植物分枝性研究進展及其在蔬菜育種中的應用
2022-02-13 12:57:20夏雨桐
中國蔬菜 2022年1期
夏雨桐 王 琛 郝 寧 武 濤*
(1 湖南農業大學園藝學院,園藝作物種質創新與新品種選育教育部工程研究中心,蔬菜生物學湖南省重點實驗室,湖南長沙 410128;2 東北農業大學園藝園林學院,黑龍江哈爾濱 150030)
分枝是植物株型形態建成的重要農藝性狀之一。分枝可影響作物產量,同時分枝狀況也會影響作物的栽培密度和營養分配。較多的分枝會導致通風不良,極易使作物發生病害造成減產。此外,分枝也是觀賞植物一個重要的株型性狀,分枝的生長狀態可影響觀賞植物的造景效果。近些年,分枝逐漸成為育種家們較為關注的作物地上部性狀之一。
植物分枝一般分為二叉分枝、假二叉分枝、合軸分枝、單軸分枝和分蘗等。單軸分枝的主要代表植物有擬南芥和黃瓜等;合軸分枝的主要代表植物是番茄;分蘗的主要代表植物為大多數禾本科植物,例如小麥和水稻;假二叉分枝的主要代表作物有茄子;而辣椒一般屬于二叉分枝或者三叉分枝(鞏鵬濤和李迪,2005)。
1 植物分枝的環境調控
植物的分枝受光照、溫度、營養狀態等環境因素的影響(Snowden &Napoli,2003),其中光照對植物分枝具有重要影響。研究表明,高強光可通過促進越桔側芽生長從而促進分枝(Kawamura &Takeda,2002)。低光量子通量密度會減少擬南芥分枝(Su et al.,2011)。紅光與遠紅光的比率降低時,分枝抑制因子()表達升高從而導致擬南芥分枝減少(Ballare,1999;Franklin &Whitelam,2005;Gonzalez-Grandio et al.,2013)。光合面積減少可促進分枝抑制基因的表達進而減少玉米的分蘗(Kebrom &Mullet,2015)。超過一定范圍的光密度,會導致番茄中抑制分枝的激素獨腳金內酯(strigolactones,SLs)含量增加和SLs 合成基因()的表達升高,從而抑制番茄分枝(Koltai et al.,2011)。此外,亮/暗(7 h/7 h)過渡頻率的增加會抑制擬南芥腋芽的發育從而減少分枝(Jouve et al.,1998a,1998b)。溫度也是影響分枝的重要因素之一,低溫會誘導月季中的SLs 信號基因()的表達,促使其在莖節形成梯度進而影響側枝的萌發率(Djennane et al.,2014)。低溫也會促進SLs 受體基因()的轉錄從而抑制水稻的分蘗(Chen et al.,2018)。營養狀態也可調控植物的分枝發育。研究表明,糖類的輸入和輸出對桃樹腋芽的萌發有重要作用(Karine et al.,2004)。豌豆莖尖通過限制轉移到腋芽的糖量來限制腋芽的生長(Michael et al.,2014)。缺糖導致了小麥突變體的少分蘗表型(Kebrom et al.,2012)。異麥芽酮糖可以促進芽的萌發從而促進分枝,蔗糖、果糖、帕拉金糖和果膠都能促進離體培養的玫瑰側芽在光照下的萌發(Rabot et al.,2012)。磷和氮通過影響植物激素的合成及運輸來調控擬南芥側枝的生長發育(Evers et al.,2011)。缺磷通過抑制菊花腋芽的伸長來減少分枝長度,使其株型緊湊(郗琳,2015)。研究表明,氮肥對水稻的分蘗有促進作用(Li et al.,2016)。低氮處理可降低擬南芥中多分枝雙突變體-/-的分枝數量(Madeleine et al.,2017)。
