基于高通量測序的白木香結香部位真菌多樣性研究
2021-12-23 16:55:29宋杰張寧南張鐘慧周再知梁俊峰陸俊錕
熱帶作物學報 2021年11期
宋杰 張寧南 張鐘慧 周再知 梁俊峰 陸俊錕

摘 ?要:本研究以廣東茂名和東莞區域的經濟植物白木香(Aquilaria sinensis)為對象,基于高通量測序方法,分析2個不同區域生境中4個品種白木香結香部位真菌群落結構多樣性及分布規律,結合不同品種的結香特性,進一步挖掘與白木香結香顯著相關的真菌類群,促進結香技術的提升。結果表明:相同生境條件下,同種白木香不同高度結香位點真菌組成結構非常相似,但不同真菌的豐度存在差異;不同區域生境下,相同品種結香部位真菌群落組成結構存在顯著差異,生境差異可能是導致結香部位真菌群落結構差異的主要因素之一;同一區域生境下,不同品種白木香結香部位的真菌群落結構存在顯著差異,品種差異可能是導致白木香結香部位真菌群落結構差異的主要因素之一,同時特有真菌類群的存在也可能是影響白木香結香特性的關鍵。Fusarium spp. 和Hypomontagnella spp. 等真菌在易結香品種分布廣泛且豐度極高,可能與易結香品種的結香特性相關,具有重要的應用潛力。
關鍵詞:白木香;結香技術;真菌多樣性;高通量測序
中圖分類號:S567.1 ? ? ?文獻標識碼:A
Abstract: Aquilaria sinensis (Lour.) Spreng, an economic plant in Guangdong, was selected as the object of this study. Using the high-throughput sequencing method, we analyzed the community structure and distribution of fungi in agar-wood wound locations of four agarwood varieties in Maoming and Dongguan, China. Combining the agarwood forma-tion characteristics of different varieties, fungal groups were further investigated which were significantly related to the agarwood formation. Under the same habitat condition, fungi community structure at different height sampling sites in the same varieties were remarkably similar, despite the different fungi abundance. The fungal community composition were significantly different in the same varieties under different habitats, and the difference of habitat may be one of the main factors leading to the difference of fungal community structure. There were significant differences in the fungal community structure among different varieties under the same habitat condition. Different agarwood varieties may be one of the main factors leading to the difference of fungal community structure. The unique fungi with high abundance maybe result in special feature of agarwood formation. Fusarium spp. and Hypomontagnella spp., which are widely distributed with high abundance in the easy agarwood formation varieties, are the best illustrations for this conclusion.
Keywords: Aquilaria sinensis; agarwood formation technology; fungi divesity; high-throughput sequencing
