
海參池中養殖銅藻的初步試驗
2021-11-19 12:01:28趙楠賽珊史良王玉本李霞
南方農業·中旬 2021年9期
趙楠 賽珊 史良 王玉本 李霞

摘 要 在海參養殖池中培養銅藻,探索銅藻在海參池中的適宜生長條件。試驗發現,銅藻移栽15 d后基本適應海參池的生存環境,在4月中旬至6月上旬,水溫在14~20 ℃,銅藻的生長狀態良好,增重也較快;到6月中下旬,水溫超過22 ℃,海參池中出現大量污損生物,如海鞘、紫貽貝等,嚴重影響銅藻的生長,此時銅藻不適宜在海參養殖池中生長。
關鍵詞 海參養殖池;銅藻;養殖
中圖分類號:S968.9 文獻標志碼:A DOI:10.19415/j.cnki.1673-890x.2021.26.009
銅藻(Sargassum horneri)屬馬尾藻科馬尾藻屬,在我國不連續分布于遼東半島、山東半島及福建省、廣東省等地,在俄羅斯、朝鮮、日本、越南也有分布。銅藻常見于近岸的淺海區,野生銅藻經常固著在海中的礁石或是纏繞在海帶養殖筏架上,少量銅藻殘枝因為外力從主枝上折斷脫落,也可單獨漂浮生存[1-2]。銅藻因其藻體細長,經常成片大量聚集,成為一些海洋生物的棲息地,因此銅藻也是構建海洋牧場的良好材料[3-5]。近年來,在北方養殖海帶,春夏兩季銅藻會大量暴發,纏繞在海帶養殖筏架上,使得筏架重量增加,浮漂無法承托重量,海帶也隨之下沉,無法受到足夠光照,容易受病,也給海帶收割帶來諸多不便。銅藻體內富含褐藻膠、褐藻多糖等物質,一些漁民會在閑時收集銅藻,曬干粉碎后作為鮑魚飼料,銅藻也是化工、醫藥、食品等產業的優質原料[6-10]。
研究表明,銅藻大量生長繁殖會對引起赤潮的藻類如甲藻等有抑制作用[11-12]。在北方海參養殖池里長期有多種雜藻,如棉花苔、滸苔等,尤其是在夏季,雜藻過度繁殖會消耗水中的大量氧氣,死亡腐爛更會導致水質的惡化,這些都會影響海參的生長發育。對此,在海參養殖池中試養銅藻,希望能夠通過銅藻的大量繁殖抑制雜藻,凈化水質,為海參的生長繁育起到一定的積極
作用。
1 材料與方法
1.1 試驗材料
試驗用銅藻于4月采集自山東省威海市的榮成東楮島海域。挑選纏繞在海帶養殖筏架上的銅藻,要求個體色澤好,生長狀態良好。將銅藻置于保溫箱中,保持濕潤,并用冰瓶保持低溫,確保銅藻的活性。
1.2 試驗地點
試驗地點為山東省煙臺市東方海洋高新技術開發區分公司所在地。試驗海參池面積約4.67 hm2,水深2.0~2.5 m,為砂質灘底。
1.3 試驗方法
在2 m的尼龍繩上,每間隔10 cm將1株銅藻穿過繩捻,多次纏繞,再用白色棉線繩固定在尼龍繩上,1條繩夾取15株銅藻,共計20繩。采用笩式養殖,平掛于海參養殖池中,每繩間距1 m,懸掛深度50 cm。分別在第0 d、第14 d、第33 d、第44 d、第68 d、第78 d對銅藻進行稱重觀測,選取其中3繩進行跟蹤記錄,編號為1號、2號、3號。
2 結果與分析
2.1 光照和溫度對銅藻生長的影響
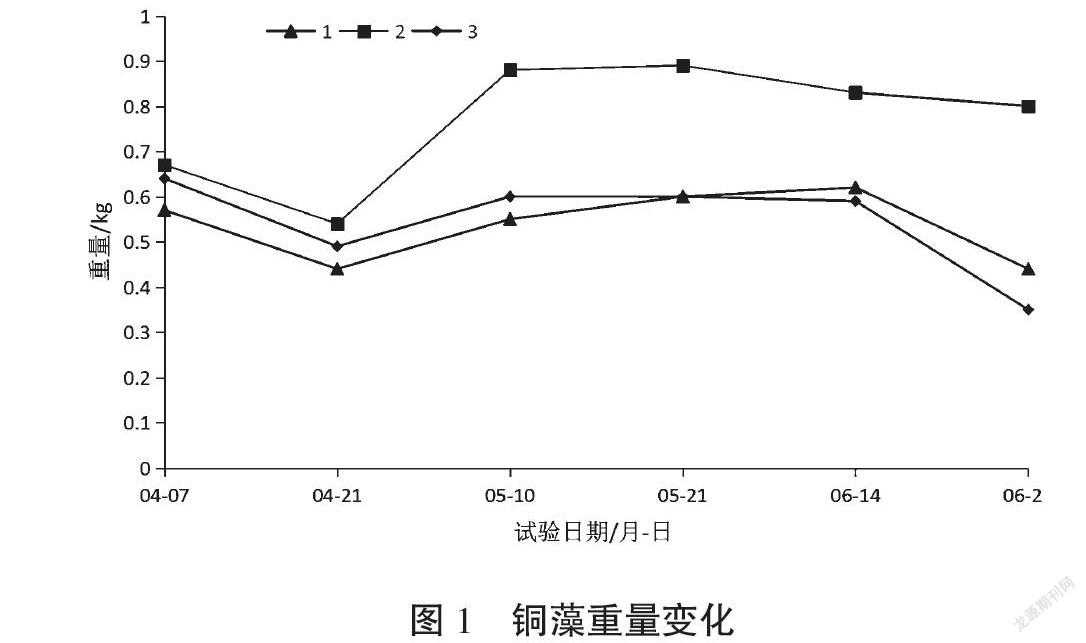
