大豆與野生大豆部分同源基因間基因置換分析
2021-11-08 08:21:34郭赫王希胤
種子科技 2021年16期
關鍵詞:大豆
郭赫 王希胤

摘? ? 要:以大豆和野生大豆為研究對象,利用系統(tǒng)發(fā)育分析比較推斷其最近一次加倍事件所產(chǎn)生的同源基因間發(fā)生基因置換的規(guī)模。結果表明,大豆中有37對同源基因間發(fā)生了基因置換,野生大豆中有33對同源基因發(fā)生了基因置換。通過對基因置換與基因在染色體上的位置關系研究發(fā)現(xiàn),靠近染色體兩端的基因更容易發(fā)生基因置換。
關鍵詞:大豆;野生大豆;基因置換
文章編號: 1005-2690(2021)16-0002-03? ? ? ?中國圖書分類號: S565.1? ? ? ?文獻標志碼: B
大豆(Glycine max)是重要的經(jīng)濟作物和油料作物。大豆經(jīng)歷過豆科共有的四倍體事件(Legume-common tetraploid,LCT)后又經(jīng)歷過一次大豆獨有的四倍體事件(Soybean-specific tetraploid,SST)[1],是同源四倍體。目前,大豆與野生大豆(Glycine soja)的全基因組測序工作已經(jīng)完成[2-3]。對大豆與野生大豆于最近一次全基因組加倍事件產(chǎn)生的同源基因間進行比較研究,有利于理解其基因組的進化。
全基因組加倍(whole genome duplication,WGD)即多倍化是基因復制的主要方式之一,植物的進化過程中多倍化是反復發(fā)生的,多倍化產(chǎn)生的大量重復基因為新功能的進化提供了原材料[4]。全基因組加倍后發(fā)生的染色體重組、基因丟失等現(xiàn)象對基因組結構及功能都造成很大影響[5]。
遺傳重組(genetic recomnnation)作為生物進化的主要推動力之一,對DNA序列損傷的修復和同源序列間信息的傳遞有著重要意義[6]。很多物種多倍化后產(chǎn)生的同源序列之間的同源重組與經(jīng)典的同源片段間重組不同,稱為非正常遺傳重組[7]。單向的一個基因替換其同源基因的過程稱為基因置換(gene conversion)[8-9]。
1? ?材料和方法
1.1? ?物種基因組數(shù)據(jù)
豆科植物大豆和野生大豆的全基因序列由公共數(shù)據(jù)庫NCBI(https://www.ncbi.nlm.nih.gov/)下載獲得,包括其全基因組DNA(cds)序列、蛋白質(zhì)序列(pep)以及其基因注釋文件(gff3)。
1.2? ?基因共線性推斷
首先,對大豆和野生大豆進行物種內(nèi)和物種間的Blast比對分析,用以確定基因同源性;然后利用共線性分析軟件ColinearScan鑒定基因是否是共線性基因,要求基因間距離大于50個基因、顯著性大于1e-10,以確定大豆與野生大豆基因組內(nèi)和基因組間同源基因間的共線性基因信息,然后以野生大豆全基因組基因作為參考,將其基因組內(nèi)的旁系同源基因和大豆之間的直系基因按親緣關系排列,生成共線性列表,并繪制全基因組聯(lián)合比對圖譜,目的在于將不同基因內(nèi)及基因組間全基因加倍事件后染色體重組、基因丟失、同源基因在染色體上的分布等現(xiàn)象進行直觀的展示。
