春小麥品系‘MY002894’和‘YJ006793’成株期抗條銹病基因遺傳分析
2021-11-12 20:19:44方世玉李秋榮侯璐等
植物保護 2021年4期
方世玉 李秋榮 侯璐等

中圖分類號:S 435. 121. 42 文獻標識碼:A DOI:10.16688/j.zwbh.2020169
小麥條銹病是由條形柄銹菌小麥專化型Puccinia strii jormis f.sp. tritici引起的氣傳病害,小麥整個生育期內都會發生。該病害具有分布范圍廣,影響面積大,危害嚴重等特點,對小麥的產量有著巨大影響。由于條銹菌新生理小種不斷產生和發展,現有的條銹抗源利用不夠合理,生產品種基因單一,使我國一些小麥品種‘喪失了對條銹菌的抗性[]。因此,從小麥材料中挖掘新的抗病基因,研究其遺傳特點,合理利用抗條銹病基因,是一項重要的基礎研究工作。
青海省是我國小麥條銹病的越夏流行地區之一。該省春小麥和冬小麥交錯種植,區域內種植‘阿勃‘京農411‘高原448等感病小麥品種,抗病春小麥品種種植較少。由于晚熟的春小麥上的菌源可以傳播到早播的冬麥苗上,導致省內條銹病流行十分頻繁,并且條銹菌可以通過大氣循環進行遠距離傳播,給我國東部冬小麥區發病提供較多的有效菌源口],對我國的條銹越夏菌源區治理產生不利影響口]。因此,尋找新的春小麥抗條銹病基因,研究其遺傳特點,進而培育出抗病新品種,對控制小麥條銹病流行十分重要。
為增加抗條銹病基因的多樣性,需要不斷尋找新的抗條銹病基因。但目前在春小麥抗條鎊病遺傳分析方面,國內研究較少。姚強等對春小麥品種‘青春39的抗條銹病能力研究結果表明,‘青春39'對CYR17的抗病性由1對顯性基因控制,而對CYR33的抗病性由1顯1隱兩對基因獨立控制。張調喜等對‘墨波/‘T29 'F2群體的抗性遺傳效應及青海春小麥品種‘青春38'成株期抗條銹性進行了遺傳解析。侯璐等先后對8個春小麥種質資源進行了苗期抗條銹病基因遺傳分析,明確了8份春小麥抗條銹病基因的遺傳特點,并且對青海春小麥品種‘高原363成株期抗條銹病基因進行了遺傳模型分析。
本研究組苗期分小種測試時發現,2個春麥品系‘MY002894和‘YJ006793表現為小種專化抗病性,對大田流行的最新小種CYR34表現感病,而2015年到2019年4年期間成株期大田測試均對條銹菌表現良好的抗病性。本研究擬分析這2個品系的成株期抗條銹病基因及遺傳特點,為后續合理有效的利用提供理論基礎。
1材料與方法
1.1試驗材料
‘MY002894'和‘YJ006793為抗條銹春小麥資源,綜合農藝性狀好,抗性強,由青海國家復份種質庫提供。感病對照春小麥品系‘Taichung 29'(T29)白中國農業科學院植物保護研究所徐世昌研究員提供。以‘MY002894'為父本與‘Taichung 29'(T29)雜交獲得F2:3群體,‘MY002894為父本與‘YJ006793雜交獲得的F2:3群體,由本研究室保存。
供試條銹菌小種Sunll-4、Sunll-6、CYR32、CYR33、CYR34由西北農林科技大學植物保護學院植物抗病遺傳研究室提供,青海省農林科學院植物保護研究所保存。
1.2試驗方法
春小麥品系‘MY002894'和‘YJ006793的苗期抗性鑒定于2015年在青海省西寧市青海省農林科學院植物保護研究所溫室進行,供試小麥材料分別播種在裝有泥炭土:蛭石:珍珠巖=2:1:1培養基質的7cm×7cm×7cm塑料花盆內,每盆種10~15粒,置于溫室按常規方法培養。待小麥幼苗第1片葉充分展開、第2片葉露尖時,采用涂抹法接種,將少許銹菌夏孢子放在潔凈的毛玻璃上,用滴管加入少量水,用接種針混勻涂抹到葉片上。幼苗放置于溫度為(10±1)℃的保濕箱內24 h后,移至溫室培養,溫度(16±1)℃,相對濕度80%,L∥D=16 h∥8h,光照強度9000 lx。待感病品系‘T29充分發病時(15~20d)調查反應型。