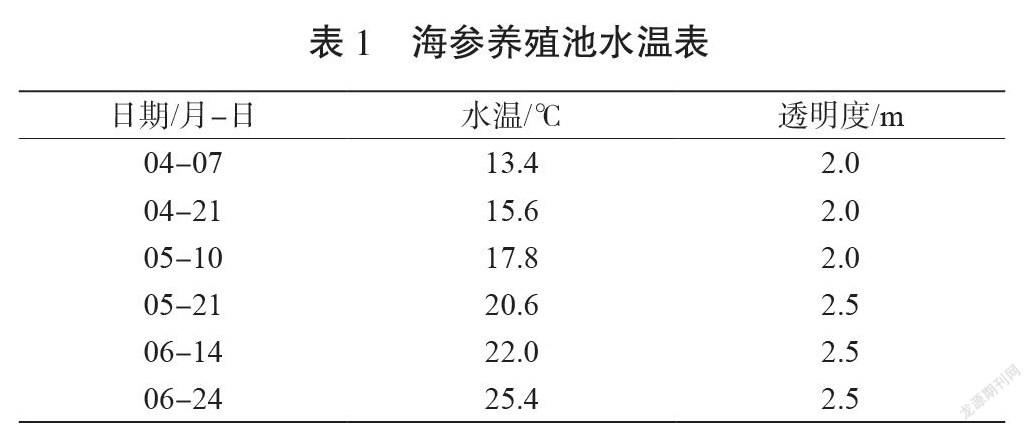
結合圖1、表1、圖2(a)可以看出,4月7日進行銅藻分苗移栽時,苗體較小,分枝較稀疏;移栽到海參池的前15 d,水溫低于15 ℃,3繩的銅藻重量均有所下降。4月20日至5月10日,水溫在15~20 ℃范圍內,銅藻進入生長期,生長繁茂增重較明顯,見圖2(b)。5月10日至6月14日,銅藻重量變化不明顯。6月14日后,水溫高于22 ℃,銅藻的藻體變短,部分養殖繩上的銅藻脫落并且長有污損生物,銅藻進入負增長期,不再適宜生存,3繩銅藻均負增重,見圖3(c)。3繩銅藻中,2號繩銅藻增重高于其他兩繩,并且在6月14日后銅藻減重亦不明顯。
通過表1可以發現,在整個培養周期中,海參養殖池的水體透明度高且基本不變,說明光照強度較高且變化不大。但是,銅藻除恢復期外,一直都處在較好的生長狀態,不但重量增加,還發育出了生殖托。這說明在海參養殖池中,光照強度不是限制銅藻生長的因素。在溫度方面,結合海參養殖池的實際情況,發現水溫14~20 ℃是適宜銅藻生長的溫度范圍。
2.2 溫度對銅藻生殖托形成的影響
在日常觀察期間發現,水溫16 ℃左右(4月下旬),少量藻體在一些側枝上出現了短棒狀的雌性生殖托和長棒狀的雄性生殖托。水溫在20 ℃左右(5月中旬)時,大部分銅藻都出現了生殖托。隨著夏季來臨,水溫的升高,發現生殖托數量逐漸減少。從中可以看出,銅藻生殖發育的時間在每年的4—5月,適宜發育水溫在16~20 ℃。
2.3 溫度對銅藻葉片形態的影響
銅藻藻體的側分枝由葉片、氣囊及生殖托構成。在水溫低于20 ℃(4—5月)時,銅藻側分枝的葉片生長在2個位置,一種為直接生長在氣囊頂部,另一種則直接生長在分枝上,此時的葉片呈披針狀或雙面鋸齒狀。到6月中旬時(水溫高于20 ℃),部分側枝的氣囊脫落,側枝的靠上部位則基本由葉片構成,葉片的形態類似于高等植物的羽狀復葉,在一棵葉片上,主枝水平向兩側分出小葉片。
3 結論與討論
3.1 環境因子對銅藻生長的影響
4月21日第一次觀測時,銅藻重量有明顯的下降,藻體顏色較上次變淺,呈黃色,葉片和氣囊有不同程度的腐爛,附有大量白色雜質,藻體在水中蕩洗后,白色雜質可以輕易脫落,這可能是投喂海參的海參餌料附著在藻體上形成的。大部分藻體梢部可見大量細小新生的分枝、葉片和氣囊,可見該部分銅藻已經適應新的環境,重新萌發新枝進入了生長狀態。
初次觀測銅藻重量下降的原因可能有以下2點。1)在夾苗階段,部分氣囊和葉片受損,從而導致部分主枝沉入水里,影響了其后期生長。2)正常的銅藻移栽至新環境后,對光照強度和水溫的變化需要重新適應和調整。
從實驗結果來看,4—5月,水溫14~20 ℃條件下銅藻進入了快速生長期,該實驗結果與孫建璋等的研究結果相同[12-14]。在這個時期內,隨著主枝增長變粗,顏色加深,其側分枝也越來越多,并且氣囊和葉片也隨之大量生長,最后變成長度長、分支繁茂的藻體,依靠大量的氣囊成片漂浮在水面。
3.2 污損生物對銅藻生長發育的影響
