水杉原生種群胸徑樹高與樹冠的通徑分析1)
2021-12-03 07:13:22趙保國朱江艾訓儒姚蘭郭秋菊洪建峰
東北林業大學學報 2021年10期
趙保國 朱江 艾訓儒 姚蘭 郭秋菊 洪建峰
(湖北民族大學,恩施,445000) (湖北星斗山國家級自然保護區管理局)
水杉(Metasequoiaglyptostroboides)是我國特有的世界著名的杉科水杉屬孑遺植物,被譽為植物界的“活化石”,一度被認為已經全部滅絕,直到20世紀40年代在我國湖北利川謀道鎮被發現[1]。水杉的發現被公認為是我國甚至世界20世紀植物界的重大發現[2-3]。水杉從發現至今已有70多年,一直受到國內外學者們極大的重視,研究表明水杉原生種群呈衰退型[4-6],原生環境林下鮮有幼苗發現,存在天然更新困難[7-9],影響水杉種群天然更新的因素存在種源限制和種子擴散限制[9]。種子擴散過程除了受擴散方式的制約,還受母體特征的特性制約[10],而樹冠作為水杉球果的生長發育場所,冠幅大小和冠形等直接影響著水杉原生母樹的種子產量和林下植被的生活型,影響了種子的著地幾率、著地后的環境,與水杉的天然更新等存在密切聯系。樹冠作為林木的重要組成部分[11],是其與周圍生長環境相互作用的結果,是林木生理過程的主要場所,對林木的生長過程具有主導地位[12-13]。然而,現有的關于水杉原生種群的研究較少涉及樹冠層。
通徑分析是數量遺傳學家Sewall Wright[14]在1921年提出來的一種多元統計技術,是簡單相關分析的繼續,在多元回歸的基礎上將相關系數加以分解,通過直接通徑、間接通徑表示某一變量對因變量的直接作用效果、通過其他變量對因變量的間接作用效果[15]。胸徑、樹高、冠幅等是樹木的重要生理指標,有許多學者對柚木[16]、樟子松[17-18]、華北落葉松[19]、杉木[12,15,20]、禿杉[11]等對象開展了類似的研究。本文選取胸徑、樹高、冠幅等生理指標,利用通徑分析[15,21]研究水杉原生種群,明確水杉原生種群各影響因子與樹冠之間的關系,充分考慮各指標對水杉原生種群樹冠的直接影響和間接影響。研究表明,不同林齡的林分樹冠存在較大的差異[22]。目前,國內外尚未對水杉原生種群的樹冠特性進行詳細分析,本文以星斗山國家級自然保護區內的水杉原生種群為研究對象,分析和闡明水杉原生種群不同林齡個體的胸徑、樹高和樹冠之間的聯系,為進一步建立胸徑、樹高、樹冠的精準模型提供依據,為水杉原生種群的形態特征和個體生長發育的研究提供理論支撐,為水杉原生種群的保護和天然更新提供參考。
1 研究區概況
星斗山國家級自然保護區分為東、西兩個片區,區域內年平均氣溫12 ℃,年平均降水1 481 mm,四季分明,雨熱同期,為典型的亞熱帶大陸性季風氣候。水杉原生種群主要分布于保護區的西區(108°31′~108°48′E、30°4′~30°14′N),海拔范圍830~1 565 m,區域內主要為黃棕壤和黃壤,水資源充足,獨特的地理條件形成了水杉原生母樹的避難所。
2 研究方法
2.1 水杉原生種群個體性狀調查
2017—2018年間對保護區內全部水杉原生種群(5 663株)進行調查,使用胸徑尺測量胸徑,使用激光測距儀測量樹高和活枝下高,利用皮尺和激光測距儀測量冠幅長度(南北、東西)。
2.2 徑級與林齡劃分
通過徑級代替齡級[5,23]的方法,結合水杉徑級分布將水杉原生種群分為3個年齡階段,分別為中齡樹(小于100 a),成年樹(100~200 a)和老齡樹(大于200 a)。
2.3 樹冠因子的計算
冠幅(W):用東西向冠幅和南北向冠幅的平均值表示樹木的冠幅長度,其計算公式為:
W=(W1+W2)/2。
式中:W1、W2表示東西向與南北向的冠幅長度(m)。
冠長(L):指樹木頂部到第一個活枝的長度。其計算公式為:
L=H-H1。
式中:H為樹高(m);H1為樹干第一個活枝高度(m)。
冠長率(R):指樹冠長度在樹干中所占的比率。具體公式為:
R=L/H。
冠形率(S):冠形率用冠長(L)與冠幅(W)的比值表示:
S=L/W。
以3 m為級距對冠幅進行分級,18 m為分級上界,共分為7級,從而使不同林齡的水杉原生母樹冠幅占比更好體現,同理,采用類似冠幅分級的方式將冠長率以0.1為級距分為10級。
參照杜家菊[24]、宋小園[25]等,使用SPSS對數據進行處理分析,根據通徑分析理論和計算公式,得出直接通徑系數、間接通徑系數和決策系數。決策系數是通徑分析中的決策指標,用它可以把自變量對響應變量的綜合作用進行排序,確定主要決策變量和限制變量[26]。其計算公式具體為:

