番茄果實成熟軟化過程中細胞壁作用機制研究進展
2022-01-07 02:28:42楊淼焱
保鮮與加工 2021年12期
敖 雁,楊淼焱,張 馳,吳 啟,*
(1.蘇州健雄職業技術學院,江蘇 蘇州 215411;2.東京大學農學生命科學研究科,東京 113-8657;3.雪城大學,雪城 13244)
番茄(Solanum lycopersicum)果實可為人類提供豐富的營養物質、水分、蛋白質及維生素,但其果實成熟后極易軟化,造成腐爛變質。因此番茄果實軟化機制及延長其保鮮時間一直是科學家研究的重點課題。作為研究肉質植物果實成熟軟化機制的模式作物[1],番茄果實成熟軟化的過程十分復雜,包括一系列的生理生化反應,如細胞壁的降解、呼吸速率以及其他代謝產物的變化等。
植物細胞壁由多種生物復合體組成,是自然界最為復雜的細胞結構之一。事實證明,成千上萬的基因參與這一過程。然而,植物細胞壁卻仍是植物中了解最少的細胞結構[2]。番茄細胞壁主要是由果膠、纖維素、半纖維素和糖蛋白等相互交錯形成的復雜網絡結構[3],而細胞壁的水解作用更是造成番茄果實軟化的重要影響因素,是一個高度有組織的過程,涉及各種細胞壁水解酶之間的復雜相互作用[4-5]。本文綜述了番茄成熟軟化過程中的細胞壁果膠、纖維素或半纖維素、多糖降解相關基因和所編碼的水解酶、伸展蛋白及其受上游調控的轉錄因子之間的主要作用機制,并在此基礎上總結了上述影響因素在造成番茄果實軟化這一過程中的相互關系。
1 番茄細胞壁果膠相關基因及其所編碼的酶
果膠通常由原果膠、水溶性果膠和果膠酸組成。番茄果實成熟過程中,原果膠分解成溶解性果膠,最終降解為半乳糖醛酸,這一過程需要多聚半乳糖醛酸酶(Polygalacturonase,PG)、果膠甲酯酶(Pectin pectylhydrolase, PME)、β-半乳糖苷酶(β-galactosidase, β-GAL)以及多果膠裂解酶(Pectate lyases, PL)等許多與果膠解聚相關的酶共同參與。
1.1 多聚半乳糖醛酸酶(PG)