DOI: 10.3969/j.issn.1000-2561.2021.11.039
沉香是由瑞香科沉香屬(Aquilaria spp.)或擬沉香屬(Gyrinops spp.)植物木質部組織及其分泌物共同組成的混合物質,在亞洲、中東和歐洲被廣泛用于醫藥,同時也是一種傳承歷史悠久的名貴香料[1-3]。沉香內含的倍半萜烯和苯乙基色酮派生物具有顯著的生物學藥理性質,包括抗菌、抗氧化和抗腫瘤活性,在治療胃潰瘍和哮喘方面具有潛在的價值[4-8]。
在我國,沉香主要來源于白木香[Aquilaria sinensis (Lour.) Spreng],是一種重要的傳統中藥材,可用于治療嘔吐、哮喘和失眠[9-10]。自然條件下,白木香樹在受到微生物刺激、昆蟲以及其他傷害后可產生分泌物,沉積形成沉香,過程復雜且需要長期的演變和積累[11]。然而,由于沉香的神秘香味和高昂價格,致使世界范圍內的沉香過度采伐和消耗,沉香日漸成為一種緊缺資源。目前所有的沉香屬物種都被列入《瀕危野生動植物種國際貿易公約》(CITES, http://www.cites.org),其進出口需要相關許可證(IUCN 2002)。在2004年,白木香又被列為國家二級保護植物,并被列入《世界自然保護聯盟受威脅植物紅色名錄》[12]。
為了保護野生沉香資源,同時增加沉香產量,我國廣東、海南和廣西等地開始了廣泛的白木香培育及促進白木香結香技術研發工作。真菌侵染長期以來被認為是一種有效促進白木香結香方法,許多從白木香結香部位分離的真菌已被報道可以顯著加速沉香油性成分形成和積累,包括Fusarium sp.、Chaetomium sp.、Trichoderma sp.、Menanotus sp.、Aspergillus sp.、Penicillium sp.、Lasiodiplodia sp.和Rigidoporus sp.等[11, 13-17]。張苗苗[18]對白木香內生菌的分布規律進行了初步研究,結果發現,白木香內生真菌在結香過程中可能具有復雜的規律性,不是簡單線性關系,結香部位菌群豐富,結構非常復雜。Liu 等[2]的研究結果也顯示,白木香不同高度結香部位的真菌類群有明顯差異,結香品質也不相同。真菌分布規律及刺激白木香結香的機制研究仍有待深入。
目前,在長期的人工培育和結香實踐過程中發現,我國白木香不同個體的結香特性并不相同,通過篩選發現多個易結香品種,在地方上被稱為“奇楠沉香”[19-23]。普通白木香品種沉香的形成和累積都較為緩慢,通常樹齡超過10 a的白木香經過物理、化學、生物等的刺激才會結香;而少部分經過篩選白木香品種結香卻較為容易,3~5 a樹齡的白木香即可開始產香,并且結香品質較高,油脂豐富、色澤較深,即所謂的“奇楠沉香”。然而,迄今仍然沒有系統性的研究,探討不同地區、不同品種白木香結香部位的真菌種群分布規律。因此,研究并闡明這種真菌種群分布規律對于探究真菌促結香技術的研發具有重要價值。
鑒于此,本研究選擇4個不同品種白木香的物理傷害結香樣本,通過高通量測序的方法,結合已有研究數據進行真菌物種多樣性分析,旨在:(1)探究不同地區環境中白木香結香部位真菌類群多樣性之間的關系;(2)探究不同品種白木香結香部位真菌類群分布規律,闡明真菌分布差異與結香特異的關聯性,挖掘更有價值的促結香真菌。本研究將為探究白木香結香技術提供重要的參考和數據支撐。
1 ?材料與方法
1.1 ?樣本采集
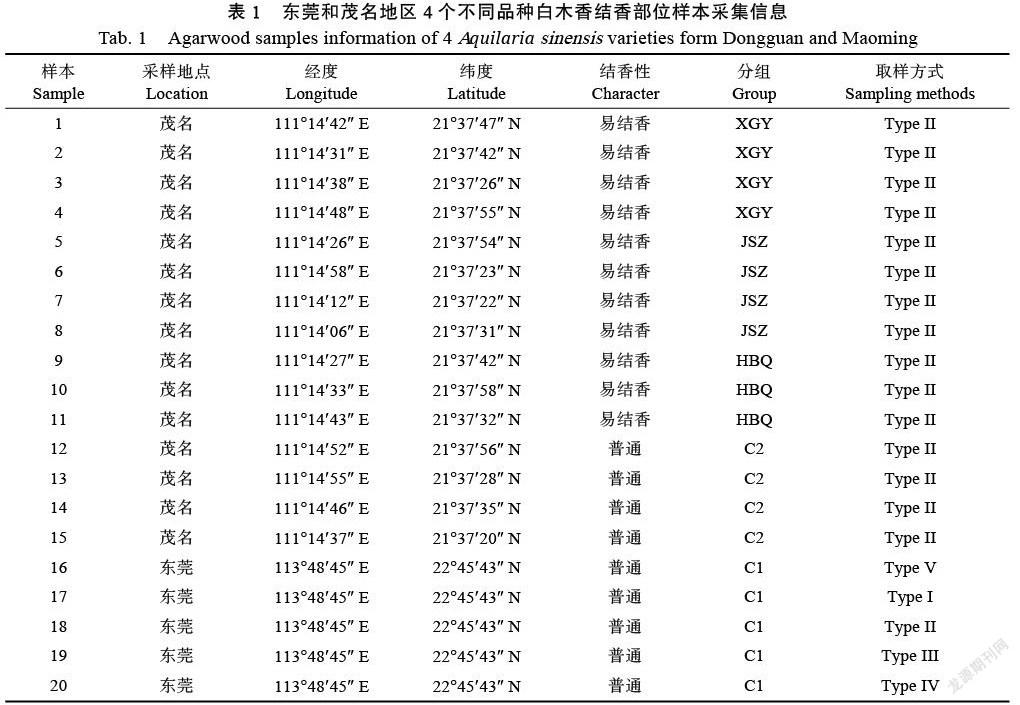
2019年10月中旬,在廣東省茂名市電白區大水坡村進行不同品種白木香樣本采集。取樣方式參照Liu等[2]Type II(Type I:距離地面2~2.5 m的損傷主干橫截面結香部位;Type II:距離地面1.5~1.8 m處損傷側枝結香部位;Type III:距離地面1~1.2 m的損傷主干橫截面結香部位;Type IV:距離地面0.8~1.2 m處損傷主干縱切面結香部位;Type V:距離地面0.2~0.4 m,靠近根部的損傷主干結香部位),每個品種確保隨機3~4份重復,依次做好標記。易結香奇楠品種3種,包括‘西瓜葉’(XHY)、‘金勺子’(JSZ)和‘虎斑棋’(HBQ)。普通沉香品種1種,與Liu 等[2]所述東莞地區白木香相同,皆為廣東省內常見的普通不易結香的對照品種,分別標記為C1和C2。采集區域及沉香品種詳情見表1。每份樣本確保取樣量大于2 g,取樣后首先使用無菌蒸餾水進行表面清洗,然后使用無菌濾紙吸干,處理后的樣本快速放入無菌凍存管,并立即進行液氮速凍。所有樣品處理方法一致,低溫保存帶回實驗室,轉移到?80 ℃冰箱保存備用。
1.2 ?樣品真菌DNA提取及文庫構建