3 結果與分析
3.1 胸徑和樹高結構
水杉原生種群不同林齡階段的胸徑、樹高的統計信息見下表。由表1、表2可知,星斗山自然保護區水杉原生種群中,胸徑變化在24.2~248.0 cm,中齡樹、成年樹、老齡樹的平均胸徑依次為60.8、72.9、98.1 cm,隨著林分年齡增大,胸徑增大的趨勢也在發生變化,其中,平均胸徑在不同年齡階段的增長率依次為19.90%、34.60%。通過表3可知,水杉原生母樹的樹高主要分布在9~51 m,中齡林、成年林、老齡林的平均樹高為25.4、29.3、36.0 m,樹高在不同年齡階段的增長率為15.35%、22.80%,但最大值出現在成年樹。通過差異性分析表明,不同林齡水杉原生母樹的平均胸徑與平均樹高均具有顯著性差異(p<0.01)。

表1 水杉原生種群胸徑分布

表2 水杉原生種群不同林齡胸徑統計
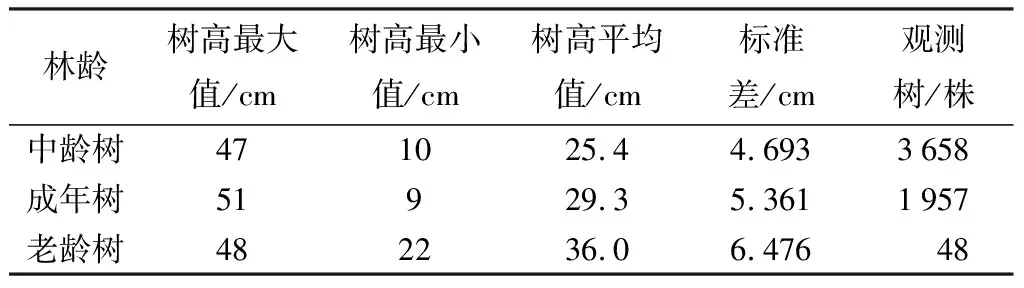
表3 水杉原生種群不同林齡樹高統計
3.2 樹冠結構
通過表4、表5可知,平均冠長和冠幅都在隨著水杉年齡的增加而增大,中齡林、成年林、老齡林的平均冠長依次為21.2、24.6、30.2 m,水杉冠長在各個階段的增長率分別為16.04%、22.93%。水杉原生母樹的平均冠幅在各個階段依次為7.7、8.9、10.6 m,冠幅的變化率依次為15.58%、19.09%,不同林齡的水杉冠幅和冠長都具有顯著差異(p<0.01)。

表4 水杉原生種群不同林齡冠長統計

表5 水杉原生種群不同林齡冠幅統計
由表6可知,中齡樹的冠幅主要分布在3~9 m,成年樹和老齡樹的冠幅主要分布在6~12 m,冠幅大于18 m的母樹有且僅有1株,冠幅小于3 m的母樹僅占中齡樹的2.73%,成年樹的1.43%。隨著年齡增長,3~6 m的冠幅占比大幅度減少,6~9 m的冠幅占比減勢微小,幾乎持平,9~12 m與12~15 m兩級別的冠幅占比呈顯著增加。水杉原生種群整體的集中區域從6~9 m往9~12 m偏移。