2 植物分枝的激素調控
植物激素是打破側芽休眠和分枝持續生長必不可少的因素。外界環境的改變可使植物激素發生相應的變化,促使植物體內激素達到動態平衡,進而激發調控植物分枝基因的表達。
2.1 生長素
最早發現作用于植物分枝的激素是生長素(auxin,IAA)。提高植物體內IAA 水平后,側枝數目會明顯降低(Hall &Hillman,1975;Morris,1977;Greg et al.,1991;Romano et al.,1993)。研究表明,黃瓜莖尖中IAA 不能獨立抑制腋芽的生長,需要與其他抑制腋芽生長的物質共同作用(徐慶華 等,2011)。對去除穎花的水稻噴施IAA后,水稻分蘗伸長受到抑制(Arite et al.,2007)。研究發現,IAA 含量處于最低值是腋生分生組織(AM)起始的關鍵(Gallavotti,2013)。IAA 合成途徑(丙酮酸途徑)中2 個重要基因()和()的突變體合成IAA 的能力不強,導致頂端優勢受到抑制進而促進擬南芥分枝(Cheng et al.,2006)。玉米的同源基因功能缺失會導致AM 起始和側部器官發育發生缺陷,從而減少分枝(Phillips et al.,2011)。黃瓜在馴化過程中側枝抑制基因的表達量逐漸增高,在側枝中直接抑制IAA 輸出基因的活性,導致側枝中IAA 過量積累,從而抑制黃瓜側枝的生長發育(Shen et al.,2019)。IAA 信號轉導關鍵基因、、的突變或缺失通過影響IAA 的信號轉導進而促進番茄側枝生長(Chaabouni et al.,2009;Deng et al.,2012)。
2.2 細胞分裂素
細胞分裂素(cytokinin,CTK)可通過促進細胞分裂來活化側芽,促進植物分枝。直接對植物側芽噴施CTK 可抑制頂端優勢使側芽生長。研究發現,擬南芥、等多分枝突變體中的CTK含量較高,說明CTK 正調控植物分枝(Catterou et al.,2002;Zubko et al.,2002)。對擬南芥基部施加CTK 可減弱頂端優勢,促進側芽生長(Chatfield et al.,2000)。CTK 還可以促進AM 的起始和側芽的形成,擬南芥()多分枝突變體中由于CTK 水平增加導致蓮座葉和莖生葉的葉腋形成多個AM,增加分枝的數量,同時也可以促進側枝的伸長(Tantikanjana et al.,2001)。CTK 合成酶關鍵基因()高表達可促進CTK 對側芽的活化作用,進而促進擬南芥的分枝(Guo &Gan,2011)。
2.3 獨腳金內酯
SLs 的出現為研究植物腋芽的生長發育機制提供了一種新的思路(Rameau,2010)。有研究表明,SLs 可以作為一種長距離信號物質抑制側芽的生長發育,并且作為一種嫁接轉移物質調控TCP 家族的分枝抑制轉錄因子BRC1、TEOSINTE BRANCHED 1(TB1)、FINE CULM 1(FC1)的表達,從而抑制側芽伸長(Antonio et al.,2007;Shinohara et al.,2017)。SLs 可通過下調豌豆腋芽內的CTK 水平來抑制側芽生長(Victoria et al.,2008)。研究發現,SLs 可抑制打頂豌豆的側枝伸長從而抑制植物分枝(Dun et al.,2009)。