產幾丁質酶菌株GXUN-20的篩選、鑒定及其產酶條件優化
2021-12-16 05:39:02王一兵申乃坤姜明國
食品工業科技 2021年24期
關鍵詞:生長
張 奇,王一兵,2, ,申乃坤,姜明國
(1.廣西民族大學海洋與生物技術學院, 廣西多糖材料與改性重點實驗室, 廣西海洋微生物資源產業化工程技術研究中心, 廣西高校微生物與植物資源利用重點實驗室, 廣西南寧 530008;2.廣西科學院, 廣西海洋天然產物與組合生物合成化學重點實驗室, 廣西南寧 530007)
幾丁質(Chitin)又名甲殼素、甲殼質,是由N-乙酰-D-葡糖糖胺以β-1,4糖苷鍵連接而成的天然多聚體,其含量在自然界僅次于纖維素,廣泛存在于海產品蝦蟹的外殼、節肢動物、大多數昆蟲的外骨骼,以及真菌和藻類等植物的細胞壁中[1]。近年來,海鮮的生產和消費不斷增加,蝦蟹殼是目前主要的海鮮生產廢棄物[2],由于技術和市場的限制,海鮮廢棄物的再利用受到了限制,其中有部分蝦蟹殼廢棄物用于堆肥或轉化為低附加值產品,比如動物飼料和化肥,大部分廢棄物在沒有處理的情況下直接丟棄到環境中,如果處理不當,會對環境造成嚴重的危害[3]。然而,這些蝦蟹殼廢棄物是具有高效益原材料的來源,可用于生產生物產品[4]。
蝦蟹殼中的幾丁質不溶于水、稀酸、稀堿等常用試劑,嚴重限制了其商業應用,幾丁質經幾丁質酶(Chitinase)水解得到易溶于酸的殼聚糖[5];殼聚糖是自然界中發現的唯一堿性多糖,因其分子中有大量的游離氨基,所以溶解性能比幾丁質有所改善,并具有生物可降解性,被廣泛應用于食品、醫藥、化妝品、環保、農業等領域,具有很大的生物技術應用價值和商業前景[6]。目前,生產殼聚糖主要采用化學方法,但此方法耗能大,較危險,且容易造成環境污染等問題[7],酶解法生產殼聚糖綠色環保,反應條件較溫和,耗材耗能相對較低,并且可以獲得脫乙酰程度較均一穩定的產品,運用幾丁質酶生產殼聚糖是近年來研究的熱點。因此,幾丁質酶研究的重點在于篩選出產酶量高、適合大規模化生產的菌株,探索適合的發酵工藝等方面[8]。
海岸灘涂具有獨特的微生物環境,蘊含著豐富的具有催化特性酶類的微生物資源[6],其沉積物中含有大量蝦蟹及貝殼廢棄物,蝦蟹殼含有豐富的蛋白質和幾丁質,是碳和氮的重要來源[9],尋找具有催化特性的幾丁質酶類,不僅對降解蝦蟹殼廢棄物以提高其利用率、保護環境具有很好的指導作用[10],而且蝦蟹殼降解后得到的豐富的碳氮源還可以減少農藥和化肥的投入使用,能夠改善土壤肥力、增強土壤微生物菌群活力,為菌肥的研發提供了新思路[11]。因此,本研究從廣西合浦縣大神木海鴨場附近灘涂沉積物中篩選產幾丁質酶細菌,并對篩選得到的菌株進行16S rDNA屬種鑒定,對其中一株產幾丁質酶活力較強菌株進行產酶條件優化研究,提高發酵產酶水平,以期為降解蝦蟹殼酶法生產殼聚糖提供實驗基礎。
1 材料與方法
1.1 材料與儀器
灘涂沉積物 中國廣西合浦縣大神木海鴨場,東經 108°54′25″,北緯 21°37′25″,置于-15 ℃ 左右車載冰箱立即帶回實驗室進行菌株篩選;細菌病原菌:金黃色葡萄球菌(Staphylococcus aureus1.10755)、大腸桿菌(Escherichia coli1.12883)、枯草芽孢桿菌(Bacillus subtilis1.15792);真菌病原菌:黑曲霉(Aspergillus niger3.3928)、尖孢鐮刀菌(Fusarium oxysporum3.18025)、立枯絲核菌(Rhizoctonia solani3.17046)、白色鏈霉菌(Streptomyces albus4.7658)