多聚半乳糖醛酸酶主要通過作用于1,4-2-D-半乳糖苷鍵,催化果膠分子裂解生成低聚半乳糖醛酸或半乳糖醛酸,使細胞壁發生解體,從而導致果實變軟。早在1988 年,Bird 等[6]就發現編碼蛋白區域的序列是一段PG cDNA,并證明了每個基因組只含有一個編碼PG 的基因。利用番茄成熟突變體alc、nor 和rin 為研究材料,研究人員發現果實成熟過程中PG 活性很低或不含PG 活性,而突變體材料的果實細胞壁層次清晰可見,胞間層結構緊密、均勻而連續,胞壁纖維排列整齊,這進一步說明PG 可以通過改變細胞壁結構來影響果實軟化的過程[7]。近年來在番茄基因組中已經鑒定出54 個PG 基因[8],本文整合了番茄不同PGs表達相關基因及研究進展(表1),關于不同PG 基因在細胞壁果膠裂解過程中的分子機理仍有待進一步探究。
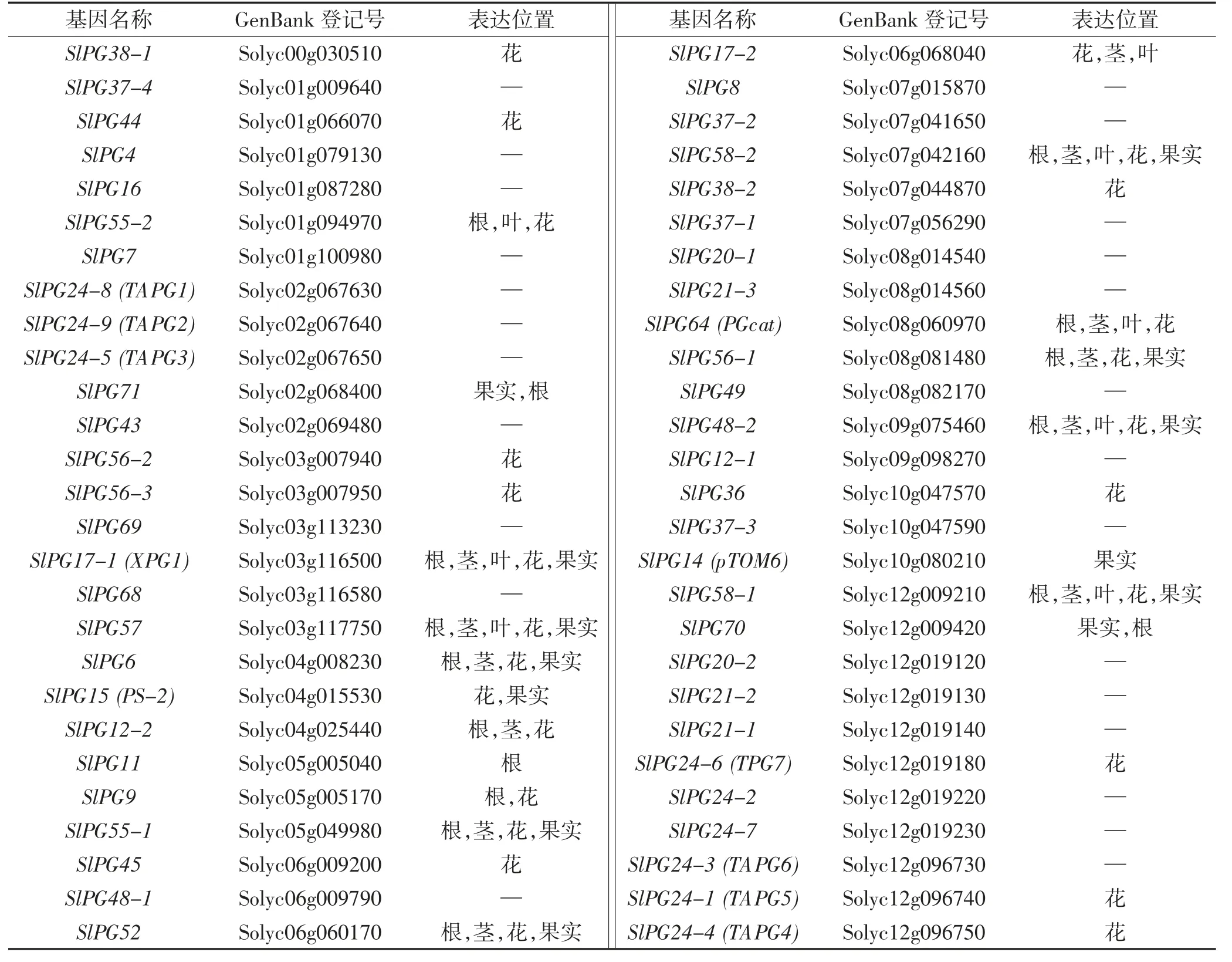
表1 番茄PGs 相關基因及其表達位置Table 1 PGs related genes and the expression positions in tomatoes
除了基礎研究領域,PG 基因也被廣泛應用到番茄的生產環節。陳麗萍等[9]采用傳統的根瘤農桿菌介導法將PG 反義基因導入加工番茄品種B04中,抑制轉基因番茄中內源PG 的活性,解決了番茄在貯存與運輸過程中腐爛變質的問題。
1.2 果膠甲酯酶(PME)
果膠甲酯酶通過作用于果膠分子鏈上半乳糖醛酸羧基上的酯化基團,使內源果膠在原位脫去甲基,果膠水溶度增大,生成便于與PG 作用的底物,進而使番茄果實軟化[10-11]。2001 年,Micheli[12]證實了植物體中果膠甲酯酶可以催化細胞壁的PG 去甲基化作用,但是與PG 基因相比,這種去甲基化作用所產生的效果又是極其不明顯的[13]。也有研究認為PME 通過激活早期成熟過程中PG 的活性來參與果實軟化過程[14],但其活性與番茄果實硬度及細胞壁組分無顯著相關[15]。近年來,研究人員從番茄果皮組織中分離得到兩種主要的PME 亞型(PME1 和PME2)并分別成功獲得它們的突變體以及雙突變體材料。研究表明,PME2 對果膠的去酯化反應有很大影響[16]。與此同時,Reca 等[17]采用功能基因組的方法從番茄中鑒定了一個果膠甲酯酶抑制蛋白并命名為SolyPMEI,可以和PME-1 互相作用來共同調節甲酯化果膠在果實成熟過程中的時空分布。
1.3 β-半乳糖苷酶(β-Gal)