1.3? ?基因置換推斷
由于大豆所經(jīng)歷的加倍事件較多,大豆與野生大豆分離在大豆獨有加倍事件之后,因此本研究只統(tǒng)計最近一次大豆獨有加倍事件后產(chǎn)生的大豆與野生大豆之間、基因組內(nèi)同源基因的差異。大豆中的一對旁系同源基因A1、A2以及它們所對應的野生大豆中的直系同源基因B1、B2構成同源基因“四聯(lián)子”。
根據(jù)系統(tǒng)發(fā)育關系,利用Clustalw[10]對同源基因四聯(lián)子中的基因進行多重序列比對,計算氨基酸一致性,同時分別計算兩兩之間的同義核苷酸置換率(Ks值),比較其Ks值大小,根據(jù)四聯(lián)子中基因的相似性和Ks值的變化判斷其是否發(fā)生基因置換。對發(fā)生基因置換的四聯(lián)子基因用MEGAx[11]進行系統(tǒng)發(fā)育構樹,對其系統(tǒng)發(fā)育樹的拓撲結構變化進行研究,從而達到置換檢驗的目的。
2? ?結果分析
2.1? ?大豆和野生大豆中同源性
以野生大豆全基因組為參考,構建與大豆間的共線性基因圖譜(圖1)。圈圖中的第一圈表示野生大豆的20條染色體,基于大豆與野生大豆共同經(jīng)歷的加倍事件,聯(lián)合圖譜展示了豆科和大豆獨有兩次加倍事件相關的同源基因的共線性關系。每一對同源基因間進行連線,不同染色體用不同顏色表示,內(nèi)圈表示最近一次加倍事件產(chǎn)生的同源基因,外圈為較遠的加倍事件產(chǎn)生的同源基因。由內(nèi)圈的圈圖可以看出,最近一次加
倍的野生大豆與大豆同源基因之間具有較完整的同源對應關系;越靠近外層的同源結構空白越多,表示同源基因丟失的越多,這是因為大豆經(jīng)歷的加倍事件次數(shù)較多,較為古老的加倍事件產(chǎn)生大量的重復基因隨著物種的進化而丟失。
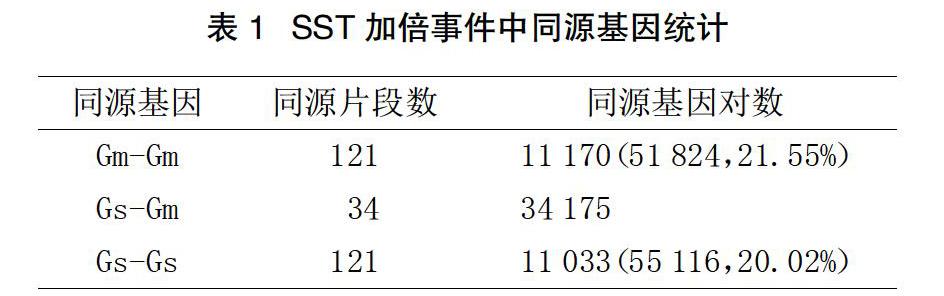
通過對共線性軟件ColinearScan分析獲得基因同源性片段和共線性列表的分析,分別統(tǒng)計了大豆與野生大豆基因組內(nèi)、基因組間與SST加倍事件相關的同源性片段的個數(shù)和所涉及的同源基因數(shù)(見表1)。對同源基因的統(tǒng)計發(fā)現(xiàn),大豆中保留的最近一次加倍事件相關的同源基因數(shù)量相對野生大豆更多,這可以說明大豆的基因保守性更好。
2.2? ?大豆與野生大豆的基因置換