以上菌種均購于中國普通微生物菌種保藏管理中心(CGMCC);幾丁質 生工生物工程(上海)股份有限公司;PCRMix 北京康潤誠業生物科技有限公司;引物 北京擎科新業生物技術有限公司;其它試劑 國產分析純試劑。
GI54DS高壓蒸汽滅菌鍋 致微(廈門)儀器有限公司;SW-CJ-1F單人雙面垂直凈化工作臺 上海汗諾儀器有限公司;ZXMP-A1230電熱恒溫培養箱、搖床 上海智城分析儀器制造有限公司;HH-2數顯恒溫水浴鍋 常州國華儀器有限公司;LEGEND MICR 017微量臺式離心機、Multiskan Go 全波長酶標儀 美國Thermo Scientific公司;BioRad T100 Mastercycler nexus聚合酶鏈式反應(PCR)儀 Bio-Rad公司。
1.2 實驗方法
1.2.1 培養基 DNS溶液、膠體幾丁質溶液的配制:根據傅麗君等[12]的方法進行DNS和膠體幾丁質溶液的配制;LB 培養基(g/L):NaCl 5.0 g,酵母膏 5.0 g,胰蛋白胨 10.0 g,pH 7.0~8.0[13](若為固體,加入瓊脂15.0 g);PDA 培養基(g/L):馬鈴薯 200.0 g,葡萄糖20.0 g,瓊脂 15.0 g,pH 自然。
復合鹽母溶液(g/L):K2HPO470.0 g,KH2PO430.0 g,MgSO4·7H2O 50.0 g,FeSO4·7H2O 1.0 g[14]。
液體富集培養基(g/L):幾丁質粉末 8.0 g,(NH4)2SO42.0 g,復合鹽母溶液 10 mL,pH7.2~7.5。
膠體幾丁質培養基(g/L):膠體幾丁質4.0 g,(NH4)2SO42.0 g,復合鹽母溶液10 mL,瓊脂15.0 g,pH7.0。
種子培養基(g/L):幾丁質粉末10.0 g,酵母粉5.0 g,蛋白胨5.0 g,復合鹽母溶液10 mL,pH7.0。
發酵培養基(g/L):幾丁質粉末10.0 g,(NH4)2SO42.0 g,復合鹽母溶液 10 mL,pH7.0。
1.2.2 產幾丁質酶菌株的篩選及發酵酶活測定 稱取灘涂沉積物樣品5.0 g,加入25 mL無菌水中,混勻后于30 ℃、180 r/min 培養4 h,取1 mL上清液加入到29 mL液體富集培養基中,混勻后于30 ℃、200 r/min 培養 3~4 d后,取樣液梯度稀釋至 10-3、10-4、10-5并分別吸取200 μL于膠體幾丁質培養基上涂布分離,30 ℃倒置培養2~5 d[15-16],觀察菌落生長及透明圈產生情況,挑取產透明圈的菌株在LB固體培養基中純化出單菌落[17],同時接種于LB斜面培養基-4 ℃低溫保藏及30%甘油管-80 ℃超低溫保藏。
挑選單菌落至種子培養基中,于30 ℃、200 r/min培養2~3 d后,再以2%(v/v)接種量接種至發酵培養基30 ℃、200 r/min培養4 d后,發酵液8000 r/min離心10 min取上清粗酶液。取1 mL膠體幾丁質溶液(1%)于5 mL離心管中,45 ℃水浴預熱10 min之后加入1 mL稀釋粗酶液。45 ℃反應1 h后,混勻取出200 μL加入300 μL DNS終止反應,在沸水中顯色10 min,冷卻后加入300 μL去離子水,12000 r/min離心5 min,取上清液于540 nm處測吸光值,以煮沸滅活的上清液作為對照。酶活力單位(U)定義為:在45 ℃下每分鐘產生1 μmol N-乙酰-D-氨基葡萄糖所需要的酶量[8]。