準確稱取2 g凍存樣品供真菌多樣性分析,所有樣本送往上海美吉生物工程有限公司進行高通量測序。利用DNA提取試劑盒(E.Z.N.A.? soil DNA kit,Omega Bio-tek, Norcross, GA, U.S.)進行微生物總DNA抽提,使用1%的瓊脂糖凝膠電泳檢測抽提的基因組DNA質量;使用NanoDrop2000測定DNA濃度和純度。本文使用ITS(Internal Transcribed Spacer)基因序列作為分子標記,使用引物ITS1F(5-CTTGGTCATTT-AGAGGAAGTAA-3)和ITS2R(5-GCTGCGTTC- TTCATCGATGC-3)進行PCR擴增。每份樣本分別重復擴增3次。將同一份樣本的PCR產物混合后使用2%瓊脂糖凝膠回收,利用AxyPrep DNA Gel Extraction kit(美國Axygen Biosciences)進行回收產物純化。2%瓊脂糖凝膠電泳檢測,并用QuantusTM Fluorometer(美國Promega)對回收產物進行檢測、定量。使用NEXTFLEX Rapid DNA-Seq Kit試劑盒構建測序文庫,然后在Illumina公司的Miseq PE300平臺上機進行雙端測序。
1.3 ?數據處理
1.3.1 ?原始數據整理、過濾及質量評估 ?從GenBank數據庫的SRA庫中,下載廣東省東莞市普通白木香結香處微生物高通量測序數據(BioProject number: PRJNA509099),結合本研究測序結果進行比較研究。本研究采用最新的性能更優的QIIME2[24](Quantitative Insights Into Microbial Ecology)分析平臺對原始數據(Raw data)進行分析,分析方法參照qiime2(https:// docs.qiime2.org)的使用說明。將數據導入QIIME2平臺后,調用q2-cutadapt插件先對reads引物序列進行剪切,然后根據序列質量分布圖,調用q2-data2插件對擴增子數據去噪,過濾質量較差的序列和嵌合序列,從而確定擴增子序列變異(Amplicon sequence variants, ASVs),然后去冗余,得到有效Fearture,包括特征表(Feature-table)及代表序列(Feature-data)。使用q2-feature-table插件過濾特征,過濾僅在一個樣本中出現的特征,避免偶然因素誤差,降低結果的假陰性率。使用相同的方法處理代表序列。
1.3.2 ?原始數據整理、過濾及質量評估 ?基于代表性Feature序列和真菌UNITE數據庫(https://unite.ut.ee/)進行序列比對,即對不同分組樣本中檢測到的全部ITS有效序列,自分類學上的界(Kingdom)、門(Phylum)、綱(Class)、目(Order)、科(Family)、屬(Genus)、種(Species)多個層級進行物種鑒定和注釋。查看代表序列和注釋結果文件,并在NCBI數據庫中進行Blast校對,去除雜質的序列,使用q2-taxa插件再次過濾特征表和代表序列,僅保留注釋到門的物種序列。依據過濾后的特征表,生成最終的樣本物種注釋結果,并對檢測到的真菌物種進行菌群群落組成分析,計算每個分類層級上每個物種的豐度占全部物種豐度的比值(即相對豐度)。使用R(v.3.6.0)軟件中ggplot2程序包繪制不同樣品在各個分類層級上的真菌堆疊柱狀圖,反映物種的組成情況。
1.4 ?統計分析
依據測序特征表結果,計算ASVs水平α多樣性,得到Richness指數,使用ggplot2軟件繪制Richness指數稀釋曲線,確定物種覆蓋度是否已經飽和。同時,依據屬水平的物種注釋結果,使用q2-diversity插件及R軟件中vegan程序包進行Alpha多樣性計算,包括Richness指數、Chao1指數、ACE指數、Shannon指數和Simpson指數,比較不同組間樣本的多樣性水平。Richness指數、Chao1指數和ACE指數反映物種豐富度,指數越大表示樣本群落豐富度越高;Shannon指數和Simpson指數反映物種的豐富度和均勻度,即多樣性,指數值越大,表示菌群多樣性越高。
本研究計算樣本距離的算法采用Bray- Curtis distance、Unweighted/Weighted UniFrac distance。本研究采用主坐標分析(PCoA)進行β多樣性分析,PCoA圖可直觀展示組間的菌群組成差異,若兩組間樣本距離較近,提示2組樣本菌群結構相似性高,菌群組成差異性小,反之則表示2組間樣本間菌群群落差異性大。進一步,本研究利用R軟件包對不同分組中豐度大于1%的物種繪制了豐度熱圖,并利用STAMP v2.1.3軟件進行屬水平真菌豐度差異顯著性分析Welch’s t-test檢驗,識別不同分組中豐度差異顯著的真菌類群[25]。
2 ?結果與分析
2.1 ?真菌多樣性測序結果
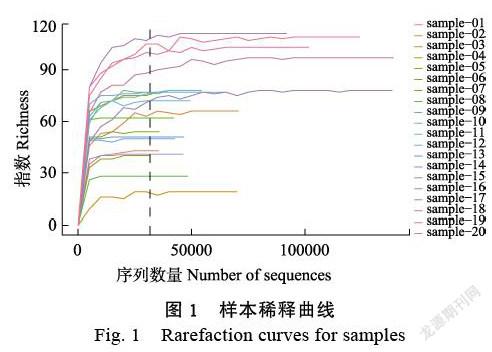
