皇后帝王花種子休眠及解除方法研究
2021-11-08 12:09:12武懷燕郭志銘山金鳳陳婕吳向崇劉子嘉祝遵凌
熱帶作物學報 2021年9期
武懷燕 郭志銘 山金鳳 陳婕 吳向崇 劉子嘉 祝遵凌

摘? 要:皇后帝王花(Protea magnifica)種子具有休眠性,不經任何處理難以發芽。本研究以皇后帝王花種子為試驗材料,通過測定皇后帝王花種子生活力、種子萌發率、種子各部分粗提液對白菜種子發芽的影響,并以不同的方式(短時高溫+變溫層積、酸蝕處理+變溫層積、赤霉素處理+變溫層積)處理種子,研究皇后帝王花種子的休眠原因及解除方法。結果表明:試驗所用皇后帝王花種子的生活力高達90.00%,但不去除種皮的完整種子30 d內不發芽。種皮粗提液不僅抑制白菜種子的發芽,還顯著影響白菜種子的根與地上部分的生長。皇后帝王花種子的休眠可能是種皮的機械障礙和種皮的內源抑制物共同引起的綜合休眠。時長為2 min的不同高溫(40~120 ℃)處理+變溫層積、不同時間(10~ 70 min)的98%濃硫酸酸蝕處理+變溫層積、不同濃度(0.3、0.5、0.7 mg/L)的赤霉素溶液處理+變溫層積都可以打破種子的休眠,提高皇后帝王花種子發芽率和發芽勢。其中各處理分別以60 ℃的短時高溫處理、30 min的酸蝕處理、0.7 mg/L的赤霉素溶液處理效果最好。綜合比較,30 min的酸蝕處理為解除皇后帝王花種子休眠的最佳方法。此外,赤霉素處理對皇后帝王花種子的休眠解除有著一定的促進作用,但相比其他兩種處理方式效果并不理想。
關鍵詞:皇后帝王花;種子休眠;破除休眠;變溫層積
中圖分類號:S682.39????? 文獻標識碼:A
Causes of Protea magnifica Seed Dormancy and Methods for Breaking Dormancy
WU Huaiyan GUO Zhiming3, SHAN Jinfeng CHEN Jie WU Xiangchong3, LIU Zijia3, ZHU Zunling1,2,4*
1. Co-Innovation Center for Sustainable Forestry in Southern China, Nanjing Forestry University, Nanjing, Jiangsu 210037, China; 2. College of Landscape Architecture, Nanjing Forestry University, Nanjing, Jiangsu 210037, China; 3. Sanya Daxing New Landscape Ecological Co. Ltd, Sanya, Hainan 572022, China; 4. College of Arts & Design, Nanjing Forestry University, Nanjing, Jiangsu 210037, China
Abstract: The seeds of Protea magnifica have dormancy and it is difficult to germinate without any treatment. P. magnatica seeds were used as the experimental materials. The viability, germination rate and the effects of crude extracts from different parts of seeds on the germination of Chinese cabbage were measured, and seeds were treated with different methods (short-term high temperature + variable temperature stratification, acid etching + variable temperature stratification, gibberellin treatment + variable temperature stratification) to research the dormancy reason and release method of P. magnatica seeds. The results showed that: the viability of P. magnatica seeds was as high as 90.00%, the intact seeds without removing seed coat would not germinate within 30 days. The crude extract of seed coat not only inhibited the germination of Chinese cabbage seeds, but also significantly affected the growth of its roots and aboveground parts. The seed dormancy of P. magnatica may be a combinational dormancy caused by the mechanical barrier of seed coat and the endogenous inhibitor of seed coat. Different high temperature (40-120 ℃) treatment for 2 min + variable temperature stratification, different time (10-70 min) of 98% concentrated sulfuric acid etching + variable temperature stratification and different concentrations (0.3, 0.5, 0.7 mg/L) of gibberellin solution + variable temperature stratification could break the dormancy of the seeds, and improve the germination rate and germination potential of P. magnatica seeds. Among them, short-time high temperature treatment at 60 ℃, acid etching treatment for 30 min, and 0.7 mg/L gibberellin solution treatment were the best. Comprehensive comparison, acid etching for 30 minutes was the best way to release the seed dormancy. In addition, gibberellin treatment had a certain promoting effect on the dormancy release of P. magnatica seeds, but the effect was not ideal compared with the other two treatments.