1.2.3 菌株GXUN-20 16S rDNA擴增及分析 煮沸法提取DNA模板:挑取單一菌落至EP管中,加入100 μL 20%的Chelex裂解液,振蕩煮沸30 min后,12000 r/min離心10 min,取上清。PCR引物:27F:5'-AGAGTTTGATCCTGGCTCAG-3';1492R:5'-GG TTACCTTGTTACGACTT-3'。反應體系 50 μL:PCR Mix 23 μL,ddH2O 21 μL,上下游引物各 2 μL,模板2 μL。反應程序:98 ℃ 預變性 5 min;95 ℃ 變性1 min,55 ℃ 退火 1 min,72 ℃ 延伸 90 s,34 個循環;72 ℃終延伸10 min,4 ℃保存。PCR擴增產物送至生工生物工程(上海)股份有限公司測序,所得序列提交GenBank,并與GenBank數據庫中的序列進行同源性比對,用MEGA 6.0軟件進行16S rDNA序列的比對分析,并構建系統發育樹。
1.2.4 菌株GXUN-20的鑒定以及抑菌實驗 產透明圈最大的菌株命名為GXUN-20,根據伯杰氏細菌鑒定手冊對菌株GXUN-20進行形態觀察和生理生化鑒定[18]。抑菌實驗采取濾紙法[19],將細菌病原菌分別接入 LB培養基,37 ℃培養 12 h,吸取 50 μL至LB固體培養基涂布,在濾紙上滴加GXUN-20菌液8 μL后放置在LB培養基上,30 ℃倒置培養1 d;真菌病原菌分別接種至PDA培養基中央,在濾紙上滴加GXUN-20菌液8 μL后放置于真菌的對立面,25 ℃倒置培養3~5 d,分別觀察濾紙片四周是否有抑菌圈生成。
1.2.5 單因素實驗 優化前發酵條件:發酵培養基,30 mL/150 mL 裝液量,2%(v/v)接種量,30 ℃、200 r/min發酵培養4 d,進行酶活測定。
1.2.5.1 生長曲線及最適發酵產酶時間 分別在發酵的第 1、2、3、4、5、6、7、8 d檢測 OD600及酶活。
1.2.5.2 碳源濃度的確定 將種子液接種到幾丁質粉末添加量分別為 6.67、10.0、13.33、16.67、20 g/L的培養基中,其它發酵因素不變,恒溫培養測定酶活。
1.2.5.3 氮源及其添加量的確定 將種子液接種到分別以2 g/L的硫酸銨、蛋白胨、氯化銨、黃豆餅粉、尿素、牛肉膏為唯一氮源培養基中,在最適氮源基礎上進行最適氮源添加量(1、2、3、4、5 g/L)優化,其它發酵因素不變,恒溫培養測定酶活。
1.2.5.4 發酵培養溫度的確定 將種子液接種至發酵培養基后,置于溫度分別為 26、28、30、32、35、37 ℃搖床中,其它發酵因素不變,培養測定酶活。
1.2.5.5 發酵液初始pH 將種子液接種到發酵液初始 pH分別為 7.8、8.0、8.2、8.4、8.6、8.8、9.0的培養基中,其它發酵因素不變,恒溫培養測定酶活。
1.2.5.6 接種量的確定 將種子液以接種量分別為0.5%、1%、2%、3%、4%、5%(v/v)接種于培養基中,其它發酵因素不變,恒溫培養測定酶活。