本研究包含白木香普通品種及易結香品種4個,總樣本20份,其中廣東省茂名市樣本15份,東莞市樣本5份。根據品種、采集地點和結香特性分別將樣本分為5組(表1)。高通量測序獲得序列(reads)歷經數據優化、拼接、過濾和質控后獲得優質ITS序列1349058條,片段長度150~308 bp,包含463個ASVs。由圖1可知,所有樣本的Richness稀釋曲線趨于平緩,ASVs覆蓋度已經達到飽和,說明所測數據能夠準確反映樣本真菌群落結構組成。
2.2 ?物種分類注釋
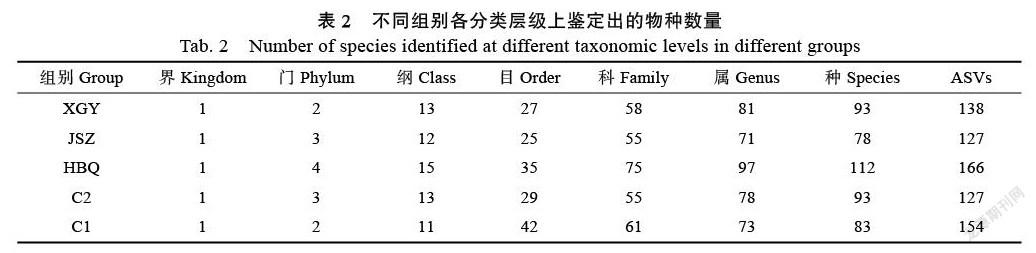
利用UNITE分類器,以及NCBI數據庫對Feature數據的注釋,463個ASVs可鑒定到真菌界4門、18綱、61目、29科、189屬、231種真菌。各分類水平的微生物類群統計結果見表2,絕大部分菌群都分類到了屬,也有部分菌群分類到了種,但仍然有很多Feature無法分類到種。依據分類數據,本研究在目水平和屬水平繪制了真菌堆疊柱狀圖,反映各組物種的組成情況,見圖2。
2.3 ?群落組成分析
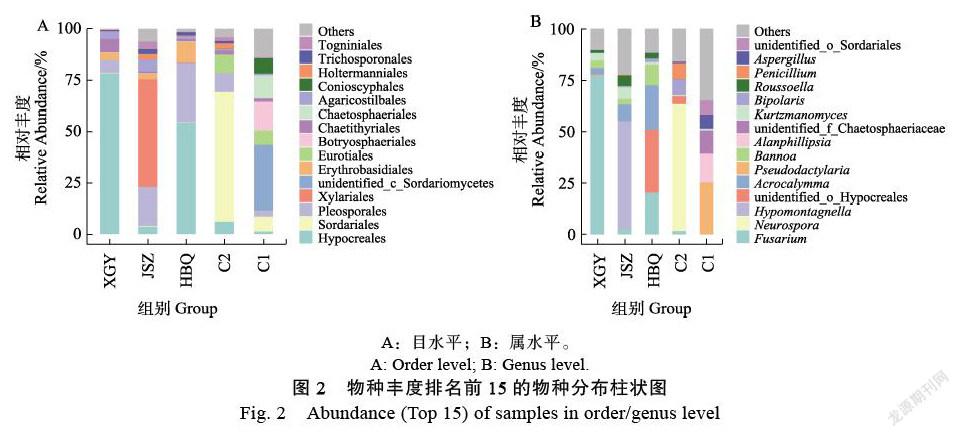
在目分類水平上,共鑒定61目真菌,其中,未知分類菌目有6個,占到13%,5組樣本中真菌分布結構存在差異。如圖2A所示,XGY組檢測出分類菌目27個,Hypocreales是明顯優勢種群,占比達78.4%,其次為Chaetothyriales(6.7%)、Pleosporales(6.0%)、Erythrobasidiales(3.6%)和Agaricostilbales(3.3%),五類菌占比都超過1%,所占比例之和達到98%。JSZ組樣本檢測出分類菌目25個,Xylariales是明顯優勢種群,占比52.4%,占比超過1%的目還包括Pleosporales(18.9%)、Agaricostilbales(5.5%)、Togniniales (3.7%)、Hypocreales(3.7%)、Glomerellales(3.2%)、Holtermanniales(2.7%)、Erythrobasidiales(2.7%)、Trichosporonales(2.5%)和Tremellales(1.8%)。HBQ組樣本檢測出分類菌目35個,Hypocreales是明顯優勢種群,占比達54.2%,占比超過1%的目還包括Pleosporales(28.9%)、Erythrobasidiales(9.9%)、Trichosporonales(1.6%)、Chaetothyriales(1.3%)和Agaricostilbales(1.1%)。C2組樣本檢測出分類菌目29個,占比超過1%的目有9個,其中Sordariales是明顯優勢種群,占比63.2%,其次為Pleosporales(9.3%)、Eurotiales(8.7%)、Hypocreales(6.1%)、Holtermanniales(2.7%)、Chaetothyriales(2.1%)、Togniniales(1.9%)、Trichosporonales(1.2%)和Glomerellales(1.1%)。C1組樣本檢測出分類菌目42個,占比超過1%的目有13個,其中norank Sordariomycetes為優勢菌,占比達25.2%,其次為Botryosphaeriales(13.8%)、Chaetosphaeriales(11.4%)、Conioscyphales(7.4%)、Sordariales(7.3%)、Eurotiales(6.9%)、unidentified Sordariomycetes(6.9%)、Helotiales(5.7%)、Wallemiales(3.5%)、Pleosporales(3.0%)、Thelebolales(2.4%)、Chaetothyriales(1.7%)、Hypocreales(1.3%)。
在屬水平上檢測分類出189類真菌屬。如圖2B所示,XGY組中共有81類真菌屬,其中Fusarium屬真菌豐富最高(77.6%),其次為Veronaea(5.7%)、Bannoa(3.6%)、Kurtzmanomyces(3.3%)、Acrocalymma(2.7%)、Roussoella(1.6%)、Corynespora(1.1%)、Cyphellophora(0.6%)和unidentified Hypocreales(0.6%)。JSZ組中共有71類真菌屬,其中Hypomontagnella屬真菌豐富最高(52.4%),其次為Acrocalymma(8.0%)、Kurtzmanomyces(5.5%)、Roussoella(5.1%)、Phaeoacremonium(3.7%)、Colletotrichum(3.2%)、Holtermanniella(2.7%)、Bannoa(2.7%)、Fusarium(2.6%)、unidentified Pleosporales(1.8%)和Cutaneotrichosporon(1.5%)。