在番茄果實成熟過程中,β-半乳糖苷酶(β-Gal)能夠將果膠半乳糖水解為葡萄糖和半乳糖而參與細胞壁代謝[18-19]。番茄果實中存在3 種β-半乳糖苷酶的同工酶,即β-Gal Ⅰ/Ⅱ/Ⅲ,但僅有β-Gal Ⅱ與果實軟化相關,可催化β-(l, 4)-半乳糖苷鍵發生降解。Brummell 等[4]研究發現了半乳糖代謝與果實成熟有一定關系。據報道,番茄基因組中存在17 個β-Gal 基因(表2),所有被檢測的β-Gal 酶都具有β-Gal 活性。采用RNAi 技術獲得β-Gal4 突變體分析發現β-Gal4 基因參與番茄果實軟化過程,但大多數β-Gals 功能仍有待鑒定[20-21]。此外,后續的轉基因實驗也證實β-半乳糖苷酶基因DKGAL1 在果實成熟過程中可以通過改變番茄果實中半乳糖代謝來減少細胞間的粘附力,促進果實的軟化[18]。

表2 番茄TBGs 相關基因及其表達位置Table 2 TBGs related genes and the expression positions in tomatoes
1.4 果膠裂解酶(PL)
果膠裂解酶主要依靠裂解果膠β-1,4-半乳糖醛酸基團從而達到細胞壁解體、細胞分離及果實軟化的目的。Yang 等[25]在番茄中鑒定了22 個同源的果膠裂解酶基因,其中SlPL(Solyc03g111690)在果實成熟階段大量表達。利用RNAi 干擾技術獲得了SlPL 沉默株系,SlPL 基因特異性沉默可以有效增強果實的硬度、降低軟化速率并延長番茄果實貨架期,同時使果實中果膠合成酶、木葡聚糖內糖基轉移酶、伸展蛋白等表達量顯著降低。
1.5 鼠李糖醛酸裂解酶(RGL)
鼠李糖苷I(RG-I)是植物細胞壁果膠的一個結構域,鼠李糖醛酸裂解酶通過裂解位于主鏈l-鼠李糖與d-半乳糖醛酸殘基之間的α-1,4 糖苷鍵來降解RG-I。為了研究RGL 在番茄果實軟化中的作用,Ochoa-Jiménez 等[26]和Berumen-Varela 等[27]在番茄35S 啟動子下表達了solyc11g011300 基因,發現轉基因番茄果實表現出更強的硬度、更長的貨架期和更少的枯萎量,同時轉基因番茄果實的成熟時間延遲了一周。
2 番茄細胞壁纖維素或半纖維素相關基因及其編碼的酶
2.1 木葡聚糖內糖基轉移酶/水解酶(Xyloglucan endotransglycosylase,XET/XTH)