Keywords: Protea magnifica; seed dormancy; dormancy breaking; variable temperature stratification
DOI: 10.3969/j.issn.1000-2561.2021.09.026
帝王花屬(Protea)植物隸屬山龍眼科(Proteaceae),全世界共有130種左右,非洲大陸擁有117種,南非就有82種[1-2],由于種子在國際間的引入時間較長,且種子具有休眠特性,其引種在我國還處于栽培試驗階段[3-4]。帝王花屬植物對南非的鮮切花產業具有極大的貢獻,其中出口量最大的為皇后帝王花(Protea magnifica)和闊葉帝王花(P. eximia)[5]。皇后帝王花為帝王花屬常綠直立或匍匐灌木[3],花兩性,頭狀花序錐形,具有色澤艷麗的大苞片,觀賞價值極高[6]。通過引進帝王花屬植物,能夠豐富我國鮮切花的種類,創造較高的經濟效益[7]。
帝王花屬植物的種子具有休眠特性,自然播種萌發率很低,需要較長的時間才能發芽[8-9]。曾力等[10]對皇后帝王花的種子進行了低溫恒溫處理和草木灰、赤霉素浸種處理,能夠顯著提高種子的發芽率與發芽勢。劉艷梅等[11]利用3種濃度的木醋液浸泡帝王花(P. cynaroides)種子,顯著提高了其發芽率。但目前對于皇后帝王花種子休眠的原因及解除方法的系統研究還較少。筆者課題組前期就皇后帝王花種皮特性對種子休眠的影響也進行了探究,本試驗以皇后帝王花種子為材料,通過對種子生活力、種子萌發特性、種皮粗提液對白菜種子發芽的影響等方面的研究,分析皇后帝王花種子休眠的原因。并用不同的方式組合一起處理種子,探索種子休眠的解除方法,以期打破皇后帝王花種子的休眠,從而縮短種子發芽時間,提高種子發芽率,為其在我國的有性繁殖及推廣應用提供理論和技術基礎。
1? 材料與方法
1.1? 材料
以2017年12月由哥斯達黎加引進的皇后帝王花種子為試驗材料,去除癟粒、腐壞的種粒及雜質,取新鮮飽滿的帝王花種子備用。白菜(Brassica campestris)種子為‘上海青四季小白菜,純度≥95%,凈度≥98%,含水量≤8%,發芽率≥80%。
1.2? 方法
1.2.1? 種子生活力測定? 采用氯化三苯基四氮唑(TTC)染色法測定種子生活力。隨機取30粒種子,重復3次。首先,將待測皇后帝王花種子用溫水浸泡24 h,剝去種皮,將去皮種子放入培養皿中,加入剛好淹沒種子的0.5%TTC溶液,然后將培養皿置于30 ℃恒溫培養箱內避光染色10 h,參考李炎坤等[12]的判定標準,在染色的第2、4、6、8、10 h觀察皇后帝王花種子的染色情況。最后,將子葉已著色的種子分成兩半,觀察內部染色程度,統計并計算最終生活力。
1.2.2? 種子萌發試驗? 選取完整的皇后帝王花種子(處理A)、去除外種皮的皇后帝王花種子(處理B)、去除內外種皮的皇后帝王花種子(處理C)各30粒,重復3次。將種子分別置于鋪有濾紙的發芽盒中,加蒸餾水后放在25 ℃的人工智能氣候箱內培養,若胚根能突破種皮或胚根伸長1 mm就認為可以發芽,記錄各處理種子30 d內的發芽情況。