HBQ組中共有97類真菌屬,其中unidentified Hypocreales豐富最高,占比30.7%,其次為Acrocalymma(21.6%)、Fusarium(20.1%)、Bannoa(9.9%)、Kiflimonium(3.3%)、Roussoella(2.5%)、Bipolaris(1.7%)、Kockovaella(1.2%)、Pseudorobillarda(1.2%)和Kurtzmanomyces(1.2%)。C2組中共有78類真菌屬,Neurospora屬真菌豐富最高,占比達61.2%,其次為Penicillium(7.8%)、Bipolaris(7.6%)、unidentified Hypocreales(3.8%)、Holtermanniella(2.7%)、Phaeoacremonium(1.9%)、Fusarium(1.6%)、Cyphellophora(1.6%)、Phialemonium(1.5%)、和Cutaneotrichosporon(1.2%)。C1組中共有73類真菌屬,Pseudodactylaria豐度最高,達25.2%,其次為Alanphillipsia(13.8%)、unidentified Chaetosphaeriaceae(11.3%)、unidentified Sordariales(6.9%)、Aspergillus(6.9%)、unidentified Sordariomycetes(6.6%)、unidentified Conioscy-phaceae(4.5%)、Wallemia(3.5%)、Claussenomyces(2.9%)和unidentified Helotiales(2.9%)。
2.4 ?真菌群落多樣性指數分析
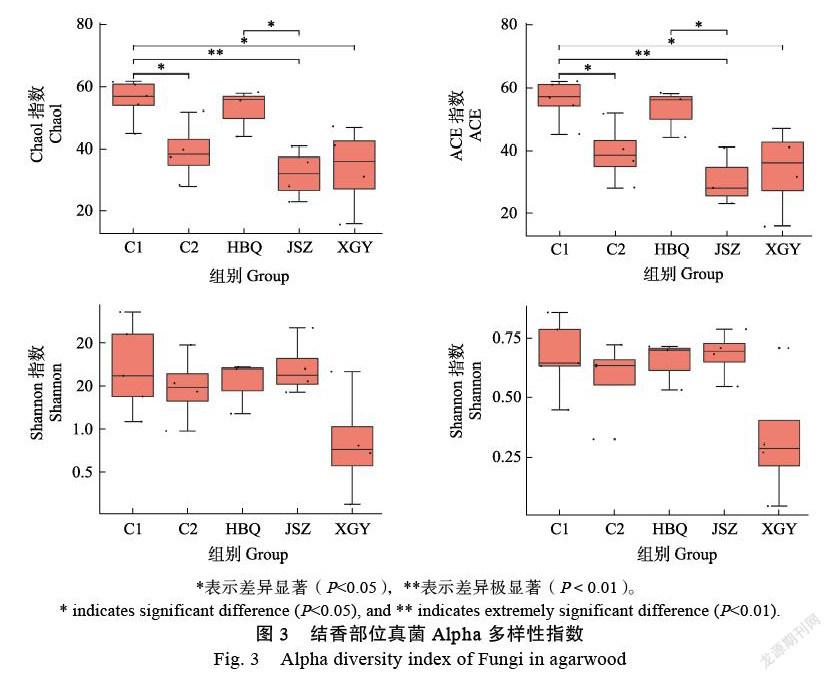
基于Feature豐度和vegan軟件包計算的多樣性指數值,使用T檢驗分析并比較各分組樣本的多樣性指數。如圖3所示,C1組Chao1指數和ACE指數高于其他組,表明東莞地區普通白木香結香部位真菌類群比茂名地區豐富度高,且存在統計學差異(P<0.05)或顯著統計學差異(P<0.01)。Chao1指數和ACE指數還顯示茂名地區4組白木香中,易結香品種HBQ結香部位真菌類群豐富度最高,且HBQ與JSZ分組中真菌類群豐富度存在統計學差異,其他分組間不存在顯著的統計學差異。Shannon指數和Simpson指數顯示XGY最低,表明易結香品種XGY結香部位真菌多樣性最低,但各類群間多樣性并不存在統計學差異。
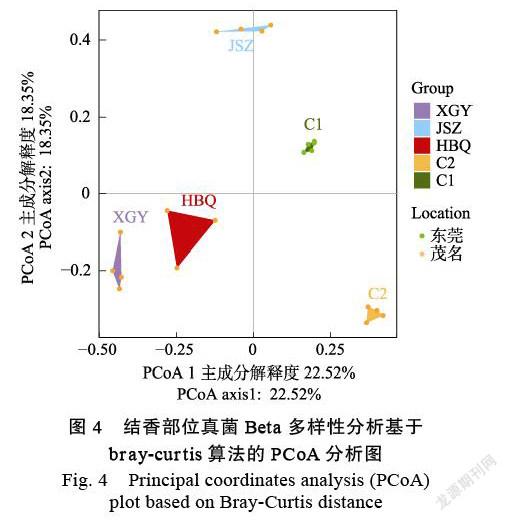
圖4為基于 Bray-Curtis距離的PCoA分析結果,其中PCoA1和PCoA2的主成分解釋度分別為22.52%和18.35%。不同區域、不同品種5個分組的真菌群落形成了各自獨立的區域,每個分組散點分布區域是完全分離的,說明5個分組間菌群結構存在可區分的差異。東莞地區C1組與茂名地區4個分組之間距離都較遠,表明2個地區白木香結香部位菌群結構組成上相似性較低,差異較大。易結香品種XGY和HBQ之間距離最近,表明2組菌群結構相似性較高,菌群組成差異較小;XGY、HBQ都與JSZ之間距離較遠,表明JSZ品種的結香部位菌群結構與其他兩個易結香品種差異較大;易結香品種與普通品種C1、C2組之間距離都相對較遠,表明普通沉香品種與易結香品種之間結香部位菌群結構存在顯著差異。
2.5 ?物種差異分析
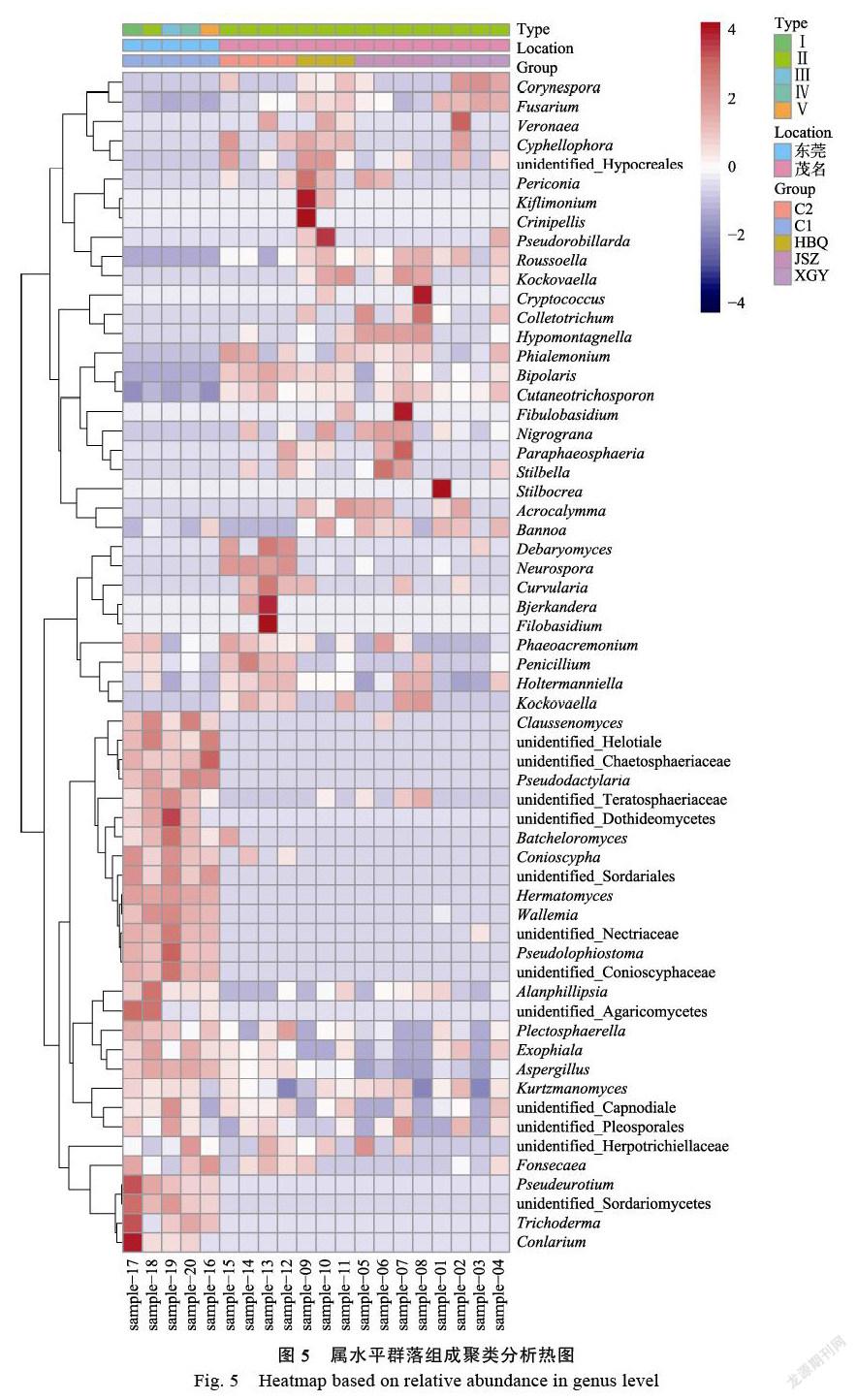
在屬水平,利用大于1%豐度真菌繪制分層聚類熱圖,如圖5所示,東莞地區普通沉香與茂名地區的4種白木香結香部位真菌組成存在顯著的差異,表明不同地區之間氣候環境的差異對結香部位菌群的影響非常顯著。C1組沉香5個不同的樣本源于東莞地區的普通白木香品種結香部位,每個樣本距離地面的高度不同、取樣部位不同,雖然各屬真菌在C1組5個樣本中的真菌物種豐度存在差異,但結香部位的菌群物種組成較為相似,特別是都含有18個較高豐度的屬或未知屬真菌,包括:Claussenomyces、Conlarium、Tricho?derma、Pseudeurotium、Pseudodactylaria、Batche?lor?omyces、Conioscypha、Hermatomyces、Wallemia、Pseudolophiostoma、unidentified_ Conios?cyphaceae、unidentified_Nectriaceae、unidentified_Sordari?omy?ce-tes、unidentified_Dothideomycetes、unide?nti-fied_Sordariales、unidentified_Teratosphaeriaceae、unidentified_Chaetosphaeriaceae、unidentified_Hel??o-tia?les。此18個屬及未知屬真菌在茂名地區C2、HBQ、JSZ、XGY等品種結香部位中豐度都極低。