成株期抗性測試分別于2015年、2016年、2017年和2019年在青海省農林科學院植物保護研究所小麥條銹病自然發病圃進行。親本以撒播的方式種植1行,行長1m,行距0.3m,每行播10粒,2017年以單粒點播的方式種植F2群體單株,每行播10粒,2017年收獲的F。家系群體的種子于2019年繼續種植測試,每個F。家系撒播種植1行,每塊試驗地中每播種20行就插種1行‘Taichung29(T29)作為感病誘發對照組,鑒定圃的四周各種3行‘Taichung29(T29)作誘發行。播種時間為每年4月上旬,在每年7月上旬小麥抽穗期開始,待‘Taichung29(T29)充分發病時,對親本及F2群體(2017年)每個單株進行調查,對F3家系(2019年)每一行進行調查,記錄反應型和嚴重度,反應型和嚴重度調查重復2次,間隔7d,取2次調查數據的平均值進行最終統計。‘MY002894和‘Taichung29(T29)進行雜交后,于2017年收獲152個F2 群體單株,2019年收獲149個F3家系。‘MY002894和‘YJ006793進行雜交后,于2017年收獲176個F2群體單株,2019年收獲171個F3家系。
‘MY002894和‘YJ006793苗期和成株期反應型按Line等的0~9級方法記錄,0~5級為抗病;6~9級為感病,病害嚴重度(DS,%)按0~100%分級記錄。采用Excel2016軟件統計數據并繪制‘MY002894/‘Taichung29(T29)和‘MY002894/‘YJ006793群體的病害嚴重度和成株期反應型的頻率分布條形圖;統計各株系的抗感分離比,實際比值與期望比值的符合程度用χ2(≤χ20.05)進行適合性檢驗,確定最適合的分離比率。
2結果與分析
2.1供試親本‘MY002894' YJ006793和‘Tai-chung 29(T29)抗條銹性分析
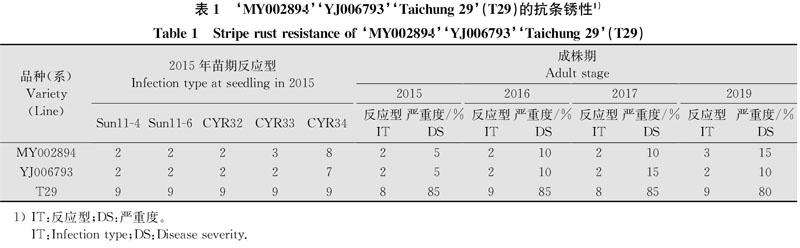
由表1可知,‘MY002894苗期對Sun11-4、Sun11-6、CYR32的反應型為2,對CYR33的反應型為3,表現為高抗:對CYR34的反應型為8,表現為高感。‘YJ006793苗期對Sun11-4、Sun11-6、CYR32、CYR33反應型為2,表現為近免疫;對CYR34的反應型為7,表現為感病。即‘MY002894和‘YJ006793苗期具有小種專化抗病性,但對近年流行的小種CYR34表現感病。通過對‘MY002894和‘YJ006793成株期病害的反應型和嚴重度的調查可以知道,2015年,2016年,2017年兩個品系的反應型為都為2,‘MY002894的病害嚴重度為5%、10%、10%,‘YJ006793的病害嚴重度為5%、10%、15%;2019年‘MY002894的反應型為3,病害嚴重度為15%;‘YJ006793的反應型為2,病害嚴重度為10%。‘MY002894和‘YJ006793苗期對近年流行的小種CYR34表現感病,但在大田成株期多年一直表現出很高的抗病性,這兩個抗性品系具有穩定的成株期抗病性。‘aichung 29(T29)苗期對上述5個小種均表現為高感;在田間的成株期調查中也表現為高感,說明‘aichung 29(T29)是一個感病品系。1) IT:反應型;DS:嚴重度。
IT:Infection type; DS: Disease severity
2.2‘MY002894抗條銹病基因遺傳分析
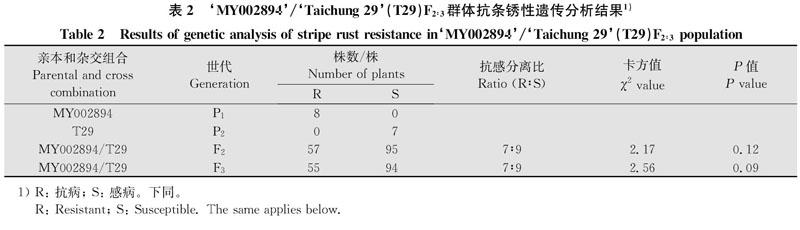
