic oxidation of methane):甲烷有氧氧化,OMD(organic matter degradation):有機質降解;修改自文獻[2]。Fig.1 Sequential utilization of electron acceptors in marine sedimentsPOM :Particular aerobic oxidation of methane, OMD: Organic matter degradation; adapted from reference [2].
盡管在海洋沉積物中已發現金屬氧化物(鐵氧化物、錳氧化物)還原驅動的AOM[48],但微生物通過AOM偶聯金屬氧化物的還原反應來獲得能量依然面臨巨大挑戰。因為在海洋沉積物中,金屬氧化物以固態礦物的形式存在,難溶且難以被利用[49]。相對容易利用的金屬氧化物在富含甲烷區域之上就會被還原,而“幸存的”金屬氧化物則呈惰性形式,例如結合在層狀硅酸鹽中的含鐵礦物,可以保存數千年[50]。但是,在海洋沉積物柱樣品的深部甲烷生成區能夠觀察到還原形式的金屬離子(例如Fe2+和Mn2+)濃度增加,這表明該層已經發生金屬氧化物的還原反應[51]。此外,微生物在呼吸過程中將電子輸送到液態和固態金屬氧化物的機制也各不相同,有些微生物利用細胞壁上的酶通道,還有一些微生物則通過導電的細胞外分泌附屬物轉移電子[43, 51]。
ANME古菌均可以進行“反向產甲烷”途徑代謝,但電子轉移到金屬氧化物的過程仍是未知的。在大多數ANME類群中,多血紅素c型細胞色素(multi-heme c-type cytochromes, MHCs)發揮了傳導電子的作用。例如,在淡水沉積物富集培養過程添加Mn(IV)氧化物(水鈉錳礦)后,通過宏基因組和宏轉錄組分析技術發現,參與甲烷氧化和MHCs通路的許多關鍵基因都發生了上調表達[52]。鐵還原菌Geobacter sulfurreducens在Fe(III)氧化物和Mn(IV)氧化物還原過程中也是通過MHCs轉移電子[53],因此,也有可能參與metal-AOM。在實驗室培養環境,將異化鐵還原菌(奧奈達希瓦氏菌ShewanellaoneidensisMR-1)加入到反硝化甲烷厭氧氧化體系中(ANME-2d為其中主要類群)共培養,并以水鐵礦(ferrihydrite)為惟一電子受體條件下,也會發生AOM[54]。通過單細胞表征技術還發現,MHCs在ANME-2a/b和ANME-2d古菌細胞中的體積要大于G. sulfurreducens和S. oneidensis[55]。有研究推測,ANME-2d和Ferroglobus placidus可以將CxxCH蛋白塑造成細胞外導電結構或菌毛,從而進行固體鐵氧化物的還原[56]。這些研究表明,ANME-2d具有將AOM與硝酸鹽以外的其他電子受體還原反應偶聯的潛能,甚至包括不溶性金屬氧化物。此外,“Photoferrotrophy”被認為在早期地質歷史時期維持了Fe-AOM的發生[57]。除了直接與AOM偶聯,金屬氧化物還可以間接影響與其他電子受體還原反應耦合的AOM過程。研究發現,向水合物及冷泉沉積物添加赤鐵礦會導致鐵還原反應與sulfate-AOM速率同時增加[58]。然而,metal-AOM是由單個微生物類群介導發生的,還是由多個微生物類群協作完成的,仍需進一步的研究。