1.2.3? 種子各部分粗提液生物活性測定? 取新鮮飽滿的皇后帝王花種子80粒,將種皮、種胚分別磨碎,稱取磨碎的種皮(約1.5 g)、磨碎的種胚(約2 g),參考劉文瑜等[13]的方法制備成粗提液。將種子各部分甲醇粗提液分別稀釋為原液的25%、50%、75%、100%,保存在4 ℃冰箱中備用。參考劉文瑜等[13]的方法,測定種子各部分粗提液生物活性,統計白菜種子發芽率,測量苗高與根長。
1.2.4? 種子休眠的解除? (1)短時高溫+變溫層積處理。將皇后帝王花種子攤平置于40、60、80、100、120、140 ℃烘箱中的玻璃培養皿內處理2 min。每皿30粒種子,重復3次。2 min后,立即取出放入燒杯中冷卻,以常溫(25 ℃)未短時高溫處理的皇后帝王花種子為對照,將種子置于25 ℃恒溫狀態下溫水浸泡24 h。
(2)酸蝕處理+變溫層積處理。將皇后帝王花種子用濃硫酸(98%)分別酸蝕處理0(CK)、10、30、50、70 min,然后置于流水中沖洗,并將種子在25 ℃恒溫狀態下溫水浸泡24 h。每個處理重復3次,每個重復30粒種子。
(3)赤霉素浸種+變溫層積處理。將皇后帝王花種子用濃度為0(CK,只加蒸餾水)、0.1、0.3、0.5、0.7、1.0 mg/L的赤霉素溶液于25 ℃恒溫狀態下溫水浸泡24 h。每個處理3個重復,每個重復30粒種子。
將各處理后的種子與消毒濕沙以1∶3的體積比混合,放入人工智能氣候箱(光照10 h溫度20 ℃/黑暗14 h溫度10 ℃)中層積40 d。定期補水并翻動沙子,以保持沙子濕潤和通氣良好。從層積的第15天起,每天下午3:00觀察并記錄種子的萌發數量,并將已萌發的種子移出自封袋進行播種,層積40 d后記錄結束,并計算最終發芽率、發芽勢。計算公式為:
發芽率=規定時間內正常發芽種子數/種子總數×100%
發芽勢=達到高峰時正常發芽種子數/種子總數×100%
1.3? 數據處理
數據采用Excel 2010軟件進行整理,SPSS 24.0軟件進行差異顯著性分析,Excel 2010軟件作圖。
2? 結果與分析
2.1? 種子生活力
經TTC染色法測定的皇后帝王花種子,有生活力的高達90.00%,喪失生活力的有7.67%,腐壞或空粒的種子占2.33%。綜合來看,試驗所用的皇后帝王花種子具有較高的萌發潛力,種子生活力本身不是影響種子發芽的主要因素。
2.2? 種子萌發試驗
由表1可知,A處理種子在30 d內均未發芽,B處理種子發芽率為25.56%,C處理種子因為去除了內、外種皮,子葉變軟,容易發霉,導致發芽率(8.89%)不高。這說明皇后帝王花種皮的某些特性是導致種子休眠的原因。
2.3? 種子各部分粗提液生物活性
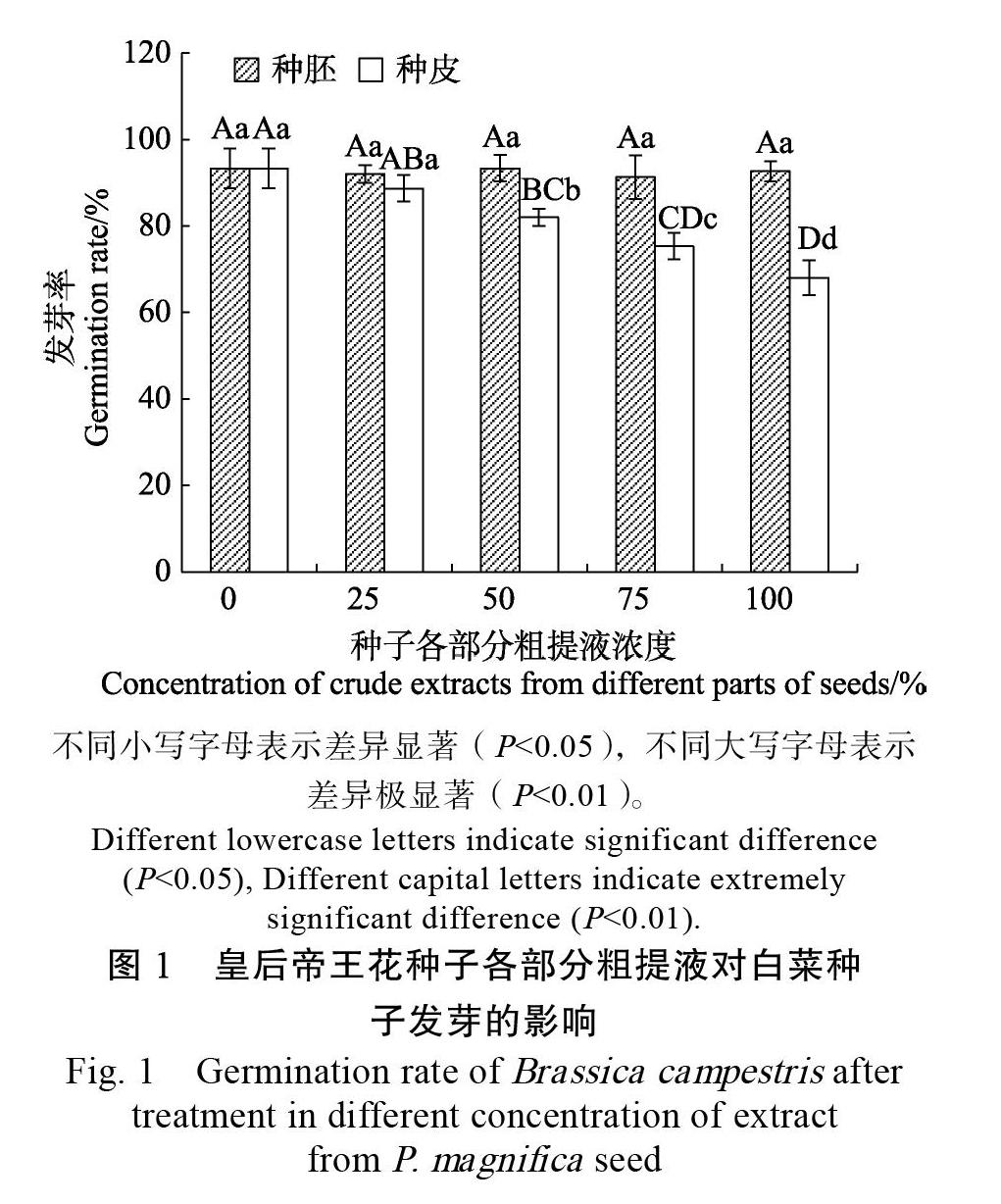