1.2.6 正交試驗 根據單因素實驗結果選出對產幾丁質酶影響較大的4個因素(氮源濃度、發酵液初始pH、碳源濃度、接種量)進行4水平正交試驗(表1)。單因素溫度優化結果比幾丁質粉末濃度優化結果酶活力高,但考慮到該菌株是以幾丁質為底物發酵產幾丁質酶,固選擇幾丁質粉末添加量為4因素之一,以幾丁質酶活力為考察指標,探究該菌株最佳產酶條件。
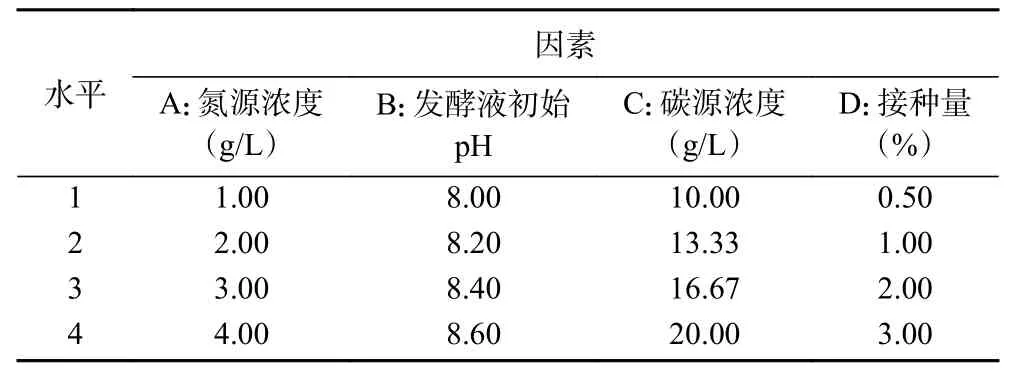
表1 4因素4水平正交實驗Table 1 Orthogonal experiment of 4 factors and 4 levels
1.3 數據處理
采用GraphPad Prism 8作圖,采用SPSS21.0進行正交實驗數據統計分析,實驗均為三組平行。
2 結果與分析
2.1 產幾丁質酶菌株的篩選及發酵酶活測定結果
將富集培養的灘涂沉積物稀釋液涂布于膠體幾丁質固體培養基,30 ℃倒置培養3 d,菌株在膠體幾丁質固體培養基上生長,并產生白色透明圈,結果如圖1所示,挑選有透明圈的菌株純化至單菌落,共得到3株產丁質酶菌株,將透明圈最大的菌株命名為GXUN-20。以N-乙酰-D-氨基葡萄糖標準品繪制標準曲線,R2=0.9971,結果如圖2所示,按照酶活測定方法,測定該菌株未優化之前幾丁質酶的酶活為0.140 U/mL。
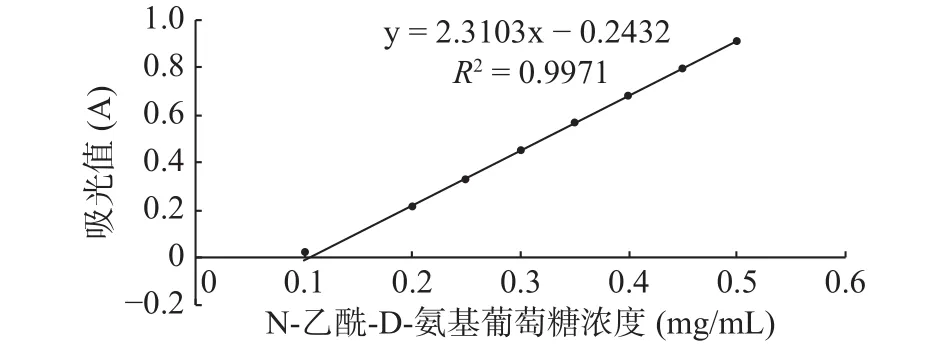
圖2 N-乙酰-D-氨基葡萄糖標準曲線Fig.2 Standard curve of N-acetyl-D-glucosamine
2.2 菌株GXUN-20 16S rDNA擴增及分析
將PCR產物送至生工生物(上海)股份有限公司,所得 1411 bp序列提交 GenBank檢索,并與GenBank數據庫里的序列進行同源比對,發現與Lysobacter firmicutimachus(登錄號:KU593484)菌株16S rDNA相似度最高為99.79%。運用MEGA 6.0軟件進行16S rDNA序列的對比分析,并構建系統發育樹如圖3,從圖中可以看出 GXUN-20與Lysobacter firmicutimachus菌株親緣關系最接近,可初步鑒定GXUN-20(登錄號:MW897735)為厚壁溶桿菌。
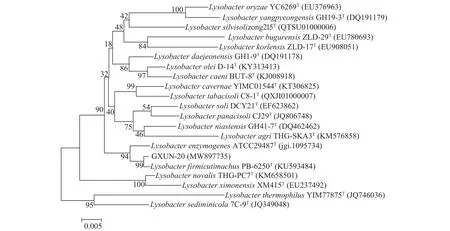
圖3 基于16S rDNA序列構建菌株GXUN-20系統發育樹Fig.3 Phylogenetic tree of strain GXUN-20 based on 16S rDNA sequence
2.3 菌株GXUN-20的鑒定以及抑菌實驗結果
2.3.1 菌株GXUN-20形態學鑒定 菌株GXUN-20的形態學觀察見圖4。菌株GXUN-20為革蘭氏陰性短桿菌,無芽孢,菌落呈淡黃色圓形,表面濕潤光滑,邊緣規則,無暈環,中央飽滿突起,直徑0.1~0.5 mm,挑起有粘性,較易挑取,與Miess等[20]研究結果相似,在有氧條件下生長于LB、R2A、TSA等培養基,菌落圓形凸出,奶油黃色,圓端非孢子棒狀,單個或成對出現,無氧條件下不生長。

圖4 菌株GXUN-20的形態觀察結果Fig.4 Morphological observation results of strain GXUN-20