影響銅藻的生長發育的因素除溫度、光照等環境因子外,其他藻類及附著生物等對銅藻的生長發育也有顯著的影響。
到6月中旬的觀測時,海參養殖池的生物量激增,水面中漂浮著大量水母,而養殖閥架、養殖繩上及藻體上則附著大量的玻璃海鞘、紫貽貝及少量石灰蟲和苔蘚蟲。玻璃海鞘附著不牢固,可以通過抖動繩子和筏架輕易去除,其他種類的污損生物則較難去除。在6月下旬的觀察中,玻璃海鞘數量較上次變少,但紫貽貝和石灰蟲依舊大量附著在藻體上。在海水養殖中,玻璃海鞘、紫貽貝和石灰蟲是幾種具有明顯優勢的污損生物。在海帶養殖的后期,尤其天氣晴朗、風浪較小的時期,污損生物極易附著在海帶表面,使海帶腐爛變質,造成海帶的減產[15]。同樣,污損生物附著也會導致藻體腐爛,尤其對于枝條細長的銅藻,腐爛會使藻體更易脫落。伴隨著污損生物的大量出現,此時海參養殖池已經無法適宜銅藻的生存。
參考文獻:
[1] 曾呈奎.中國海藻志 第三卷 褐藻門[M].北京:科學出版社,2000:43-44.
[2] 王蕾.枸杞島海藻場生態系統與魚類群落季節變化的關系[D].上海:上海海洋大學,2011.
[3] 李美真,詹冬梅,丁剛,等.人工藻場的生態作用、研究現狀及可行性分析[J].漁業現代化,2007(1):20-22.
[4] 陳震,劉紅兵.馬尾藻的化學成分與生物活性研究進展[J].中國海洋藥物,2012(5):45-55.
[5] 方玉春.中國藥用馬尾藻資源調查及開發利用建議[J].中藥材,2013,36(8):1241-1244.
[6] 田鑫,李秀霞,吳科陽,等.海藻多糖提取純化及生物活性的研究進展[J].食品與發酵科技,2015,51(6):81-85.
[7] 劉振鋒,戴圣佳,呂衛金,等.褐藻多糖硫酸酯的生物活性與提取技術研究進展[J].食品與藥品,2015(1):71-75.
[8] 顧麗霞,劉麗佳,何淑婷,等.銅藻多糖水提法工藝優化及其抗氧化活性研究[J].安徽農業科學,2014(29):10139-10141.
[9] 劉婷.金潮原因種銅藻(Sargassum horneri)對關鍵環境因子的生理響應與機理解析[D].溫州:溫州大學,2019.
[10] 蔡佳宸.銅藻金潮對有害藻華原因種和浮游動物影響的模擬研究[D].北京:中國科學院大學,2019.
[11] 蔡佳宸,耿慧霞,孔凡洲,等.銅藻金潮對滸苔綠潮和幾種赤潮原因種生長影響的模擬研究[J].海洋與湖沼,2019,50(5):1050-1058.
[12] 孫建璋,莊定根,王鐵桿,等.南麂列島銅藻增殖技術的初步研究[J].現代漁業信息,2010,25(1):23-27.
[13] 孫建璋,莊定根,陳萬東,等.銅藻Sargassum horneri繁殖生物學及種苗培育研究[J].南方水產科學,2008,4(2):6-14.
[14] 孫建璋,莊定根,孫慶海,等.銅藻人工栽培的初步研究[J].南方水產科學,2009,5(6):41-46.
[15] 張明明,趙文,于世超.我國海洋污損生物的研究概況[J].水產科學,2008(10):46-50.
(責任編輯:趙中正)