2.3.1? 種子各部分粗提液對白菜種子發芽率的影響? 由圖1可看出,皇后帝王花種子不同部位的粗提液對白菜種子發芽的影響不同,與CK相比,種胚粗提液對白菜種子的發芽率影響差別不大,而種皮粗提液對白菜種子發芽率的影響非常明顯。隨著種皮粗提液濃度的升高,對白菜種子萌發的抑制效果越來越強,當種皮粗提液濃度為100%時,白菜種子的發芽率僅為68%,比CK降低了25.33%。與CK相比,濃度為25%的種皮粗提液對白菜種子發芽率的影響未達到顯著差異 (P>0.05),而其他各濃度的種皮粗提液對白菜種子發芽率的影響均達到極顯著水平(P<0.01),可初步確定皇后帝王花種皮中含有抑制白菜種子發芽的物質。
2.3.2? 種皮粗提液對白菜苗高與根長的影響? 由圖2可看出,不同濃度的皇后帝王花種皮粗提液對白菜苗高與根長的影響均達到極顯著水平(P<0.01)。與CK(12.09 mm,18.09 mm)相比,采用100%濃度的種皮粗提液處理后的白菜苗高僅為5.81 mm,根長僅為9.45 mm。并且,皇后帝王花種皮粗提液對白菜苗高、根長的抑制作用隨著種皮粗提液濃度的升高而增強。由此可知,皇后帝王花種皮粗提液中存在的抑制物質不僅影響著白菜種子的生根,還影響它地上部分的生長。
2.4? 種子休眠的解除
2.4.1? 短時高溫+變溫層積處理對種子萌發的影響? 采用時長2 min的短時高溫處理再變溫層積的方法對皇后帝王花種子破除休眠進行試驗,結果表明,適宜的短時高溫處理對皇后帝王花種子萌發具有促進作用。從圖3可見,采用時長2 min的60 ℃高溫處理,皇后帝王花種子的發芽率(70%)、發芽勢(40%)均最高,對比CK(17.78%,7.78%),其發芽率提高了52.22%、發芽勢提高了32.22%;從60 ℃開始,隨著溫度的升高皇后帝王花種子的發芽率逐漸降低;當溫度達到140 ℃時,發芽率為0,說明時長2 min的140 ℃高溫已經是皇后帝王花種子的耐受極限,讓其喪失了生活力。40、60、80、100 ℃短時高溫處理的皇后帝王花種子發芽率、發芽勢與CK相比有著極顯著的差異(P<0.01)。可見,溫度對皇后帝王花種子休眠的解除至關重要,適當的增溫處理可以有效提高皇后帝王花種子的發芽率和發芽勢。
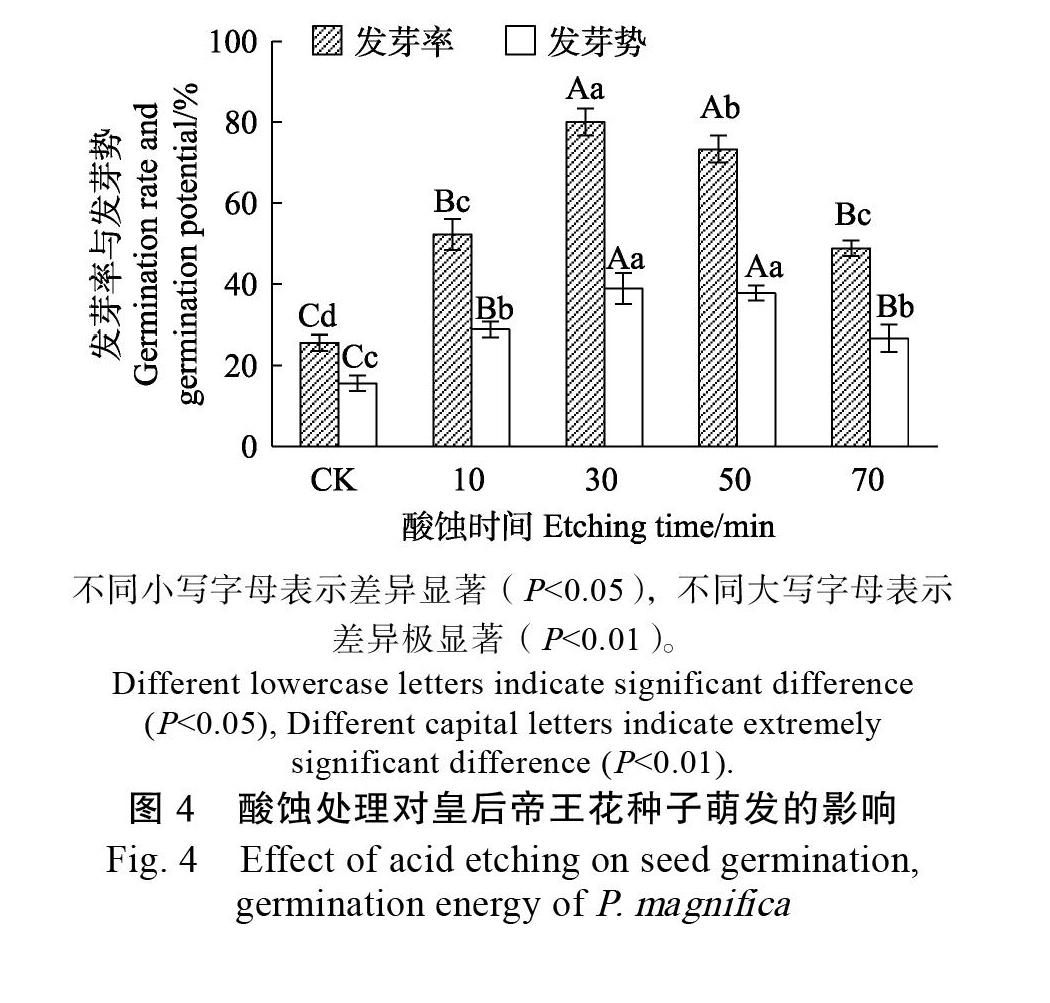
2.4.2? 酸蝕處理+變溫層積處理對種子萌發的影響? 濃硫酸處理會使皇后帝王花種子的種皮結構疏松,從圖4可見,經酸蝕處理的種子無論是發芽率還是發芽勢都有所提高,不同酸蝕時間對皇后帝王花種子發芽率、發芽勢的影響均達極顯著水平(P<0.01)。酸蝕30 min的皇后帝王花種子發芽率(80.00%)、發芽勢(38.89%)達最高,與CK(25.56%,15.56%)相比,分別提高了54.44%、23.33%;而隨著酸蝕時間的延長,處理50、70 min的種子發芽率與發芽勢逐漸降低;酸蝕70 min的皇后帝王花種子在層積過程中更容易腐爛。因此控制好皇后帝王花種子的酸蝕時間, 能夠減少對種子內部的傷害,從而有效提高種子的發芽率和發芽勢。