分析大豆與野生大豆分歧后的基因置換規(guī)模,涉及大豆最近一次特異性全基因組加倍事件,根據(jù)共線性分析結果,得到17 306個最近一次加倍事件的同源基因四聯(lián)子,每個四聯(lián)子包括大豆和野生大豆的同源基因。根據(jù)同源基因間的親緣關系,大豆與野生大豆間由物種分化產(chǎn)生的旁系基因對的相似性應高于大豆與野生大豆基因組內(nèi)由加倍事件產(chǎn)生的直系基因對。但是由于基因置換,旁系基因對之間相似性變得更高。根據(jù)檢驗同源基因四聯(lián)子之間的拓撲結構,發(fā)現(xiàn)37對大豆同源基因和33對野生大豆同源基因之間發(fā)生了基因置換。因此,大豆中發(fā)生基因置換的基因數(shù)量多于野生大豆。
2.3? ?基因置換發(fā)生的位置分析
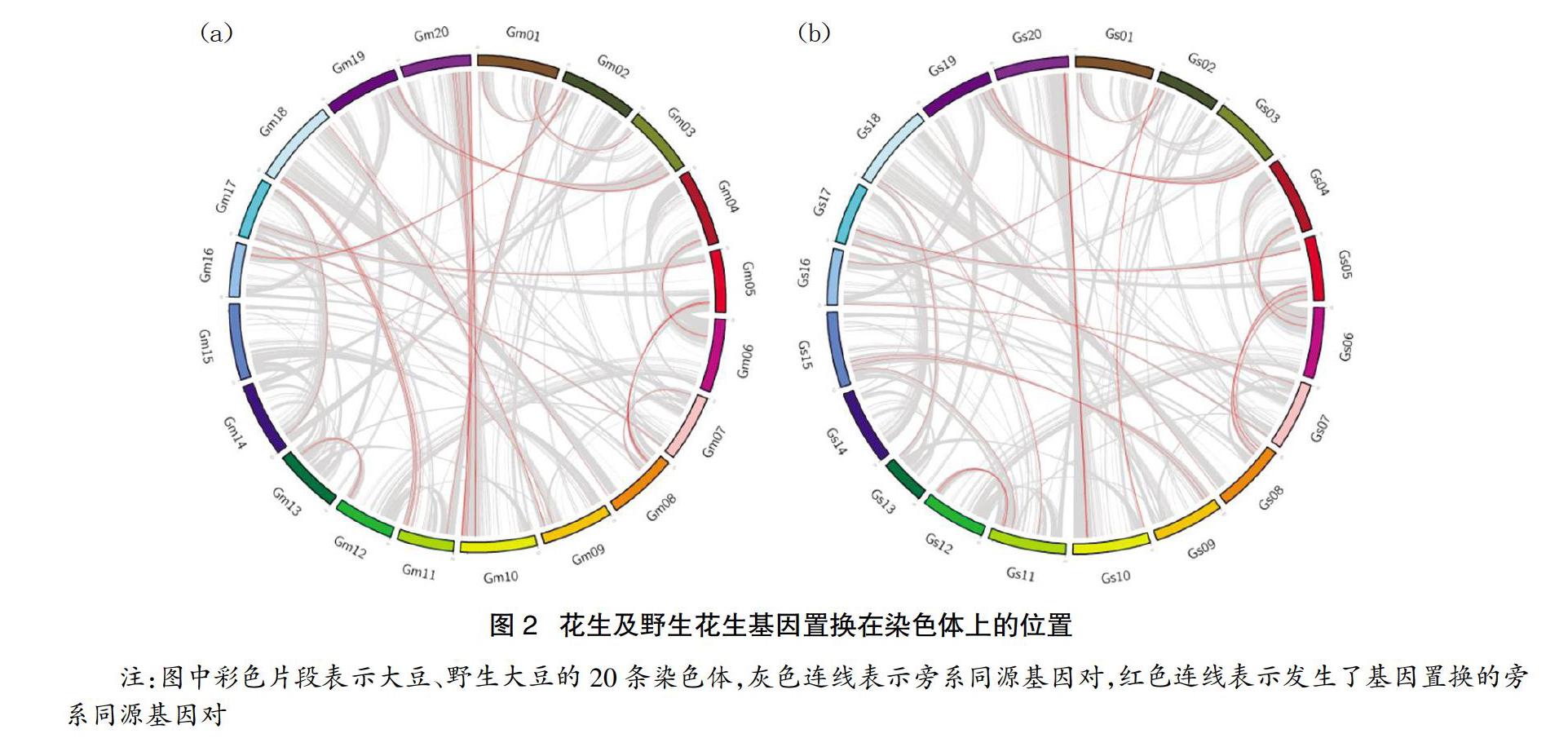
為了分析基因置換的發(fā)生與基因在染色體上物理位置的關系,分別將大豆與野生大豆發(fā)生基因置換的基因對應到其所在染色體的物理位置,并將結果繪制了圈圖進行展示(見圖2)。大豆發(fā)生基因置換的基因中有50%的基因位于靠近兩端10%的位置,86.46%的基因位于染色體兩端20%的位置,98.69%的基因位于靠近兩端30%的位置;野生大豆發(fā)生基因置換的基因中有48.48%的基因位于靠近兩端10%的位置,81.81%的基因位于染色體兩端20%的位置,98.48%的基因位于靠近兩端30%的位置。這可以表明,靠近染色體兩端的基因更容易發(fā)生基因置換。
3? ?討論與結論
植物基因組在進化過程中經(jīng)過一次或多次全基因組加倍事件[12- 13]。通過對禾本科物種研究發(fā)現(xiàn),基因置換等非正常遺傳重組的發(fā)生可以持續(xù)數(shù)千萬年,進而導致水稻、高粱的同源染色體之間形成一種特殊的結構,在高粱和水稻基因組中的很多基因可能由其同源基因置換而來。
研究在大豆與野生大豆分離后的同源基因中分別發(fā)現(xiàn)37對和33對同源基因發(fā)生了基因置換。對大豆與野生大豆發(fā)生基因置換的同源基因對在染色體上的位置統(tǒng)計發(fā)現(xiàn),有98%以上的基因置換發(fā)生在距離染色體兩端30%的位置,這一現(xiàn)象表明靠近染色體兩端的基因更容易發(fā)生基因置換現(xiàn)象,這與之前的研究結果相符合[14-15]。產(chǎn)生這一現(xiàn)象的原因是由于位于染色體兩端的基因往往保留有更好的共線性,保留的同源基因數(shù)量更多,因此這個區(qū)域發(fā)生基因置換的概率更大[16]。
對大豆與野生大豆分離后基因置換的統(tǒng)計結果顯示,在大豆與野生大豆分離后,由最后一次加倍事件形成同源四倍體基因組中的非正常遺傳重組等現(xiàn)象還在繼續(xù)發(fā)生。這一結果進一步表明在全基因組加倍事件后非正常遺傳重組等現(xiàn)象的影響非常持久[17-19]。
參考文獻:
[ 1 ] SHI X L,LING H Q.Current advances in genome sequencing of common wheat and its ancestral species[J].The Crop Journal,2018,6(1):15-21.