綜上所述,在不同生態環境發生的metal-AOM,可能存在不同的微生物功能群和代謝過程(表2)。但是目前還無法確定能夠介導metal-AOM的微生物類群,其具體的生物學過程及代謝機制也無法確定。對metal-AOM發生機理的研究在很大程度上還是受到現有技術方法的限制。近期在第三代測序和單細胞分選等技術上的突破將為海洋微生物研究提供新的方式,有助于揭示海洋極端環境中微生物代謝途徑和生存策略的多樣性。

表 2 不同生態環境中metal-AOM潛在功能群Table 2 Potential microbial communities of metal-AOM from different ecosystems
3 海洋沉積物中metal-AOM機制模型
就metal-AOM在沉積物不同層位發生的機制問題,目前已提出4種不同的模式(圖2A—D)。在富鐵的Matano湖有氧-厭氧界面以下,缺乏硫酸鹽、硝酸鹽和亞硝酸鹽,但富含鐵氧化物和錳氧化物,這為metal-AOM提供了金屬氧化物來源[23](圖2A)。此外,Sivan等[21]發現,如果金屬氧化物的還原反應在很大程度上受到了動力學和生物地球化學的限制,那么大量金屬氧化物就會被埋藏在深部缺氧區,同時,在產甲烷區以下只存在微量的硝酸鹽和硫酸鹽,metal-AOM則可在該區域內發生(圖2B)。相反,在丹麥日德蘭半島?rn湖泊沉積物頂部22 cm就發現了AOM信號,并且硫酸鹽和Fe(III)共存于此處[24](圖2C)。然而,上述3種metal-AOM發生類型尚未在海洋生態系統得到證實。
發生metal-AOM的基本前提是在其他電子受體虧損體系中同時存在大量可利用的CH4和金屬氧化物。在SMTZ以下,硫酸鹽、硝酸鹽和亞硝酸鹽的濃度通常非常低,因此,金屬氧化物可能成為AOM在該區域內惟一電子受體。以Aromokeye等[38]在Helgoland Mud地區的研究為例,孔隙水剖面數據顯示在低于SMTZ的樣品中檢測不到硫酸鹽(SMTZ深度:30~85 cm;檢測限:50 mM),而甲烷和可溶性鐵的濃度分別可高達6和380 mM。該研究結果可與之前Oni等[37]在該區域的研究互相印證,而且,對波羅的海沉積物的地球化學分析結果[59]也支持這種假設。可溶性鐵濃度升高可以指示鐵還原反應持續發生,鐵還原反應廣泛發生在海岸帶淺層沉積物(例如波斯尼亞海)和大陸架邊緣深部海底(例如阿根廷盆地)[38],該過程很可能與Fe-AOM相關。這些環境通常具有高沉積速率特征,從而便于將活性氧化鐵埋藏在SMTZ下。除了較高的沉積速率,人類活動造成的富營養化也會導致海岸帶沉積物中SMTZ抬升并將金屬氧化物埋藏在還原區內。根據富集培養實驗結果和數值模型估算得出Bothnian 海沉積物中Fe-AOM速率可達1.32 ±0.09 μmol·cm?3·a?1,約消耗甲烷總量的3%[60]。在對冷泉區碳酸鹽巖的研究中發現,通過同位素探針技術(包括 δ13C、δ34S和δ56Fe)發現metal-AOM應該是造成該區域沉積物中高濃度鐵、錳的主要途徑,并且富含金屬元素的礦物形成于SMTZ以下[45-47]。根據上述這些針對特定區域內metal-AOM研究的結果,可以發現,在海洋生態系統中,metal-AOM通常發生在SMTZ以下(圖2D)。然而,海洋沉積物中metal-AOM區是否也存在多種不同機制,還需要更多的研究來證明。總之,metal-AOM很可能是海洋沉積物中甲烷的另一個重要的匯。
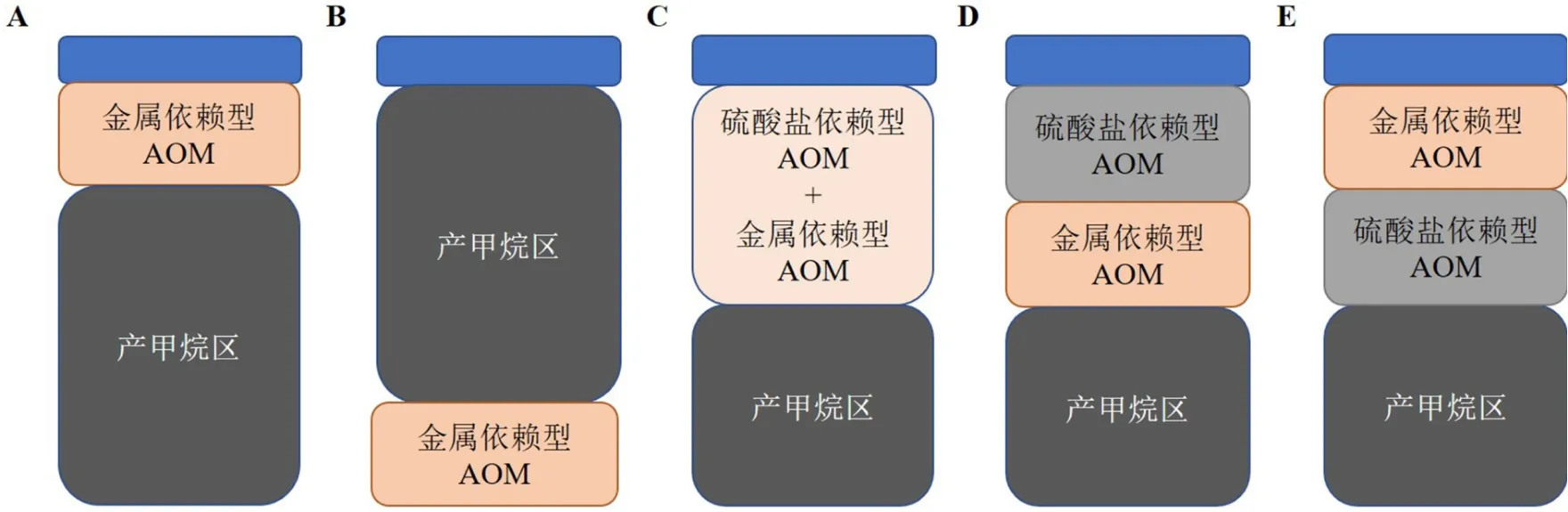