2.4.3? 赤霉素浸種+變溫層積處理對種子萌發的影響? 從圖5可見,經不同濃度的赤霉素溶液浸種后再變溫層積的皇后帝王花種子發芽情況整體來看差異不大,但與CK相比其他各處理的種子發芽率與發芽勢略有提升。經0.7 mg/L的赤霉素溶液處理的皇后帝王花種子的發芽率最高(35.56%),比CK(17.78%)提高了17.78%;經0.5 mg/L的赤霉素溶液處理的皇后帝王花種子的發芽勢最高(21.11%),比CK(11.11%)提高了10.00%;當赤霉素溶液濃度達到0.7 mg/L時,隨著濃度繼續增大,皇后帝王花種子的發芽率、發芽勢反而降低,說明較高濃度的赤霉素溶液會對皇后帝王花種子的萌發有一定的抑制作用,可見,適當濃度的赤霉素溶液才可以促進皇后帝王花種子的萌發。綜合發芽率與發芽勢情況來看,采用0.7 mg/L的赤霉素溶液處理的皇后帝王花種子萌發效果最好。
2.4.4? 綜合比較? 綜合比較(圖6),可看出這3種處理方式的種子發芽情況均比CK好,酸蝕處理30 min為最佳,但發芽勢比短時高溫60 ℃處理的略低。方差分析顯示,它們對皇后帝王花種子發芽率(F=206.42,P<0.01)、發芽勢(F=45.88,P<0.01)的影響均達極顯著水平。赤霉素處理對皇后帝王花種子休眠的解除有著一定的促進作用,但相比其他兩種處理方式效果并不理想。
3? 討論
3.1? 皇后帝王花種子的休眠原因
種子休眠是植物在長期系統的發育過程中,為了適應環境、保證自身的繁衍而形成的一種生物學特性[14-15]。皇后帝王花花朵巨大、色彩絢麗、觀賞期長,是我國近年來炙手可熱的觀賞植物材料。種子硬實,使得皇后帝王花在大火頻發的南非得以留存,但這卻是其種子繁殖的一大障礙[7]。從哥斯達黎加引進的皇后帝王花種子,雖然種子生活力高達90.00%,但不經任何處理很難發芽。皇后帝王花種子的種胚能夠在30 d內萌發,帶有內種皮的種胚亦能萌發,但完整種子卻不能萌發,這說明皇后帝王花種子的種皮具有休眠特性。皇后帝王花種子的種皮堅硬厚實、結構致密,導致皇后帝王花種皮存在機械障礙[16]。用98%的濃硫酸對皇后帝王花種子進行不同時間的處理,均可極顯著提高種子的發芽率與發芽勢,也證明了這一點。種子的不同部位存在的一些抑制物是種子休眠的重要原因之一[17]。內源抑制物在一些具有休眠特點的種子中很常見,酸櫻桃(Prunus cerasus)種子的胚休眠是由子葉中的內源抑制物引起的,去除部分子葉可消除內源抑制物對萌發的阻礙,使幼苗得以正常生長[18]。研究表明,紫斑牡丹(Paeonia rockii)[19]、滇重樓(Paris polyphylla)[20]、金絲李(Garcinia paucinervis)[15]、野生黃瑞香(Daphne giraldii)[17]、麻花秦艽(Gentiana straminea)[21]以及烏桕(Sapium sebiferum)[22]等物種的種子中也都含有抑制種子萌發的物質。通過對皇后帝王花種子各部分粗提液進行生物活性測定后發現,其種胚中無抑制物,而種皮粗提液中含有抑制白菜種子發芽的物質,該抑制物還顯著影響著白菜種子的根與地上部分的生長,并且隨著種皮粗提液濃度的升高,抑制作用越發明顯。種子中存在的內源抑制物種類很多,如脫落酸、香豆素、酚類、醛類等物質[23-24]。本試驗雖已證明皇后帝王花種子的種皮粗提液中含有抑制種子萌發的內源抑制物,但具體為哪類物質還有待鑒別。綜上可知,皇后帝王花種子的休眠是種皮障礙和種皮的內源抑制物共同引起的。