[ 2 ] YANTING, SHEN,JING,et al.De novo assembly of a Chinese soybean genome[J].Science China Life Sciences,2018,61(8):871-884.
[ 3 ] XIE M,CHUNG Y L,LI M W,et al.A reference-grade wild soybean genome[J].Nature Communications,2019,10(1):
85-96.
[ 4 ] XIAO G,WANG K,HUANG G,et al.Genome scale analysis of the cotton KCS gene family revealed a binary mode of action for gibberellin a regulated fiber growth [J].Journal of Integrative Plant Biology,2016,58(6):577-589.
[ 5 ] PATERSON A H,BOWERS J E,CHAPMAN B A.Ancient polyploidization predating divergence of the cereals,and its con-
sequences for comparative genomics[J].Proceedings of the National Academy of Sciences of the United States of America,2004,101(26): 9903-9908.
[ 6 ] JIA J,ZHAO S,KONG X,et al.Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation[J]. Nature,2013,49(7443):91-95.
[ 7 ] WANG X,TANG H,BOWERS J E,et al.Comparative inference of illegitimate recombination between rice and sorghum duplicated genes produced by polyploidization[J].Genome Research,2009,19(6):1026-1032.
[ 8 ] WANG X,TANG H,BOWERS J E,et al.Extensive concerted evolution of rice paralogs and the road to regaining independence [J].Genetics,2007,177(3):1753-1763.
[ 9 ] WANG X,SHI X,LI Z,et al. Statistical inference of chromosomal homology based on gene colinearity and applications to Arabidopsis and rice[J].BMC Bioinformatics,2006,7(1):447.
[ 10 ] LARKIN M A,BLACKSHIELDS G,BROWN N P,et al. Clustal W and Clustal X version 2.0 [J].Bioinformatics,2007,23(21):2947-2948.
[ 11 ] SUDHIR,KUMAR,GLEN,et al.MEGA7:Molecular evolutionary genetics analysis version 7.0 for bigger datasets[J]. Molecular Biology Evolution,2016,33(7):1870-1874.
[ 12 ] TANG H,BOWERS J E,WANG X,et al.Synteny and collinearity in plant genomes[J].Science,2008,320(5875):
486-488.
[ 13 ] SOLTIS D E,VISGER C J,MARCHANT D B,et al.Polyploidy:Pitfalls and paths to a paradigm[J].American Journal of Bot-
any,2016,103(7):1146-1166.
[ 14 ] WANG X Y,PATERSON A H. Gene conversion in angiosperm genomes with an emphasis on genes duplicated by polyploidization[J].Genes,2011,2(1):1-20.
[ 15 ] LIU C,WANG J,SUN P,et al.Illegitimate recombination between homeologous genes in wheat genome[J].Frontiers in Plant Science,2020(11):1076.
[ 16 ] CARVALHO,BERNARDO A.The advantages of recombination[J].Nature Genetics,2003,34(2):128.
[ 17 ] JACQUEMIN J,LAUDIé M,COOKE R.A recent duplication revisited: phylogenetic analysis reveals an ancestral duplication highly-conserved throughout the Oryza genus and beyond [J]. Bmc Plant Biology,2009,9(1):146.
[ 18 ] LING H Q,ZHAO S,LIU D,et al.Draft genome of the wheat A-genome progenitor Triticum urartu[J].Science Foundation in China,2013,496(7443):87-90.
(編輯:郝煥霞)
猜你喜歡
農(nóng)業(yè)科技通訊(2023年1期)2023-02-12 07:09:18
今日農(nóng)業(yè)(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農(nóng)業(yè)(2021年20期)2021-11-26 01:23:56
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農(nóng)產(chǎn)品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