圖 2 沉積物中metal-AOM潛在發生區模型[12]A.位于產甲烷區以上(例:文獻[23]),B. 位于產甲烷區以下(例:文獻[21]),C. 位于SMTZ之中(例:文獻[24]),D. 位于SMTZ以下(例:文獻[37-38, 45-47, 59-60]),E. 位于SMTZ以上(本文依據沖繩海槽相關研究提出的假設);AOM(anaerobic oxidation of methane):甲烷厭氧氧化。Fig.2 Models of metal-AOM potential zones in sediments[12]A.metal-AOM occurs over the zone of methanogenesis(drawn based on the descriptions in reference [23]), B. metal-AOM occurs under the zone of methanogenesis(drawn based on the descriptions in reference [21]), C.metal-AOM occurs in the SMTZ(drawn based on the descriptions in reference [24]),D. metal-AOM occurs under the SMTZ(drawn based on the descriptions in reference [37-38, 45-47, 59-60])E. metal-AOM occurs above the SMTZ(drawn based on the researches of Okinawa Trough in this study); AOM(Anaerobic oxidation of methane).
研究海洋沉積物中metal-AOM發生機制的一個關鍵點在于活性金屬氧化物的來源問題。與其他水生生態系統相比,海洋中金屬氧化物存在多種外部輸入,包括粉塵、河流和熱液噴口等,它們還可以通過成巖作用在沉積物中累積[2,61-62]。Luo等[61]曾提出,來源于火山灰的鐵元素對Hikurangi margin沉積物中的碳循環有重大影響。在海洋沉積物SMTZ以下,以還原性含水形式存在的錳和鐵的濃度通常非常低。而在硝酸鹽還原區以下直到硫酸鹽還原區以上,還原性的錳、鐵濃度增加,反映出金屬氧化物的減少。產生的還原性錳、鐵又可在較淺的沉積物中被重新氧化或在較深的區域中與SR偶聯生成硫化物沉淀(如黃鐵礦)。在熱液流體與冷的氧化海水混合過程中也會產生錳和鐵的氧化物,這些氧化物可以促進熱液區周圍沉積物中微生物的代謝[63]。因此,海洋沉積物中活性金屬氧化物主要有兩種來源:深部沉積物和熱液羽流。先前的研究表明,在深部沉積物中,濁積作用可以促進鐵氧化物的輸入,將活性鐵氧化物埋藏在硫酸鹽還原帶以下[47]。這種事件在大陸坡等非穩態系統中應該是普遍存在的。然而,不可忽視的是,海洋環境中鐵氧化物的高輸入量很可能與熱液活動相關,特別是在熱液活動十分繁盛的地區,例如沖繩海槽。
沖繩海槽作為西太平洋一個典型的弧后盆地,發育了繁盛的熱液和冷泉活動[64-65]。2018年,青島海洋地質研究所利用FCV3000型水下遙控機器人在沖繩海槽內相距約50 km的冷泉和熱液噴口附近,分別發現了化能自養生態群落,并采集到在形貌上具有非常高相似度的貽貝、管蟲等。這兩地之間沒有任何的地形起伏和天然阻擋,難免會借助海水或相互連通的沉積物媒介發生交流或相互影響。同時調查發現,隨著采樣點和熱液噴口之間距離的減小,沉積物中鐵、錳、銅和鉻的濃度增加了2~7倍,并且當熱液噴口成簇出現時,活性金屬的輸出通量由于疊加而明顯增強[64]。因此,研究者認為,由于半封閉的地形特征以及冷泉和熱液系統的獨特共存,沖繩海槽擴張中心的現代熱液活動為其覆蓋的冷泉區沉積物提供了豐富的活性金屬[64-65]。沖繩海槽海底冷泉區沉積物中metal-AOM的發生很可能受到熱液輸入活性金屬的影響,并通過生物地球化學循環將兩個生態系統有機地聯系起來。