3.2? 皇后帝王花種子解除休眠的方法
植物種子對高溫處理比低溫處理更加敏感[25],萌發前對一些植物種子進行一定時間的高溫處理可以改變種皮結構,促進蛋白質的合成,增加種子萌發相關酶活性,從而促進種子萌發[26-28]。余嬌娥等[27]對黃茅(Heteropogon contortus)種子進行不同溫度和時長的預處理,發現在80 ℃下處理20 min的種子發芽情況最好。張輝紅等[28]對4種松樹種子進行不同高溫不同時間的處理,發現60~120 ℃以內的短時高溫處理均能提高馬尾松(Pinus massoniana)與高山松(P. densata)種子的發芽率。對于不同植物的種子,因個體差異所需的溫度有所不同,所以在實踐操作過程中,找到適宜的溫度范圍及處理時間很重要。溫度太高或熱處理時間太長,都有可能引起種子內部酶活性降低,導致蛋白質變性,最終喪失生活力。于皇后帝王花種子而言,短時高溫處理能夠有效的提高種子發芽率,當溫度達到60 ℃時其發芽率達70%、發芽勢達40%。當溫度升高到140 ℃時,在相同的變溫層積時間內種子全部未萌發,說明140 ℃高溫讓其喪失了生活力。由此可見,皇后帝王花種子對溫度具有較高的耐受性,這一特性對其在大火頻發的南非得以留存有著很好的幫助。
濃硫酸處理可有效打破種子的物理休眠[29-32],經濃硫酸處理后,皇后帝王花種子表面結構疏松,角質層、柵欄層逐漸被腐蝕,形成大小不同的孔洞,減小了種皮的機械束縛。試驗表明,經不同酸蝕時間處理的皇后帝王花種子,發芽時間比CK提前,發芽率、發芽勢也大幅度提升。但酸蝕時間過長容易造成皇后帝王花種子內部結構受損,受損種子在潮濕的層積環境下更容易腐爛。對于本試驗而言,皇后帝王花種子的酸蝕時間以30 min(發芽率80.00%、發芽勢38.89%)左右為最佳。
外源赤霉素可以幫助種子增加種胚活力,弱化胚根周圍組織來促進種子解除休眠[33],同時還與種子休眠有關的脫落酸有拮抗作用[34]。劉文瑜等[13]發現用濃度為0.1 mg/L的赤霉素處理蒺藜苜蓿(Medicago truncatula)種子后再低溫層積可以顯著提高種子發芽率,李淑嫻等[22]研究發現烏桕先用赤霉素(500或1000 mg/L)處理,再層積100 d可以有效解除種子休眠。卓露等[35]研究發現200 mg/L赤霉素能夠顯著提高喜鹽鳶尾(Iris halophila)種子的萌發。本試驗利用不同濃度的赤霉素溶液對皇后帝王花種子進行浸泡處理,發現各處理的發芽率、發芽勢差異不大,但與CK相比其他各處理的發芽率與發芽勢略有提升。經0.7 mg/L的赤霉素溶液處理的皇后帝王花種子的發芽率最高(35.56%),經0.5 mg/L的赤霉素溶液處理的皇后帝王花種子的發芽勢最高(21.11%)。濃度繼續升高時,皇后帝王花種子的發芽率、發芽勢反而降低,說明較高濃度的赤霉素溶液不利于皇后帝王花種子的萌發。綜合發芽率與發芽勢情況來看,采用0.7 mg/L的赤霉素溶液處理皇后帝王花種子的萌發效果最好。