2.3.2 菌株GXUN-20生理生化特征 菌株GXUN-20部分生理生化試驗結果及說明見表2。從結果得知,明膠液化試驗、過氧化氫試驗、SIM生化試驗、吲哚試驗、七葉苷試驗均呈現陽性,VP、MR、硝酸鹽試驗、甘露糖、麥芽糖、葡糖糖、木糖、鼠李糖試驗均呈現陰性。

表2 菌株GXUN-20的生理生化試驗結果Table 2 Physiological and biochemical tests results of strain GXUN-20
2.3.3 菌株GXUN-20抑菌實驗結果 幾丁質是真菌細胞壁的主要成份[23],幾丁質酶可通過水解作用將幾丁質裂解為可溶性寡糖[24],從而裂解病原真菌細胞壁使其死亡[25],利用分泌幾丁質酶菌株對植物病原真菌進行防治是目前生防領域研究熱點之一[26]。因此,實驗菌株在細菌病原菌的基礎上加入真菌病原菌,探究GXUN-20菌株是否對細菌和真菌病原菌具有抑制效果。抑制實驗結果如圖5所示,GXUN-20對細菌金黃色葡萄球菌、大腸桿菌、枯草芽孢桿菌、真菌白色鏈霉菌、尖孢鐮刀菌沒有明顯的抑菌圈,初步判斷沒有抑制作用,在真菌黑曲霉、立枯絲核兩株真菌生長范圍內看見較明顯抑菌圈,初步判斷該菌株對部分真菌有抑制作用,GXUN-20菌株抑制黑曲霉抑菌圈平均直徑為17.3 mm,抑制立枯絲核抑菌圈平均直徑為23.8 mm。
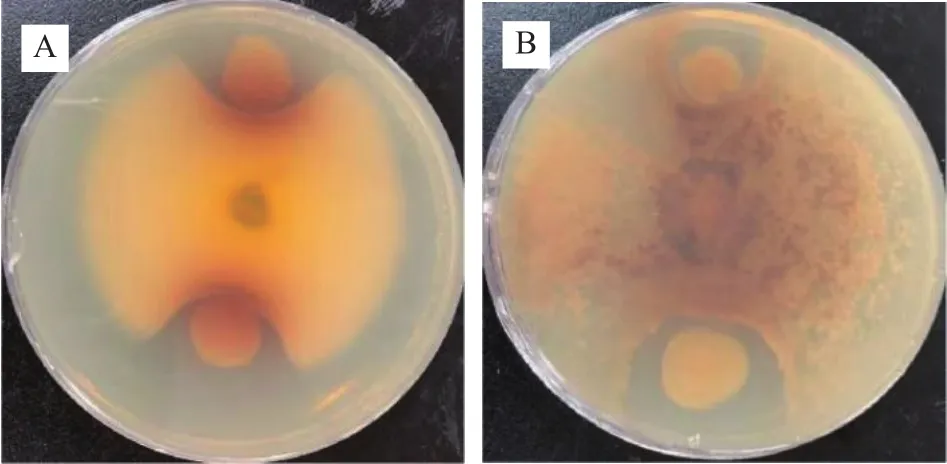
圖5 菌株GXUN-20抑制真菌實驗結果Fig.5 Experimental results of fungi inhibition by strain GXUN-20
2.4 單因素實驗結果
2.4.1 生長曲線及其最適發酵產酶時間的確定GXUN-20菌株的生長曲線及發酵時間對該菌株產酶的結果如圖6所示。菌株GXUN-20在發酵條件下培養1~3 d繁殖數量少,處于生長延滯期,酶活力也相對較低,3~6 d,處于生長對數期增殖加快,發酵液里菌體濃度增大,在這一階段幾丁質酶活力也快速升高,并在第6 d菌液濃度和幾丁質酶活力達到最高,最高酶活為 0.689 U/mL,6~8 d,進入衰亡期,可能是由于發酵液里的營養物質消耗過多,產物的積累、pH變化等不利因素的影響[27],菌株繁殖速度逐漸降低,幾丁質酶活力也相對減弱。通過對GXUN-20菌株生長曲線和發酵時間的監測最后確定該菌株發酵產酶的最適時間為6 d。
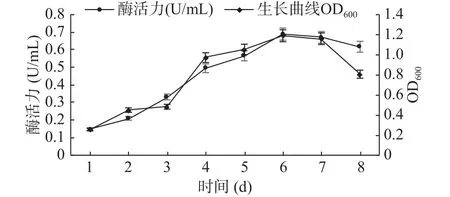
圖6 菌株GXUN-20的生長曲線以及最適發酵產酶時間曲線Fig.6 Growth curve and optimal fermentation time curve of strain GXUN-20
2.4.2 碳源濃度的確定 幾丁質粉末為GXUN-20菌株發酵產酶的唯一碳源,最適添加量對產酶酶活力影響如圖7所示。未優化之前該菌株酶活力為0.140 U/mL,幾丁質粉末添加量為16.67 g/L時,產幾丁質酶活力最高,可達到0.351 U/mL,酶活力約為優化前的2.51倍,因此確定GXUN-20菌株發酵產酶的碳源最適添加量為16.67 g/L。碳源添加量過少時,菌體生長所需碳源不足,導致菌株生長不充分,酶活力降低,碳源濃度增大,在發酵過程中發酵液會越來越粘稠,搖瓶不充分,導致酶活力降低[8]。