在擬南芥中SLs 可通過減少IAA 輸出載體蛋白PIN 的數量來降低IAA 極性運輸的能力,從而抑制側枝生長(Ongaro &Leyser,2007)。隨著對SLs 的研究越來越深入,從生物合成到激素運輸再到信號轉導,SLs 調控植物分枝的機理也越來越清晰。SLs受體復合物D14-MAX2 的底物D53-like-SMXLs和BES1 相互作用,抑制的表達,從而促進擬南芥分枝(Hu et al.,2020)。SLs 的信號通路中有3 個抑制因子,分別是、、,研究發現在沒有SLs 的情況下SMXL6 +TPL 蛋白可直接與、、的啟動子結合并抑制其表達來增加SLs 的含量,從而抑制擬南芥分枝。同時,SMXL6 還可以與未知的轉錄因子形成復合物,這些轉錄因子有望識別并結合、或的啟動子,從而抑制它們的轉錄來促進擬南芥分枝(Wang et al.,2020)。
2.4 其他激素
研究發現,赤霉素(gibberellin,GA)、脫落酸(abscisic acid,ABA)和油菜素內酯(brassinosteroids,BRs)都可以調節植物分枝。水稻()基因可抑制分蘗,高水平的GA 可以激活APC/C(E3 泛素連接酶復合物),促進AM 的MONOCULM 1(MOC1)降解,從而抑制水稻分蘗(Lin et al.,2020)。高濃度ABA 會抑制擬南芥側芽的萌發從而抑制分枝(Pei et al.,1998)。BRs 信號通路調控基因會抑制擬南芥AM 起始關鍵基因-()的表達,從而抑制AM 啟始,抑制分枝(Gendron et al.,2012)。為獲得影響植物分枝的激素類型,對相關研究文獻進行了總結(表1)。

表1 影響植物分枝的激素類型
3 植物分枝的基因調控
影響植物分枝除了環境因素和植物激素外,還有遺傳因子。植物的分枝主要是由AM 起始,發育成腋芽,再由腋芽發育成側枝。所以其內在調控基因分為3 種:一是側芽起始發育調控基因;二是側芽伸長發育調控基因;三是側芽起始和伸長發育協同調控基因。
3.1 側芽起始發育調控基因
植物側芽的起始主要受到基因的調控。()屬于WRKY 轉錄因子家族,可促進AM 的啟動,產生較多的側芽來促進擬南芥分枝(于延沖,2011;Guo et al.,2015)。-(-)屬于MYB 轉錄因子家族,正調控擬南芥分枝(Muller et al.,2006)。在的上游,正調控的轉錄促進AM 起始,從而促進擬南芥分枝(于延沖,2011;Guo et al.,2015)。番茄中基因是的同源基因,參與AM 起始(Schmitz et al.,2002)。NAC 轉錄因子CUC 參與擬南芥AM 啟動(Stirnberg et al.,1999;Raman et al.,2008;宋玉光 等,2016),在擬南芥AM 形成中發揮重要作用(Hibara et al.,2006)。研究發現,CUC2 和CUC3 直接與DA1(其肽酶底物UBP15 可抑制AM 起始)啟動子結合并激活其表達,形成CUC2/CUC3-DA1-UBP15 調控模塊來控制擬南芥AM 的啟動(Li et al.,2020)。()屬于GRAS 轉錄因子家族,促進AM 形成,正調控擬南芥分枝(Greb et al.,2003)。和分別是在水稻和番茄中的同源基因,正調控AM 起始(Groot et al.,1994;Li et al.,2003)。