參考文獻
[1]? 艾尼瓦遜, 托尼羅德, 包志毅. 世界園林喬灌木[M]. 北京: 中國林業出版社, 2004: 302-322.
[2]? Vogts M. South Africas proteaceae: Know them and grow them[M]. South Africa: Struik Publishers, 1982.
[3]? 樊則郁, 鄧莉蘭, 樊國盛. 中國山龍眼科植物分類與分布[J]. 西南林學院學報, 2009, 29(1): 1-7.
[4]? 曾? 力, 樊國盛, 段曉梅. 3種山龍眼科植物種子生活力快速測定方法的效果比較[J]. 西部林業科學, 2007(4): 119-122.
[5]? Martapizano, 遠 飛. 南非: 現代花卉的搖籃[J]. 中國花卉園藝, 2003(19): 38-39.
[6]? Watson D P, Parvin P E. Ornamental Proteas: new cut flower crop[M]. Hawaii: Hawaii Agricultural Experiment Station, 1973.
[7]? 曾 力. 六種山龍眼科木本切花植物引種繁殖的研究[D]. 昆明: 西南林學院, 2008.
[8]? Kraaij T, Cowling R M, van Wilgen B W, et al. Vegetation responses to season of fire in an aseasonal, fire-prone fynbos shrubland[J]. Peer J, 2017, 5(8): 1-21.
[9]? Deall G B, Brown N A C. Seed germination in Protea magnifica Link[J]. South African Journal of Science, 1981, 77(4): 175-176.
[10]????? 曾? 力, 樊國盛, 段曉梅. 不同預處理對2種山龍眼科種子出苗率的影響[J]. 福建林業科技, 2009, 36(3): 141-144.
[11]????? 劉艷梅, 王曉旭, 汪曉峰, 等. 木醋液對帝王花種子萌發的影響[J]. 浙江林業科技, 2012, 32(4): 51-53.
[12]????? 李炎坤, 卓一南, 曾湘達, 等. 崗梅種子生活力測定方法的優化[J]. 種子, 2018, 37(11): 128-131.
[13]????? 劉文瑜, 魏小紅, 許可成, 等. 蒺藜苜蓿種子休眠機制及其破除方法研究[J]. 草地學報, 2015, 23(2): 358-365.