木葡聚糖是半纖維素多聚糖的重要組分之一,與半纖維素及纖維素微纖絲協同構成網狀結構,是細胞壁的主要機械支撐物[28]。Maclachlan 等[29]和Sakurai 等[30]通過比較番茄果實成熟突變體rin 與野生型果實中的木葡聚糖分子量的變化,發現果實的軟化可能與半纖維素木葡聚糖的降解有關。木葡聚糖內糖基轉移/水解酶(XET/XTH)是木葡聚糖在細胞壁解聚和再連接的關鍵酶,具有催化木葡聚糖分子水解和糖基轉移的雙重作用,它在番茄果實成熟軟化過程中發揮重要功能。XET 通過水解木葡聚糖β-1-4 糖苷鍵的主鏈,將新生成的降解末端轉移至其他木葡聚糖多聚體的C 端[31]。同時,細胞壁還能通過XTHs 編碼的木葡聚糖水解酶XET 的活性來催化木葡聚糖的水解[32]。Saladié等[33]在番茄中發現了25 個XTHs 基因,本文整合了番茄不同XTHs 相關基因及研究進展(表3)。研究表明,乙烯處理也能激活XET 的活性,尤其是SlXTH5和SlXTH8 能受到乙烯的顯著誘導,協同通過影響細胞壁水解的過程來參與對番茄果實成熟的調控[34],但其具體的分子調節機理仍有待進一步探討。
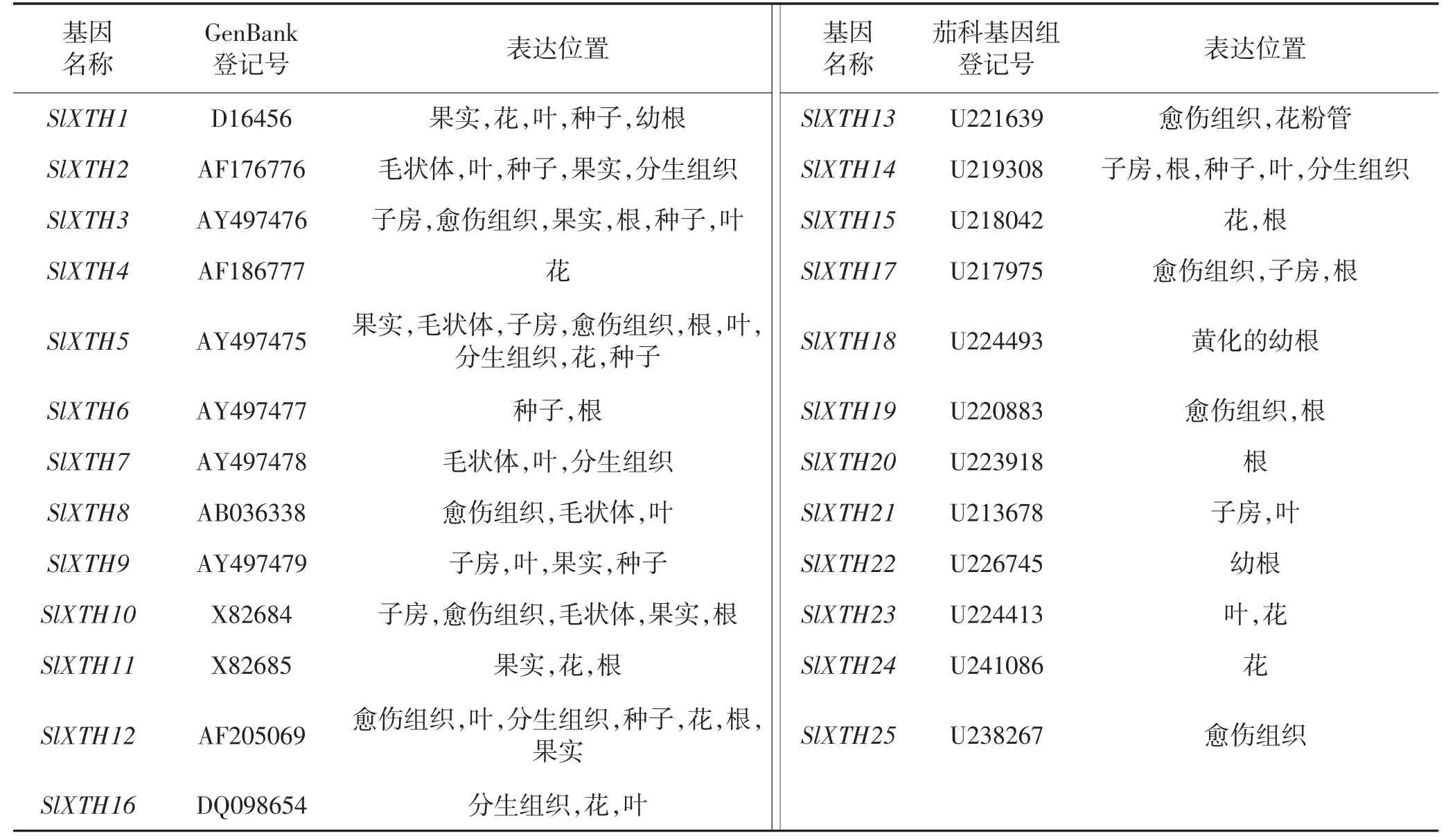
表3 番茄不同XTHs 相關基因及其表達位置Table 3 XTHs related genes and the expression positions in tomatoes
2.2 內切-1,4-β-D-葡聚糖酶(Endo-1,4-β-Dglucanase,EG)
EG 是纖維素酶3 種主要組分之一,對木葡聚糖、羧甲基纖維素以及具有葡聚糖結構的物質表現出活性。在番茄中目前一共發現7 個編碼EG 的基因。其中隨著番茄果實的成熟,LeCel1 和LeCel2 在mRNA水平上逐漸積累,且能夠受到外源乙烯的誘導,說明其轉錄表達可以受到植物激素乙烯的調控[4]。除此之外,研究表明LeCel1 和LeCel2 蛋白易于在番茄葉片中高度積累,并在病原菌抗性方面發揮著重要作用[35]。而在其他物種中,研究還發現EG 活性與白梨果實成熟密切相關,PbEG3 在果實成熟和軟化過程中可能發揮著更為重要的作用[36]。
3 番茄細胞壁多糖降解相關基因及其編碼的酶