表6 水杉原生種群不同林齡冠幅占比
3.3 冠長率結構
由表7可知,整個原生種群冠長率分布區間為0.148~0.970,中齡樹到老齡樹變異系數成先減小后增大趨勢,說明成年樹的冠長率波動較另外兩齡級小一些,整體的冠長率呈遞增趨勢。

表7 水杉原生種群不同林齡冠長率統計
通過表8可知,中齡樹的冠長率主要聚集于0.8~0.9,中齡樹的冠長率主要集中于0.8~1.0階段,老齡樹主要分布在0.9~1.0區間,整個水杉原生母樹種群冠長率在0~0.1沒有分布,隨著年齡的增加,0.1~0.3區間的冠長率占比逐漸減少,0.5~0.6、0.7~0.8區間的冠長率占比幾乎不變,0.8~0.9區間的冠長率占比呈下降趨勢,0.9~1.0區間的冠長率所占比例增加。

表8 水杉原生種群不同林齡冠長率占比
3.4 胸徑、樹高與樹冠因子的通徑關系
3.4.1 胸徑、樹高與冠幅的通徑分析
水杉原生種群的胸徑、樹高和冠幅之間的關系如表9所示。胸徑與樹高對冠幅的決策系數在不同林齡間都為正數,說明胸徑與樹高對冠幅都為促進作用。樹高與冠幅之間的直接通徑系數呈先增大后減小的趨勢,且樹高對冠幅的正向作用在成年林時最大。胸徑對冠幅的正向作用在老齡林中表現顯著,不同年齡階段胸徑對冠幅的直接作用均大于間接作用,通過分析直接通徑系數和決策系數可知,胸徑對于冠幅的影響均大于樹高對冠幅的影響。

表9 水杉原生種群不同林齡胸徑、樹高與冠幅的通徑分析
3.4.2 胸徑、樹高與冠長率的通徑分析
由表10可知,水杉原生母樹在不同林齡階段樹高對冠長率的直接通徑系數均大于0,說明樹高對冠長率均為正向作用,且在成年林時作用最大。胸徑與冠長率之間的直接通徑系數整體處于下降的趨勢,在成年林達到最小值,表明胸徑對于冠高率的正向作用在逐漸減小并一度出現負向作用。