[14]????? Finch-Savage W E, Leubner-Metzger G. Seed dormancy and the control of germination[J]. New Phytologist, 2006, 171(3): 501-523.
[15]????? 張俊杰, 韋? 霄, 柴勝豐, 等. 珍稀瀕危植物金絲李種子的休眠機理[J]. 生態學雜志, 2018, 37(5): 1371-1381.
[16]????? 山金鳳, 劉子嘉, 曾? 瑩, 等. 皇后帝王花種皮特性對種子休眠的影響[J]. 熱帶作物學報, 2019, 40(1): 67-72.
[17]????? 閆? 芳, 張恩和, 王勤禮, 等. 野生黃瑞香種子的休眠機制與破除方法[J]. 林業科學, 2016, 52(4): 30-37.
[18]????? Jensen M, Kristiansen K. Removal of distal part of cotyledons or soaking in bap overcomes embryonic dormancy in sour cherry[J]. Propagation of Ornamental Plants, 2009, 9(3): 135-142.
[19]????? 李婉茹, 唐? 紅, 何麗霞. 紫斑牡丹種子浸提液對植物種子萌發和幼苗生長的影響[J]. 東北林業大學學報, 2020, 48(3): 8-12.
[20]???? 魏玉蘭, 毛美琴, 趙? 燕, 等. 滇重樓種子水浸液對三種植物種子萌發和幼苗生長的影響[J]. 植物科學學報, 2018, 36(1): 94-102.
[21]????? 李兵兵, 魏小紅, 徐? 嚴. 麻花秦艽種子休眠機理及其破除方法[J]. 生態學報, 2013, 33(15): 4631-4638.
[22]????? 李淑嫻, 劉菁菁, 田樹霞, 等. 烏桕種子休眠原因及解除方法研究[J]. 南京林業大學學報(自然科學版), 2011, 35(5): 1-4.
[23]????? 賈彩霞. 核桃種子內源抑制物質的初步研究[D]. 楊凌: 西北農林科技大學, 2006.
[24]????? 張艷杰. 南方紅豆杉種子休眠機理的研究[D]. 南京: 南京林業大學, 2007.
[25]????? 索風梅, 張? 昭, 陳? 瑤, 等. 不同溫度處理條件下植物種子萌發的研究進展[J]. 世界科學技術-中醫藥現代化, 2017, 19(4): 706-710.
[26]????? 李昭華, 郭超凡, 劉? 婕, 等. 不同溫度處理對疏序黃荊種子萌發的影響[J]. 種子, 2020, 39(9): 1-7.
[27]????? 余嬌娥, 吳雪濤, 司宏敏, 等. 萌發前高溫處理對黃茅種子萌發率的影響[J]. 種子, 2018, 37(10): 26-30+35.
[28]????? 張輝紅, 孟麗媛, 余? 婷, 等. 短時高溫處理對4種松樹種子萌發的影響[J]. 種子, 2018, 37(1): 32-37.
[29]????? 薛婷婷, 沈永寶, 劉? 嘉, 等. 種子物理休眠研究進展[J]. 植物學報, 2016, 51(6): 863-871.
[30]????? 王小雪, 孫海菁, 劉? 蕓, 等. 濃硫酸處理對海濱木槿10個家系種子萌發的影響[J]. 應用生態學報, 2012, 23(11): 2968-2974.
[31]????? 賈? 娟, 劉? 芳, 蘇紅田, 等. 不同處理方法對5種豆科牧草種子萌發特性的影響[J]. 草地學報, 2012, 20(2): 342-347.
[32]????? 侯文煥, 趙艷紅, 唐興富, 等. 不同處理方法對菜用黃麻種子萌發的影響[J]. 熱帶作物學報, 2018, 39(2): 231-236.
[33]????? Kucera B, Cohn M A, Leubner-Metzger G. Plant hormone interactions during seed dormancy release and germination[J]. Seed Science Research, 2005, 15(4): 281-307.
[34]????? Gomez-Cadenas A, Zentella R, Walker-Simmons M K, et al. Gibberellin/abscisic acid antagonism in barley aleurone cells: Site of action of the protein kinase PKABA1 in relation to gibberellin signaling molecules[J]. Plant Cell, 2001, 13(3): 667-679.
[35]????? 卓? 露, 管開云, 李文軍, 等. 喜鹽鳶尾種子休眠與萌發特性初步研究[J]. 干旱區研究, 2014, 31(4): 739-743.
責任編輯:沈德發