基于這些研究,本論文提出在沖繩海槽該特殊海洋生態環境沉積物中metal-AOM發生的機制模型(圖2E),其主要內容可概括為下述過程:熱液羽狀流攜帶大量的還原性金屬物質進入鄰近冷泉區,經海水氧化沉淀后形成活性金屬氧化物,并不斷在海底沉積;其次,冷泉區流體向上運移,那么冷泉流體中的甲烷將會和正常向下擴散的熱液活性金屬氧化物相遇,并在SMTZ上發生metal-AOM(圖2E)。熱液和冷泉地區分布有不同類型的電子供體和電子受體[66],這些差異直接導致了海洋微生物多樣性和生物地理分區研究常常是獨立進行的。一直以來,海底熱液和冷泉系統在生態學上通常被認為屬于“島狀生境(island-like habitats)”[67]。然而,熱液物質通過羽流源源不斷輸入冷泉區后很可能會發生一系列氧化還原反應,從而使熱液和冷泉區的元素循環過程發生耦合,而這一過程勢必會引起該區域微生物的生態響應,其過程和機制需要進一步的探索。
4 總結與展望
AOM在控制全球甲烷排放過程中起著至關重要的作用,海洋沉積物中metal-AOM在該過程中所扮演的角色仍需要詳細闡明。海底深部生物圈孕育了類型和生物量巨大并且代謝獨特的微生物[68-69]。海洋沉積物中微生物群落組成隨氧含量、碳含量、沉積物深度和熱液影響而變化。綜合前人研究,參與淡水生態系統中metal-AOM的微生物類群主要是ANME-2d,其介導metal-AOM的代謝機制也得到了初步鑒定。但是,巨大的空間以及生物地球化學過程和循環在空間和時間上的異質性對海洋微生物研究提出了重大挑戰。發掘海洋沉積物中能夠介導metal-AOM的微生物類群及其代謝機制仍需不斷完善研究技術。除了依靠傳統的純培養技術,新興的海底原位拉曼激光和流式單細胞分選技術也可能為海洋極端環境微生物研究打開突破口。
當前對metal-AOM發生機制的解釋多以淡水生態系統中的研究為主。這是由于海洋生態系統具有更加復雜的環境條件。同時,metal-AOM的產物(Mn(II)、Fe(II)和CO2)也會參與到其他金屬、硫和磷等元素的地球化學循環以及礦化過程。因此,metal-AOM對其所在生態系統及全球海洋元素循環的重要影響有待進一步的揭示。沖繩海槽具有熱液和冷泉系統共存的獨特特征,來源于熱液的相對還原的流體通過循環可為周圍貧電子受體的海底沉積物提供氧化劑,刺激微生物的活動。為沉積物中metal-AOM的發生提供了有利條件。為了進一步證實沖繩海槽海底沉積物中可能存在的metal-AOM發生機制,青島海洋地質研究所搭乘2020年“海洋地質9號”科考船對該區域進行了多方位取樣調查。同時,借助第三代微生物高通量測序技術及原位模擬富集培養技術,以發掘可能參與metal-AOM的微生物及相關代謝過程。
以沖繩海槽為代表的熱液-冷泉系統流體相互作用創新性理論的提出,從更廣闊的視角研究海洋生態系統之間相互作用及物質和能量的循環模式,為探究海洋沉積物中metal-AOM的發生機制提供了新的思路,并對海洋極端環境微生物的代謝途徑耦合和生命演化理論具有重要指導意義。不僅如此,除了在沖繩海槽等弧后盆地環境中存在metal-AOM,在開闊海和其他大陸邊緣,包括極地,都有可能存在metal-AOM。因此,今后的研究還應關注不同海洋環境中微生物的親緣關系、生態群落特征的對比,以及對不同地理和理化環境的適應性。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