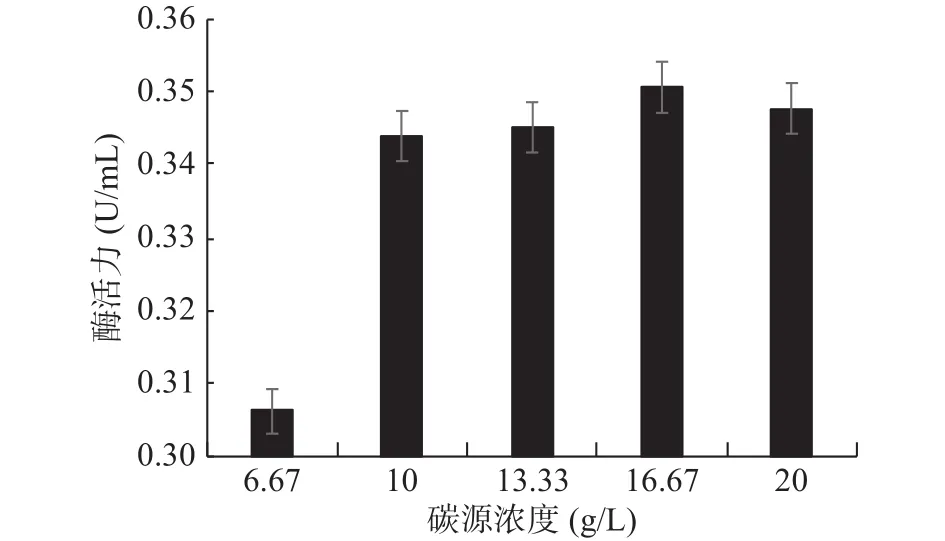
圖7 菌株GXUN-20碳源濃度的確定Fig.7 Determination of carbon source concentration of strain GXUN-20
2.4.3 最適氮源的篩選 氮源是微生物生長必不可少的營養物質之一,也是生物體酶、結構蛋白的重要來源[28],可作為代謝的前體物質,直接參與次生代謝[29]。GXUN-20最適氮源篩選結果如圖8所示,該菌株發酵產酶的氮源分為無機氮和有機氮,以無機氮硫酸銨和氯化銨為氮源時該菌株的酶活分別達到0.570和0.519 U/mL,以黃豆餅粉為氮源時該菌株酶活力最高,酶活力為0.600 U/mL,是優化前的4.29倍。有研究表明與其他氮源相比,黃豆餅粉營養更豐富,更容易被利用,是因為黃豆餅粉不僅僅提供必要的氮元素之外,還提供了很多必需的生長因子[30]。因此,確定GXUN-20菌株發酵產酶的最適有機氮源為黃豆餅粉,硫酸銨是較好的無機氮源,也說明該菌株既能利用有機氮源也能利用無機氮源。

圖8 菌株GXUN-20最適氮源的篩選Fig.8 Screening of optimum nitrogen source for strain GXUN-20
2.4.4 最適氮源添加量的確定 氮源添加量對GXUN-20菌株發酵產酶酶活力的影響如圖9所示。低濃度和高濃度的黃豆餅粉都不利于該菌株發酵產幾丁質酶,馬璐等[31]研究發現低濃度氮源不足以維持菌體生長需要,當氮源濃度高時相對碳源濃度低,細菌沒有充足能量合成蛋白質和進行生命活動,因此合適的碳氮比可促進菌體生長及次生代謝產物的積累。隨著黃豆餅粉濃度的增加,幾丁質酶活力先增加后下降,在黃豆餅粉達到3 g/L時,該菌株產幾丁質酶活力最高,達到0.486 U/mL,是優化前的3.47倍。因此,確定GXUN-20菌株發酵產酶的最適氮源添加量為3 g/L。

圖9 菌株GXUN-20最適氮源黃豆餅粉添加量的確定Fig.9 Determination of the optimum nitrogen source for soybean cake powder of strain GXUN-20
2.4.5 最適溫度的確定 GXUN-20菌株酶活力隨溫度變化結果如圖10所示。低溫菌體生長減緩,釋放酶蛋白減少,總體酶活性不高,在一定溫度范圍內,隨著溫度的上升,呼吸強度增加,代謝速度增強,最終導致細胞生長繁殖加速[32],幾丁質酶活力上升,在30 ℃時,幾丁質酶活力達到最高為0.458 U/mL,是優化前的3.27倍,隨著溫度繼續升高,高溫會讓酶蛋白失活變性,酶活力有所下降,因此,確定GXUN-20菌株發酵產酶的最適溫度為30 ℃。

圖10 菌株GXUN-20最適溫度的確定Fig.10 Determination of the optimum temperature of strain GXUN-20
2.4.6 發酵液初始最適pH的確定 菌株GXUN-20發酵液初始最適pH對酶活力的影響結果如圖11所示。pH偏低,發酵液偏酸性,不利于菌體生長,酶活力不高,在一定pH范圍內,隨著pH的升高,發酵液偏弱堿性,有利于該菌株生長,酶活力增強,在pH=8.4時,GXUN-20菌株產幾丁質酶活力最大,為0.659 U/mL,是優化前的4.71倍,隨著pH繼續增大,發酵液偏堿性,堿性會讓酶蛋白變性失活,酶活力減弱,表明該菌株適合弱堿性的產酶環境,可能是因為該菌株生長在堿性的海岸環境中,而大部分細菌最適發酵產酶的pH在6.3~7.5[32]。