甘露聚糖內切酶是一種多糖降解酶,主要水解甘露聚糖之間的β-1,4 共價鍵,甘露聚糖酶被認為通過參與細胞壁的降解在番茄果實成熟過程中發揮作用[37-38]。1997 年,Huysamer 等首先發現了番茄果實中存在甘露聚糖,但甘露聚糖在番茄果實細胞壁中比例并不高。隨后不久,甘露聚糖酶活性也在番茄果實中被進一步證實,且在番茄品種Trust 中,甘露聚糖酶活性可以隨著果實成熟而增加[39]。截至目前,研究人員已經鑒定出番茄果實中的甘露聚糖酶基因并命名為LeMan4 基因[40]。為了進一步探究LeMan4 基因在細胞壁降解和果實軟化中所發揮的功能,Wang 等[41]分別構建了GUS 和GFP 轉基因株系,發現Leman4a 主要定位在細胞壁,且在果實果皮和外果皮中高度富集。然后利用RNAi 和反義RNA 干擾技術,研究者獲得了突變體材料,分析發現突變體果實硬度與野生型材料相比略有降低,因此分析認為,Leman4a 基因確實參與果實成熟過程,但不足以引起細胞壁的水解,推測可能還與其他已知的細胞壁修飾酶起協同作用共同參與細胞壁重排的過程。赫崇慧[42]發現在番茄果肉中甘露聚糖酶與PG、PME 和纖維素酶存在著一定的協同作用關系,并且這種協同關系影響著果肉的成熟軟化過程。
4 伸展蛋白(Expansin proteins,Exp)
伸展蛋白通過打斷細胞壁纖維素和半纖維素之間的共價鍵,使它們之間發生位移,最終引起細胞壁松弛[43]。雙子葉植物的Exp 是一個小分子蛋白,由25個基因構成的基因家族編碼。Brummell 等[44-45]認為在番茄中,擴展基因LeExp1 誘導果實成熟相關基因的mRNA 和蛋白質積累,LeExp1 突變體果實在整個成熟過程中明顯比對照更堅硬。此外,研究發現SlExp1的碳水化合物結合基序(Carbohydrate binding module,CBM)對植株和果實表型都有影響,SlExp1-CBM 轉基因植株表現出延遲軟化和結實的表型[46]。大多數情況下,Exp 蛋白被認為可以影響番茄果實成熟早期引起的細胞壁變化的某些酶的活性變化,特別是木葡聚糖內糖基轉移/水解酶和β-半乳糖苷酶的活性等[19]。例如Minoia 等[47]發現番茄突變體中SlExp1 基因功能的缺失可以通過影響XET/XTH 活性來改變纖維素的結構,最終延緩果實的衰老。進一步對SlExp1-7 突變體植株的細胞壁組分分析,發現木葡聚糖結構發生了較大的變化。
5 轉錄因子