表10 水杉原生種群不同林齡胸徑、樹高與冠長率的通徑分析
3.4.3 胸徑樹高與冠形率的通徑分析
不同林齡水杉原生母樹胸徑樹高和冠形率之間通徑分析結果如表11。樹高對冠形率的間接作用一直小于直接作用。水杉原生母樹處于中齡林時冠形率與胸徑和樹高之間的直接通徑系數均為最大,表明在這個階段胸徑和樹高對冠形率有較大的正向作用。樹高對冠形率的直接通徑系數在成年樹達到峰值,整體趨勢為呈現先減后增,說明樹高對水杉原生母樹冠形率的正向作用力先減小后增大。胸徑對于冠形率的決策系數呈遞減趨勢且變為負值,說明胸徑對于冠形率正向作用一直減小甚至成為負向作用。
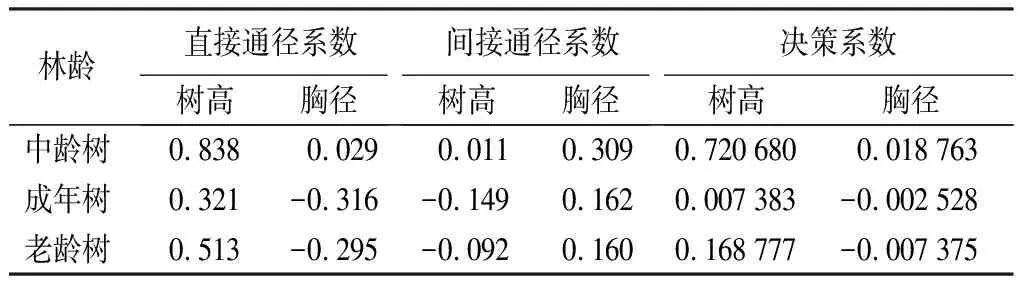
表11 水杉原生種群不同林齡胸徑、樹高與冠形率的通徑分析
4 討論
4.1 水杉原生種群樹冠特征
隨著林齡的增加,主要分布冠幅由3~9 m到9~15 m過渡,高級別的冠長率所占比例增加。最大冠幅22 m為謀道1號母樹,僅為個例,整個水杉原生種群冠長率在0~0.1沒有分布,但0.1~0.3有極少個體分布,這部分個體多為受雷擊個體,對比熊彪[4]和黃小[27]對水杉原生種群的研究結果,近二十幾年死亡個體中雷擊導致的死亡樹木約占31.3%,其受雷擊影響的個體約占整體1.61%。水杉原生種群中現因雷擊和斷梢造成個體瀕危所占比例均為21.31%,生長狀況衰弱的個體中雷擊和斷梢所占比例為16.49%、30.05%。雷擊與斷梢降低了樹高和冠長,嚴重影響冠長率,而斷梢個體大部分原因也是雷擊,所以對水杉原生種群產生主要影響的是雷擊。對水杉原生種群的后續保護過程中,可以為較高處于空曠地帶的水杉原生母樹人為安置避雷設施,例如避雷針等,通過避雷措施使其避免雷害,提高存活率。
通過實地調查結合林勇[5]和Tang et al.[28]對水杉種群的研究,50.89%的水杉個體分布在居民房前屋后,由于水杉個體地處農田、距離房屋太近、前期水杉種子被當地交易當做經濟收入來源等原因,水杉個體被人為剃枝,對冠幅寬度、第一活枝高等產生直接影響。部分水杉個體剃枝嚴重導致生長狀況呈瀕危狀,但僅為個例,對整個水杉原生種群而言影響較小。對水杉原生種群的后續保護過程中,應加強當地居民保護意識,進一步避免人為因素直接導致水杉個體死亡,影響水杉種群樹冠結構。
4.2 胸徑、樹高與樹冠因子的關系
通徑分析[29]中的直接作用和間接作用能夠更加精確地顯示各因素的相關關系大小和相對重要性。胸徑與樹高對冠幅的決策系數在不同林齡間都為正數,說明胸徑與樹高對冠幅都為促進作用,這與盧妮妮[15]所調查的杉木情況類似。在水杉原生種群不同林齡間,胸徑對冠幅的作用呈先減小后增大的趨勢,成年樹階段胸徑對冠幅作用變小,導致原因可能是受人為影響,早期當地居民人為剃枝采種導致。中齡樹和成年樹階段,對冠長率的影響排序為樹高大于胸徑,在老齡樹階段,對冠長率的影響胸徑大于樹高,這除了與水杉本身性質有關,還可能是由于部分老齡樹遭受雷擊斷梢導致,所以日后的水杉母樹保護中,避免老齡樹遭受雷擊是重中之重。
樹高對冠長率、冠形率均為正向作用,胸徑對冠形率冠長率的作用整體呈下降趨勢甚至表現為抑制作用。通過比較杉木[15]和水杉,樹高對冠長率作用變化趨勢相同,成年林階段胸徑對冠長率的間接作用大于直接作用,胸徑對冠長率作用在成年林階段表現為負作用可能是受人為因素影響。通過比較不同林齡間胸徑和樹高對各冠幅因子的直接作用和間接作用,可以提高水杉母樹胸徑、樹高、冠幅數學模型的精確性,為進一步建立數學模型提供依據。
5 結論
整體來說,水杉原生種群平均冠長和冠幅都在隨年齡的變大而增加,且增加趨勢近乎一樣,冠形率和冠長率都由低級別向高級別過渡,雷擊是影響水杉原生種群樹冠特征的主要因素。不同林齡階段的胸徑對冠幅的直接作用大于間接作用,它的影響程度大于樹高,這與冠形率、冠長率相反。在中齡林和老齡林階段,胸徑對冠長率呈正向作用,整個生長階段胸徑對于冠長率、冠形率之間的直接通徑系數整體處于下降的趨勢,甚至成熟林均表現為負作用。胸徑和樹高對冠形率影響在中齡林均為正向作用,在成年林和老齡林,胸徑對冠形率呈現顯著負向作用,不過胸徑對老齡林冠形率的抑制作用小于成年林。通過分析了解到水杉原生種群胸徑、樹高對冠幅等因素的作用大小,可以結合水杉原生種群生長特性,進一步建立精確的不同林齡階段的冠幅預測模型,進一步探討胸徑、樹高等與天然更新之間的聯系,為水杉原生種群的管理和保護提供參考和依據。