圖11 菌株GXUN-20最適發酵液pH的確定Fig.11 Determination of the optimum pH of fermentation liquor of strain GXUN-20
2.4.7 發酵液最適接種量確定 GXUN-20菌株發酵液最適接種量優化結果如圖12。接種量在2%(v/v)時,GXUN-20產幾丁質酶活力最大,為0.501 U/mL,是優化前的3.58倍。隨著接種量的增加,過高的接種量會極具地消耗培養基中的氧氣和營養物質[17],所以菌株的酶活力逐漸減弱,因此,GXUN-20菌株發酵產酶的最適接種量為2%。
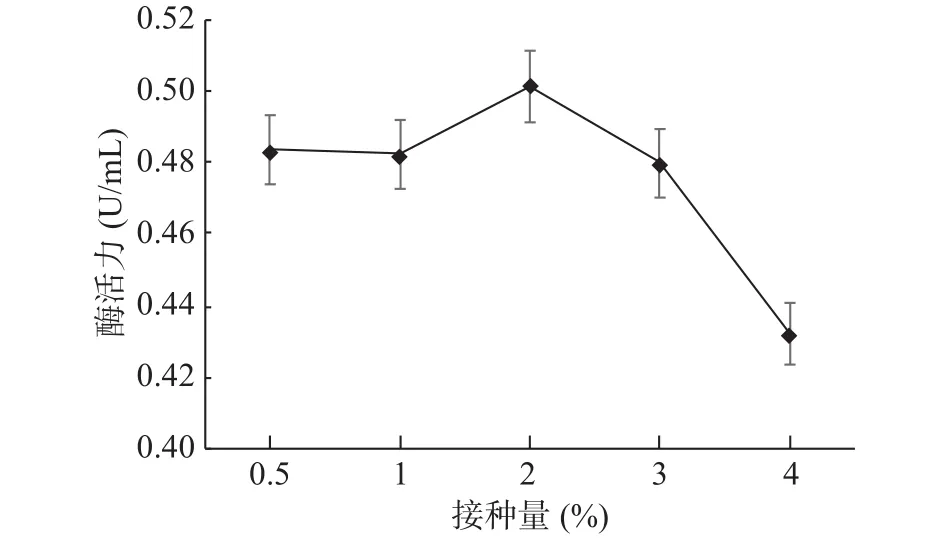
圖12 菌株GXUN-20最適接種量的確定Fig.12 Determination of optimum inoculation amount of strain GXUN-20
2.5 正交試驗結果
在單因素實驗結果的基礎上設計L16(44)正交試驗,對因素A:氮源黃豆餅粉濃度、B:pH、C:碳源幾丁質粉末濃度、D:接種量,4因素之間的相互作用做進一步的研究。正交試驗結果見表3,根據實驗結果計算出kn值和R值,得出的最佳發酵方案為:A1B3C4D4,即黃豆餅粉 1 g/L,發酵液初始 pH8.4,幾丁質粉末 20 g/L,接種量 3%;R值 A>C>B>D,A為最顯著因素,正交實驗方差分析結果見表4,得出黃豆餅粉對試驗結果影響顯著(P=0.013<0.05),即氮源對GXUN-20菌株產酶影響最為明顯。以正交試驗得出GXUN-20菌株最適發酵產酶條件進行產酶酶活力驗證,黃豆餅粉1 g/L,幾丁質粉末20 g/L,復合鹽母溶液10 mL,pH8.4,接種量3%,200 r/min、30 ℃振蕩培養6 d后,檢測液體酶活力為0.965 U/mL,是優化前的6.89倍。
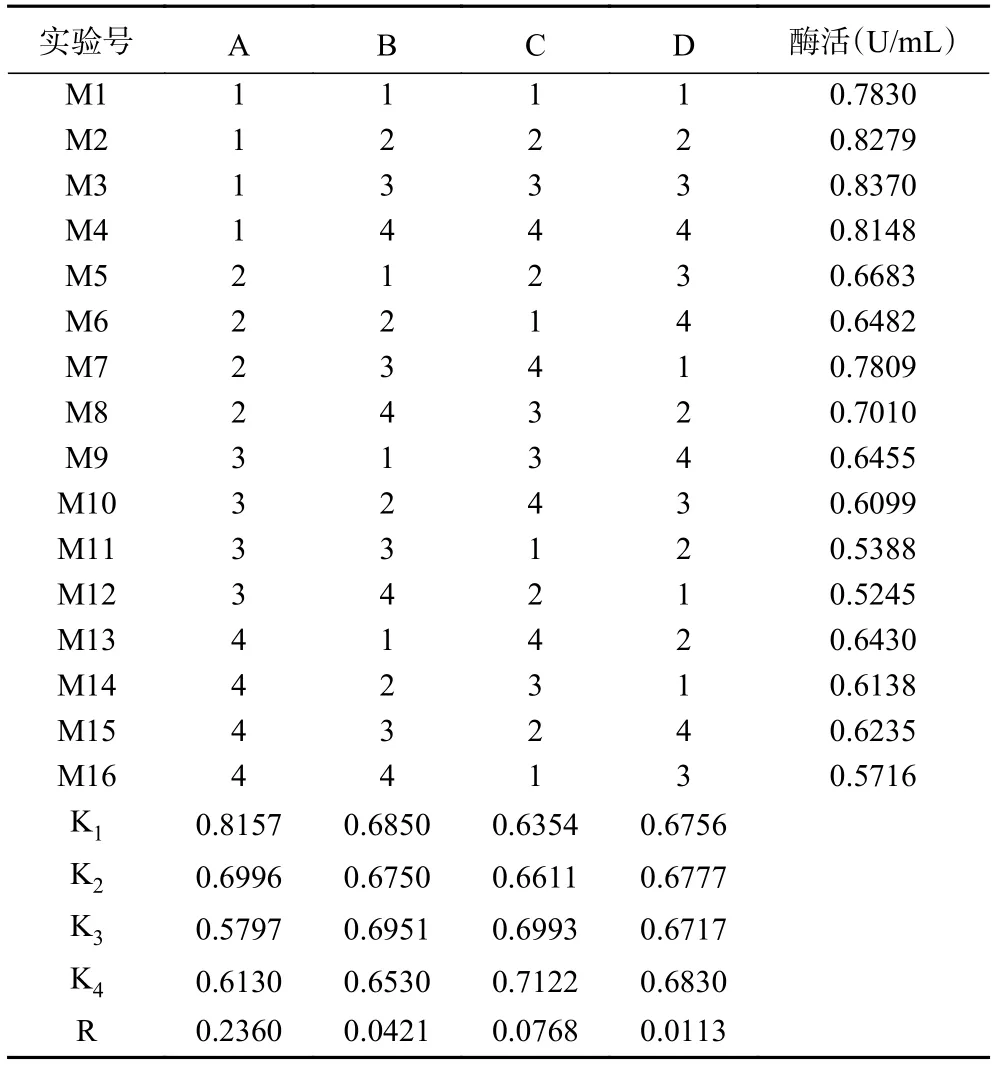
表3 L16(44)正交試驗結果Table 3 Results of L16 (44) orthogonal experiment

表4 GXUN-20菌株正交實驗方差分析Table 4 Variance analysis of orthogonal test of GXUN-20 strain
3 結論與討論
自然環境中蝦蟹殼的降解主要是由微生物完成的,為了提高蝦蟹等海產品廢棄物利用率及其附加值,本研究從廣西合浦縣大神木海鴨場附近灘涂沉積物(蝦蟹及貝殼廢棄物)中篩選出一株產幾丁質酶菌株GXUN-20,經16S rDNA序列構建系統發育樹,鑒定與Lysobacter firmicutimachus相似度為99.79%,為厚壁溶桿菌,與蝦蟹殼土壤[33]、黃粉蟲蟲蛻[16]中篩選到的幾丁質降解菌株類芽孢桿菌屬的結果不一致,說明產幾丁質酶菌株分類廣泛。以幾丁質酶活力為指標,對GXUN-20菌株進行發酵條件單因素優化,在單因素優化結果較顯著的4因素(氮源濃度、發酵液初始pH、幾丁質粉末濃度、接種量)基礎上進行4水平正交試驗及正交試驗方差分析。結果顯示:氮源濃度對產酶影響顯著(P=0.013<0.05),在氮源為1 g/L黃豆餅粉,幾丁質粉末為20 g/L,接種量為3%,發酵液初始 pH 8.4,30 ℃、200 r/min發酵 6 d條件下,發酵液中幾丁質酶活力最大,達0.965 U/mL,是優化前酶活力的6.89倍。本研究結果為GXUN-20菌株產幾丁質酶進一步開發與應用提供了基礎數據和實驗方法,也為進一步探究降解蝦蟹殼酶法生產殼聚糖提供實驗基礎。