隨著分子生物學技術的不斷發展,一些調控細胞壁水解的轉錄因子也陸續被發現。GRAS 家族蛋白是植物所特有的轉錄因子,Zhang 等[48]進一步研究發現一種新的GRAS 基因,命名為SlFSR (Fruit shelf-life regulator)。在果實成熟過程中SlFSR 的表達顯著增加,但在番茄成熟突變體rin 中顯著降低。同時SlFSR的表達受到外源乙烯的誘導,推測SlFSR 基因可能位于乙烯信號的下游發揮作用。RNAi 抑制SlFSR 基因后導致多個細胞壁修飾相關基因表達量減少,PG、TBG 和CEL 活性明顯降低,果實貨架期顯著延長。此外,rin 中SlFSR 基因過表達導致PG、TBG4、CEL2、PL、PME、MAN1、EXP1 和XTH5 等多個細胞壁修飾相關基因的表達上調,顯著縮短了果實的貨架期,表明SlFSR 基因是控制番茄果實貨架期的另一個潛在的生物技術靶點。
目前在番茄中共發現101 個不同的NAC 轉錄因子,其中SlNAC1 和SlNAC4 被證明可以參與調控番茄果實成熟的過程。SlNAC1 轉錄因子能與乙烯合成基因PSY1、ACS2、ACO1 的啟動子特異性結合,通過在轉錄水平上調控番茄乙烯的生物合成過程,進而影響果實軟化[49]。相反,在SlNAC1-RNAi 轉基因株系中,果實軟化過程受到明顯抑制并且伴隨著乙烯生物合成相關基因的大量誘導[50]。同樣,利用RNAi 技術沉默SlNAC4 的表達后,番茄果實成熟滯后,體內乙烯含量也急劇減少,說明SlNAC4 也可以通過乙烯合成的途徑來調控番茄果實軟化的過程[51]。此外,最近一個新型的NAC 轉錄因子NOR-like1 也被發現可以參與番茄果實的調節過程。RNA 測序結果顯示NOR-like1 不僅能夠影響乙烯的生物學合成(SlACS2,SlACS4),還能夠通過改變細胞壁代謝相關酶PG、PL、CEL2、EXP1 的活性來參與番茄成熟的過程[52]。RIN 轉錄因子是MADS-box 家族成員之一,主要控制番茄果實成熟。近年來通過蛋白質組學、電泳遷移率分析(EMSA)和染色質免疫共沉淀(ChIP)等分析手段鑒定了一系列新的RIN 靶基因。研究表明,LeACO4 和α-Gal 基因的啟動子區域檢測到RIN 蛋白特定結合位點CArG 基序,表明RIN 可能直接調控它們的轉錄,這些結果進一步證明了RIN 在番茄果實成熟軟化尤其是在細胞壁降解過程中所發揮的作用[53]。