Alanphillipsia、Plectosphaerella、Exophiala、Aspergillus、Kurtzmanomyces、Phaeoacremonium、Penicillium、Holtermanniella、unidentified_ Capn?od?i-ales、unidentified_Pleosporales等10屬或未知屬在5組中都有廣泛分布,但在普通沉香C1和C2組中通常分布更多。Phialemonium、Bipolaris和Cutaneotrichosporon屬真菌在茂名地區C2、HBQ、JSZ和XGY中普遍豐度較高。Fusarium、Corynes-p?ora、Roussoella、Kockovaella、Acrocaly?mma、Hypomontagnella和Bannoa等7屬真菌僅在易結香品種中廣泛分布且具有較高的豐度,普通品種中分布較少且豐度極低。
Neurospora、Debaryomyceser、Curvularia等3屬真菌僅在C2組普通白木香中普遍存在且豐度較高,與其他組有顯著豐度差異。Cyphellophora、Veronaea、Periconia、Kiflimonium、Pseudorobil?lar?da、unidentified_Hypocreales等6屬及未知屬真菌僅在易結香品種HBQ中普遍存在,且豐度較高,具有一定的特異性。Hypomontagnella、Colle?tot?richum、Nigrograna等3屬在易結香品種JSZ中具有特異性,普遍存在且物種豐富較高。易結香品種XGY與其他4個品種相比較,沒有明顯豐度特異的菌群。
為了進一步探究不同分組間真菌類群豐度差異顯著性,本研究利用STAMP軟件對大于1%豐度屬進行Welch’s t-test檢驗。如圖6所示,差異分析結果表明,C1與C2組結香部位樣本菌群存在3種顯著差異的屬,包括Neurospora、Aspergillus和Hermatomyces。普通品種C2與易結香品種HBQ、JSZ、XGY間存在4種豐度顯著差異的屬,包括Neurospora、Fusarium、Roussoella和Hypomontagnella;品種XGY與JSZ之間存在2種顯著豐度差異的屬,包括Fusarium和 Hy-pomontagnella;品種XGY與HBQ之間僅有Fusarium存在顯著豐度差異;品種JSZ與HBQ之間僅有Hypomontagnella存在顯著豐度差異。
3 ?討論
本研究在以往研究的基礎上,以不同地區、不同品種白木香結香部位為研究對象,通過對真菌ITS1區域進行高通量測序分析,探究結香部位真菌物種多樣性差異,為真菌高效人工促香技術、加快沉香生產速度提供理論依據。高通量測序得到白木香結香部位樣本中共4門、18綱、61目、29科、189屬、231種真菌。
通過對東莞和茂名地區普通品種結香樣本中真菌物種多樣性進行比較發現:(1)相同環境下,同種白木香不同高度結香位點真菌類群豐度存在差異,但菌群結構極為相似,C1組不同樣本真菌組成結構相似,此結果與Liu等的結果一致;(2)不同區域環境中,同種白木香結香部位真菌群落之間差異非常顯著,區域生境差異可能是導致結香部位真菌群落結構差異的主要因素之一。C1與C2中,Neurospora、Aspergillus和Hermatomyces屬真菌的豐度在統計學上存在顯著差異,特別是Neurospora在C2組中為優勢物種,豐度占比達到61.2%。根據基因ITS序列鑒定,Neurospora屬真菌主要為Neurospora crassa Shear & B.O. Dodge。
Neruospora crass隸屬于子囊菌門、糞殼菌綱、糞殼菌目、糞殼菌科,該菌是天然的纖維素快速降解菌,可以快速降解木質纖維素,并具有利用木糖、纖維二糖、寡糖等組分生成乙醇及木糖醇的能力,是研究絲狀真菌纖維素酶表達分泌和木質纖維素降解機制的模式真菌[26-27],同時在工業生產上也具有重要的應用價值。該菌對纖維素的快速降解能力已經得到了廣泛的認可,其纖維素酶產率與著名的瑞氏木霉(Trichoderma)的纖維素酶產率相近[27-28]。這種降解能力預示了其對白木香受傷部位的刺激能力極強,可促進白木香的抗逆性反應并結香。以往并沒有該菌促進白木香結香的研究,本研究首次揭示了茂名普通品種結香部位中該菌的顯著優勢地位。目前,普通品種的白木香在廣東廣西等地方廣泛種植,該菌的進一步深入研究對人工結香的研究具有重要的意義。