目前國內外對溶桿菌屬產幾丁質酶方面的研究較少,有研究發現Lysobacter enzymogenes菌株可產幾丁質酶[20],Qian 等[34]、朱慧等[35]研究Lysobacter enzymogenes對辣椒疫霉具有抑制作用以及該菌株產幾丁質酶的分子機制。由于Lysobacter firmicutimachus菌株產幾丁質酶相關研究缺乏,本研究通過單因素實驗優化了GXUN-20菌株產幾丁質酶發酵培養條件,通過對該菌株的生長曲線以及最適產酶時間的測定,得出該菌株生長密度與產酶活力呈正相關,在一定時間范圍內,隨著該菌株的生長,發酵液里菌體濃度不斷增加,該菌株降解幾丁質產還原糖的能力增加,即幾丁質酶活力增加,并在菌體生長達到頂峰時,幾丁質酶活力也達到最大,這與陳立功等人的研究基本一致[36]。隨著發酵時間延長,發酵環境越來越不適合該菌株的生長,發酵液中氧氣和營養物質的消耗以及代謝物的增加,菌株濃度慢慢降低,幾丁質酶活力也呈現下降的趨勢[27]。
幾丁質粉末為該菌株發酵培養的唯一碳源,在單因素優化中,分別以不同濃度的碳源進行發酵產酶活力測定,結果在幾丁質粉末為16.67 g/L時,GXUN-20菌株產酶活力最高,碳源添加量過少,菌體生長所需碳源不足,導致菌株生長不充分,酶活力降低,碳源濃度增大,在發酵過程中發酵液越發粘稠,搖瓶不充分,導致酶活力降低。不同氮源對GXUN-20菌株產幾丁質酶具有明顯的影響,以硫酸銨、氯化銨、黃豆餅粉作為氮源時該菌株產幾丁質酶活力較高,以蛋白胨、牛肉膏、尿素為氮源時該菌株幾丁質酶表達量較低,有機氮源黃豆餅粉單因素優化之后該菌株產酶活力最大,但與無機氮源硫酸銨差別不大,為考慮到工業生產成本問題,后期可以考慮選擇無機銨鹽代替黃豆餅粉進行工業發酵生產。
pH能影響菌株形態、生長以及對營養物質吸收和代謝物產生[32]。在菌株GXUN-20 pH單因素優化研究中,前期pH單因素研究設計pH范圍為4~10,得出最適產酶pH為8,考慮到較強度的酸堿會影響菌株產酶能力,所以在前期的實驗基礎上縮小pH優化范圍,最后pH優化范圍為7.8~9.0,得出該菌株最適發酵產酶pH為8.4。在單因素溫度優化研究中GXUN-20菌株最適產酶溫度為30 ℃,隨著溫度的升高,酶蛋白失活變性,GXUN-20菌株產幾丁質酶活力降低。
幾丁質酶是一類差異較大的水解酶,由于菌株的來源不同,其酶學性質不盡相同,幾丁質酶對底物的親和力也不同[16],本研究測定酶活力是以幾丁質粉末為底物,還需進一步使用蝦蟹殼粉末為底物,測定該菌株產生的幾丁質酶對蝦蟹殼粉末的降解效果,利用幾丁質酶直接降解蝦蟹殼粉末,制備蝦蟹殼降解產物,才能真正地實現蝦蟹殼廢棄物的資源化利用。研究結果還發現GXUN-20對黑曲霉、立枯絲核具有抑菌作用,因此,可以嘗試將該菌株投入大田實驗,研究生防菌劑來達到生物防治植物真菌病害的目的。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