6 小結與展望
在番茄果實成熟軟化中,多種細胞壁相關基因、酶和轉錄因子共同起作用,王倩等[54]研究了不同耐貯性的番茄品種,發現隨著貯藏時間的延長,與普通的“津粉207”番茄品種相比,耐貯性強的“歐盾”品種多聚半乳糖酶、果膠酯酶、β-半乳糖苷酶和纖維素酶等相關酶活性明顯偏低,乙烯釋放量低,番茄果實硬度下降較慢。該結果進一步驗證了乙烯與細胞壁水解酶相互交織,共同影響番茄正常的果實衰老過程。綜上結論,本文繪制了番茄果實成熟過程中幾種常見轉錄因子通過調節乙烯合成途徑,最終影響細胞壁代謝的調控網絡(圖1)。
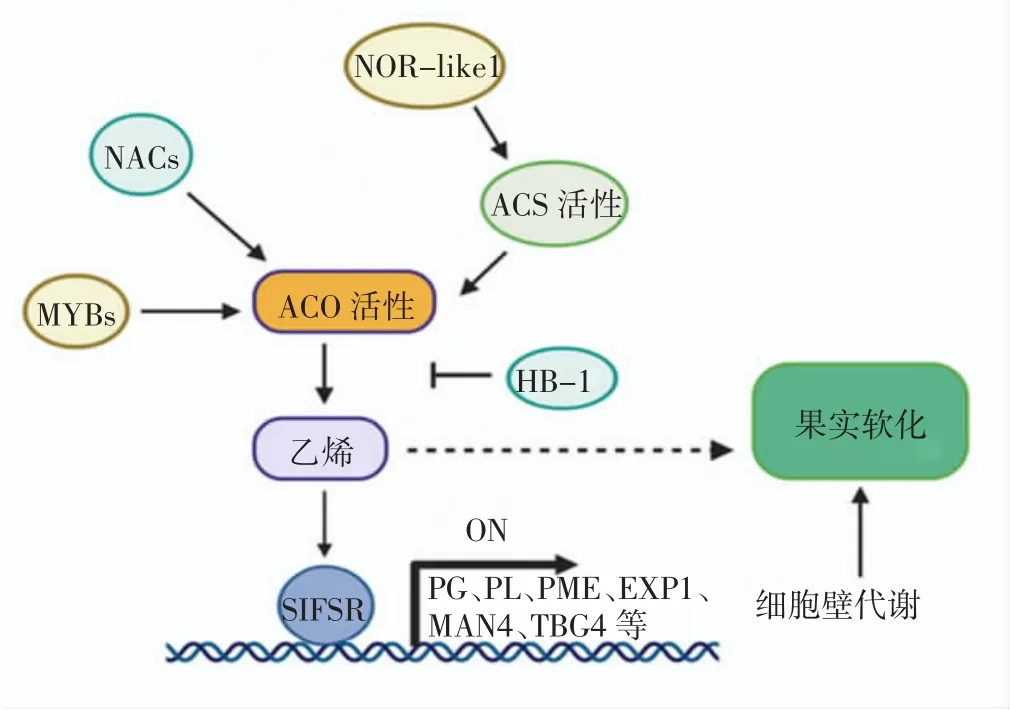
圖1 轉錄因子、乙烯和細胞壁水解酶調控果實成熟軟化機制Fig.1 Regulatory mechanism of transcription factors,ethylene and cell wall hydrolase in tomato fruit softening
從圖1 可以看出,乙烯(Ethylene)的合成影響番茄果實軟化,兩個主要酶ACO(ACC 氧化酶)和ACS(ACC 合成酶)影響著乙烯的合成,轉錄因子MYBs和NACs 影響ACO 的活性,轉錄因子NOR-like1 通過影響ACS 的活性而影響ACO 的活性。乙烯通過影響轉錄因子SIFSR 而進一步激活細胞壁中PG、PL、PME、EXP1、MAN4 和TBG4 等酶的活性,從而影響細胞壁的代謝,最終影響果實軟化。
近年來隨著基因編輯手段在水稻、擬南芥等模式作物中的大規模應用,番茄遺傳材料的獲得也越來越便捷有效。然而,目前針對細胞壁水解酶的研究仍多集中在單個水解酶或者某一類水解酶家族在番茄果實成熟中所起的作用,細胞壁水解酶之間存在的相互關聯,植物激素以及轉錄因子之間的調控網絡有待進一步完善。此外,目前影響番茄果實成熟的細胞壁水解酶相關基因的研究仍集中在同源基因功能的驗證,采用正向遺傳學的方法挖掘細胞壁水解過程中的新基因也應該是科研工作者們關注的重點內容。與此同時,針對不同番茄品種、不同種植地域、同一番茄品種的不同生育期,控制其果實軟化的關鍵水解酶也不盡相同,因此在農業生產應用上還需注意因時因地制宜,其更深層次的調控機理還需進一步研究。