對茂名地區4個不同品種白木香結香部位的真菌多樣性進行比較分析,揭示不同品種白木香結香部位真菌組成差異明顯,但Fusarium、Corynespora、Roussoella、Kockovaella、Acrocalymma、Hypomontagnella和Bannoa等7屬真菌在易結香品種中廣泛分布且具有較高的豐度,普通品種中分布較少且豐度極低(圖2,圖5)。特別是在易結香品種中,Fusarium屬真菌在XGY中為優勢屬,其豐度占比高達77.6%;Fusarium屬(Hypocreales目)及未鑒定的unidentified_ o_Hypocreales目真菌在HBQ中占比最高,合計占比50.8%(20.1%和30.7%);Hypomontagnella屬真菌為JSZ結香部位中的優勢真菌,豐度占比達52.4%。Fusarium和Hypomontagnella屬真菌的顯著優勢可能與易結香特性顯著相關。
Fusarium隸屬于子囊菌門、糞殼菌綱、肉座菌目、叢赤殼科,該屬已被證實是一類白木香內生真菌,主要分布在白木香腐木部分,特別是腐皮鐮刀菌Fusarium solani (Mart.) Sacc.,已有多項研究證實該真菌能夠與白木香形成互惠互利關系,實現與白木香的協同進化,同時該屬真菌的部分次級代謝產物還是沉香的特征性成分,如色酮類化合物[29-31]。使用該屬真菌刺激,可誘導白木香代謝更加旺盛,加速誘導白木香結香,油脂積累時間短且量大。
Hypomontagnella隸屬于子囊菌門、糞殼菌綱、炭角菌目、炭團菌科,基于形態學特征和系統發育分析,該屬自炭團菌屬(Hypoxylon)分離,于2018年被建立為新屬[32]。該屬與炭團菌屬親緣關系相近,是一種林木致病菌,可引起植物的潰瘍病,特別是Hubbes于1964年首次報道了炭團菌毒素的存在,在潰瘍病的發展中起到關鍵作用[33]。炭團菌毒素可以顯著抑制寄主病斑處瘤狀愈傷組織的形成,對植物受傷害后的影響非常強烈[34]。本研究首次在易結香品種JSZ結香部位中發現Hypomontagnella屬真菌,且該屬為顯著優勢物種,豐度占比達到52.4%,據此推斷,該屬真菌在白木香受傷結香的過程中發揮了重要的作用,Hypomontagnella屬真菌在白木香人工結香技術的開發中具有重要的開發潛力。
4 ?結論
真菌刺激已被證明是一種有效的方法促進白木香結香,多種真菌已被證明可以顯著促進沉香的形成,然而,生境差異、不同品種結香特性和真菌分布之間關系仍然是個未解的謎題。本研究結果表明:相同生境條件下,同種白木香不同高度結香位點真菌組成結構非常相似,但不同真菌的豐度存在差異;不同區域生境下,相同品種結香部位真菌群落組成結構存在顯著差異,生境差異可能是導致結香部位真菌群落結構差異的主要因素之一;同一區域生境下,不同品種白木香結香部位的真菌群落結構存在顯著差異,品種差異可能是導致白木香結香部位真菌群落結構差異的主要因素之一,同時特有真菌類群的分布也可能是影響白木香結香特性的關鍵。不同生境下白木香結香中發揮主要刺激作用的真菌并不相同;不同品種沉香形成過程中可能有不同的真菌在發揮主導作用。因此,在利用真菌刺激白木香結香的過程中,要因地制宜,根據不同的生境條件和白木香品種來選擇最適的真菌。
參考文獻
[1] 楊德蘭, 梅文莉, 楊錦玲, 等. GC-MS 分析4 種奇楠沉香中致香的倍半萜和2-(2-苯乙基)色酮類成分[J]. 熱帶作物學報, 2014, 35(6): 1235-1243.
[2] Liu J, Zhang X, Yang J, et al. Agarwood wound locations provide insight into the association between fungal diversity and volatile compounds in Aquilaria sinensis[J]. Royal Society Open Science, 2019, 6(7): 190211.
[3] Persoon G A, van Beek H H. Growing ‘the wood of the gods’: agarwood production in southeast Asia[M]//Snelder D J., Lasco R D. Smallholder tree growing for rural development and environmental services. Dordrecht: Springer, 2008: 245-262.
[4] Chen H Q, Yang J S, et al. Chemical constituents of agar-wood originating from the endemic genus Aquilaria plants[J]. Chemistry & Biodiversity, 2012, 9(2): 236-250.
[5] Naef R. The volatile and semi-volatile constituents of agar-wood, the infected heartwood of Aquilaria species: a re-view[J]. Flavour Fragrance Journal, 2011, 26(2): 73-87.