‘MY002894/‘T29F2:3群體單株(家系)的反應型和病害嚴重度數值分布圖如圖1,F2:3群體在2個環境下反應型和病害嚴重度中間型較多,反應型和病害嚴重度整體上均未呈現連續性分布,但是不同的區段內卻又出現了連續性分布的現象。對‘MY002894的抗條銹遺傳分析結果見表2,‘MY002894/‘Taichung29F2群體單株調查結果中,抗病植株為57株,感病植株為95株,即‘MY002894與‘Taichung 29雜交后代F2群體卡方測驗符合7R:9S的抗感分離比(X2=2.17,P=0.12)。對F3群體家系調查中,抗病家系有55個,感病家系有94個,也符合7R:9S的抗感分離比(X2=2. 56,P=0.09),由卡方分析結果得出‘MY002894有2對獨立作用的隱性成株期抗條銹病基因。
2.3‘YJ006793/‘MY002894F2:3群體抗條銹病基因遺傳分析
‘MY002894/‘YJ006793F2:3群體的反應型和病害嚴重度頻率分布見圖2,2017年F2群體和2019年的F3群體的反應型抗性較多,反應型和病害嚴重度整體均未呈連續分布,但是不同的區段內卻又出現了連續性分布的現象,兩年的測試結果呈現相同的變化趨勢。對‘MY002894/‘YJ006793F2:3群體的抗條銹遺傳分析結果見表3,在F2群體中,抗病植株為148株,感病植株為28株,‘MY002894和‘YJ006793雜交后代F2群體卡方測驗符合55R:9S的抗感分離比(X2=0.36,P=0.48)。對F3家系群體中,抗病家系有151株,感病家系有20株,‘MY002894與‘YJ006793F3群體感染條銹菌后卡方測驗也符合55R:9S的抗感分離比(X2=0.61,P=0.37)。即‘MY002894與‘YJ006793雜交F2:3群體對條銹菌的抗病性由1對顯性基因2對隱性基因獨立控制。由‘MY002894抗條銹病基因遺傳分析結果,‘MY002894中有2對獨立作用的隱性成株期抗條銹病基因,可推出‘YJ006793中有1對顯性成株期抗條銹病基因。
3討論
目前國內對小麥抗條銹病基因的遺傳分析主要是根據孟德爾經典遺傳定律進行分析,通過X2(≤X20.05)進行適合性檢驗,確定其最適合的分離比率,進而確定抗病基因的顯隱性、數目以及基因間作用方式,得出抗條銹病基因作用規律。張瑩等對小麥品系‘P9897成株期抗條銹性進行分析,發現其抗條銹性是由一顯一隱2對基因獨立控制或起重疊作用控制的。蘇萍萍等對小麥種質資源‘P10078成株期抗性進行研究,得出其由1對顯性主效基因控制。周春宏等通過研究西藏地方小麥品種‘曲白春后,發現其成株期對條銹菌的抗性由2對獨立顯性基因控制。馬東方等研究了‘小偃6號成株期對CYR30、CYR32的抗病性,得出‘小偃6號的抗病性均由2對隱性基因累加作用控制,對Su11-4的抗病性由1對顯性基因控制。邱亨池等發現‘秦農142成株期對條銹病的抗性由1對顯性基因和1對隱性基因共同決定。
本研究中,2個雜交群體成株期對條銹病的抗性由2對基因或3對基因控制。通過采用經典遺傳學的方法結合基因推導的方法分析‘MY002894和‘YJ006793的抗病遺傳規律,結果表明,‘MY002894成株期抗條銹性由2對隱性基因獨立控制,‘YJ006793中有1對顯性成株期抗條銹病基因。在4年的成株期抗性調查中發現‘MY002894'和‘YJ006793的抗病性比較穩定,對小麥條銹病有很高的抗性,對其抗性基因的研究有重要的意義。本研究通過抗感雜交確定‘MY002894中抗性基因的數目,依據抗感雜交的結果,再根據抗一抗雜交的結果,可以推測‘YJ006793中的抗條銹病基因類型。使用這樣的方法可以快速確定其抗條銹遺傳特點,直接篩選出比2個抗病親本表現更高抗病性和抗性持久性的家系,以期為這2個春小麥種質資源在抗病育種中的利用提供理論指導。‘MY002894和‘YJ006793都為抗性材料,抗一抗雜交可以實現抗病基因的累加,在后代中,有可能篩選出比2個抗病親本更加優良的抗條銹性和抗性持久性的家系,直接為育種家提供優良的抗性基因聚合材料和理論參考;也可以通過分子標記技術進行下一步研究,為春小麥抗病育種提供服務,從而獲得更有應用價值的抗病新種質或生產品種。