[6] Wu B, Kwon S W, Hwang G S, et al. Eight new 2- (2-phen-yl?e?thyl)chromone (=2-(2-phenylethyl)-4H-1-ben-zopyran-4- one) derivatives from Aquilaria malaccensis agarwood[J]. Helvetica Chimca Acta, 2012, 95(9): 1657-1665.
[7] 李月菲, 田從魁, 孟嘉星, 等. 沉香的化學成分及藥理作用研究進展[J]. 國際藥學研究雜志, 2019, 46(7): 498-506.
[8] 劉洋洋, 楊 ?云, 林 ?波, 等. 四批通體香沉香藥材的揮發油成分分析[J]. 化學與生物工程, 2014, 31(5): 67-71.
[9] Xu Y H, Zhang Z, Wang M X, et al. Identification of genes related to agarwood formation: transcriptome analysis of healthy and wounded tissues of Aquilaria sinensis[J]. BMC Genomics, 2013, 14(1): 227.
[10] Ye W, He X, We H Q, et al. Identification and characteriza-tion of a novel sesquiterpene synthase from Aquilaria sinensis: an important gene for agarwood formation[J]. International Journal of Biological Macromolecules, 2018, 108: 884-892.
[11] Chhipa H, Chowdhary K, Kaushik N. Artificial production of agarwood oil in Aquilaria sp. by fungi: a review[J]. Phy-tochemistry Reviews, 2017, 16(5): 835-860.
[12] Wilson J J, Kitto M. Aquilaria malaccensis Lam., a red-listed and highly exploited tree species in the Assamese home garden[J]. Current Science, 2012, 102(4): 546-547.
[13] 戚樹源, 林立東, 葉勤法. 沉香中芐基丙酮及其在黃綠墨耳真菌中的生物轉化[J]. 生物工程學報, 1998, 14(4): 464-466.
[14] Subehan S, Ueda J, Fujino H, et al. A field survey of agar-wood in Indonesia [J]. Journal of Traditional Medicines, 2005, 22(4): 244-251.
[15] Tamuli P, Boruah P, Nath S C, et al. Fungi from disease agarwood tree (Aquilaria agallocha Roxb): two new records [J]. Advances in Forestry Research in India, 2000, 22: 182-187.
[16] 徐維娜. 真菌侵染誘導沉香形成關鍵技術效果評價及結香機制初步研究[D]. 廣州: 廣東藥學院, 2011.
[17] Chen X Y, Chun S, Liu Y Y, et al. Agarwood Formation Induced by Fermentation Liquid of Lasiodiplodia theobro-mae, the Dominating Fungus in Wounded Wood of Aquilaria sinensis[J]. Current Microbiology, 2017, 74(4): 460-468.
[18] 張苗苗. 白木香中內生菌的分離鑒定及其分布規律的初步研究[D]. 廣州: 廣州中醫藥大學, 2017.
[19] 谷 ?田, 彭海剛. 奇楠沉香揮發油化學成分分析[J]. 廣東化工, 2012, 39(4): 257-258.
[20] 李 ?薇, 梅文莉, 董文化, 等. 國產綠奇楠沉香的化學成分研究[J]. 熱帶亞熱帶植物學報, 2019, 27(2): 196-202.
[21] 張巧葉. 奇楠沉香品種及其品質分析[J]. 南方農業, 2019, 13(32): 142-157.
[22] 李 ?薇, 梅文莉, 董文化, 等. 國產綠奇楠沉香的化學成分研究[J]. 熱帶亞熱帶植物學報, 2019, 27(2): 196-202.
[23] 梅文莉, 楊德蘭, 左文健, 等. 奇楠沉香中2-(2-苯乙基)色酮的GC-MS 分析鑒定[J]. 熱帶作物學報, 2013, 34(9): 1 819-1 824.
[24] Bolyen E, Rideout J R, Dillon M R, et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2[J]. Nature Biotechnology, 2019, 37: 852-857.
[25] Parks D H, Tyson G W, Hugenholtz P, et al. STAMP: Statistical analysis of taxonomic and functional profiles[J]. Bioinformatics, 2014, 30(21): 3 123-3 124.
[26] 張 ?瀟, 朱冬青, 王 ?丹, 等. 粗糙脈孢菌( Neurospora crassa) 木糖發酵的研究[J]. 微生物學報, 2003, 43(4): 466-472.
[27] 馮 ?炘, 王 ?丹, 辛麗霞, 等. 粗糙脈孢菌( Neurospora crassa) 產纖維素酶發酵條件研究[J]. 食品科學, 2005, 26(1): 67-70.
[28] 遲乃玉, 張慶芳, 劉長江, 等. 纖維素酶高產菌株最適發酵條件的研究[J]. 沈陽農業大學學報, 2000, 31(4): 380-382.
[29] 李雯珊. 白木香內生真菌Fusarium solani A2和Fimetariella rabenhorstii A20次級代謝產物研究[D]. 廣州: 廣東藥科大學, 2016.
[30] 王 ?磊, 章衛民, 潘清靈, 等. 白木香內生真菌的分離及分子鑒定[J]. 菌物研究, 2009, 7(1): 37-42.
[31] 高曉霞, 周偉平, 王 ?磊, 等. 鐮刀菌誘導結香對白木香葉內生真菌分布和群落構成的影響研究[J]. 中國中藥雜志, 2014, 39(2): 197-203.
[32] Lambert C, Wendt L, Hladki A I, et al. Hypomontagnella (Hypoxylaceae): a new genus segregated from Hypoxylon by a polyphasic taxonomic approach[J]. Mycological Progress, 2019, 18(1): 187-201.
[33] Hubbes M. The incasion site of Hypoxylon pruinatum on Populus tremuloides[J]. Phytopathology, 1964, 54: 896.
[34] Bagga D K, Smalley E B. Factors affecting inoculated with Hypoxylon pruinatum[J]. Canadian Journal of Botany, 1969, 47: 907-914.
責任編輯:崔麗虹