研究表明,在AM 的啟動和建立過程中,和在擬南芥()的上游起作用(Matsuoka et al.,1993;Long et al.,1996;Li et al.,2003;Liang et al.,2014)。()屬于HDZIP 轉錄因子家族,作用于擬南芥AM 起始(Talbert et al.,1995)。可上調基因表達來啟動擬南芥AM(Otsuga et al.,2001)。屬于KNOX轉錄因子家族,通過調控AM 細胞的分化來促進擬南芥AM 形成(Williams &Fletcher,2005;Scofield &Murray,2006)。()基因通過維持干細胞的活性和調控下游基因促進腋芽原基形成;-(-)基因主要功能是促進擬南芥AM 細胞分化,STM 蛋白結合在啟動子上,上調其表達,同時受到的調控從而影響擬南芥腋芽原基形成(李亞棟 等,2009;周 超,2015)。()和()均 屬 于bHLH 轉 錄 因子家族,調控AM 形成,在水稻和突變體中都會產生較少的AM 和分蘗(張彩霞,2014)。玉米中的()基因和擬南芥中()基因為的同源基因,通過編碼蛋白調節分蘗或分枝(Gallavotti et al.,2004;Yang et al.,2012)。為獲得影響側芽起始發育調控基因的類型,對相關研究文獻進行了總結(表2)。
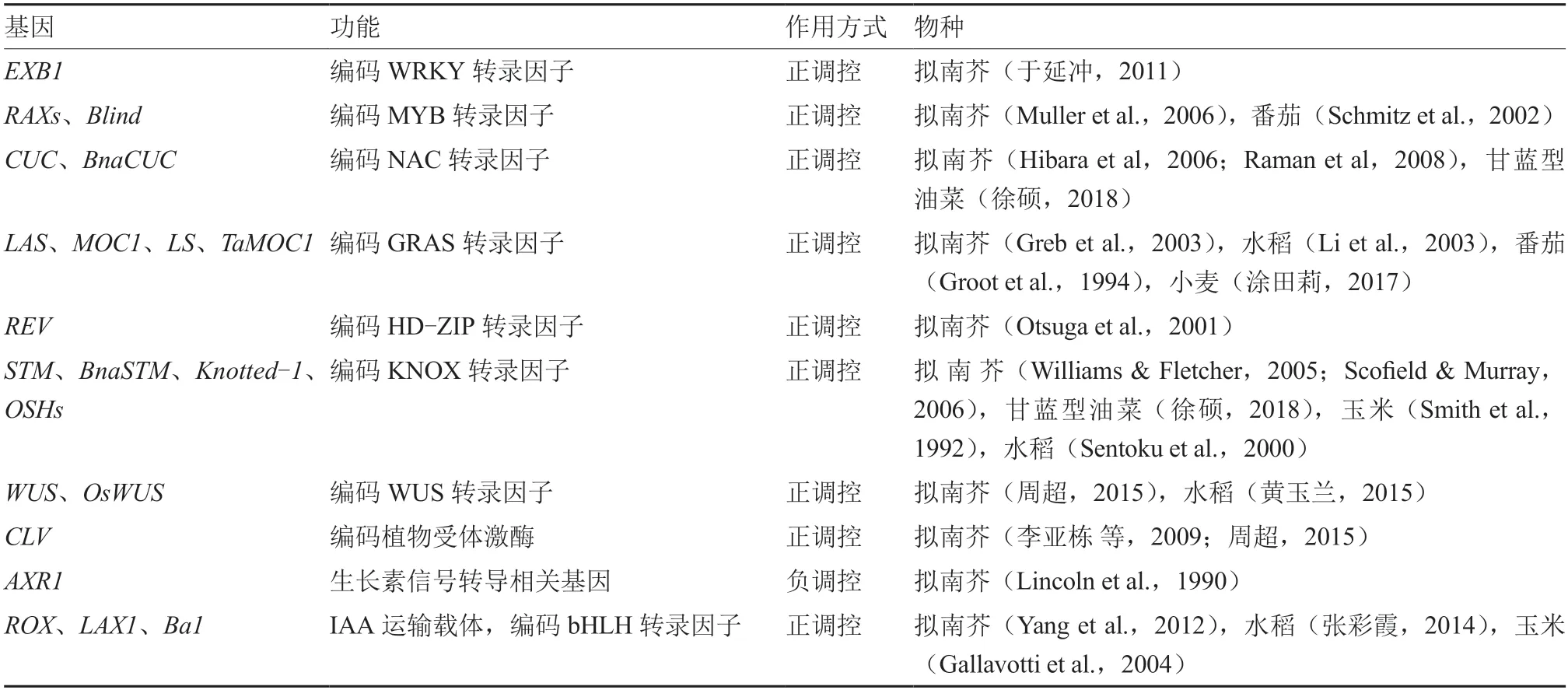
表2 側芽起始發育調控基因
3.2 側芽伸長發育調控基因
、、分別是擬南芥、玉米、水稻中的同源基因,都是TCP 轉錄因子家族的成員,抑制分枝、分蘗的生長。通過QTL 定位發現,玉米的基因造成了玉米和多分蘗Teosinte 玉米之間的結構差異(Wang et al.,1999;Hubbard et al.,2002)。主要在AM 中表達,以協調玉米芽的 生 長(Finlayson,2007)。在AM 和SAM基部均有表達,通過促進芽休眠來抑制水稻分蘗(Takeda et al.,2003)。擬南芥的基因在腋芽中表達,抑制腋芽伸長發育,是多個調控途徑的下游基因(Antonio et al.,2007)。黃瓜分枝抑制基因可以和生長素極性運輸基因協同調控黃瓜分枝(Shen et al.,2019)。基因屬于細胞色素P450 家族,通過減少IAA 含量,抑制頂端優勢促進擬南芥分枝(Bak et al.,2001)。在擬南芥中,基因是IAA 反應基因,可抑制腋芽的生長(Lincoln et al.,1990;Stirnberg et al.,1999)。()基因負調控IAA 信號轉導,可以促進擬南芥分枝(Michniewicz et al.,2007)。基因負調控CTK 含量,從而抑制擬南芥分枝(Guo &Gan,2011)。基因參與多胺的形成,突變體會導致擬南芥分枝增加(Cui et al.,2010)。基因參與水稻中SLs合成來抑制分枝(Lin et al.,2009)。基因編碼細胞色素P450,作用于、的下游,是SLs 信號轉導的關鍵基因,抑制擬南芥分枝(Booker et al.,2005)。水稻中基因與擬南芥中基因同源,是SLs 調控基因中的重要基因,在、、下游起作用,可抑制分 枝(Zou et al.,2005,2006;Hu et al.,2020)。水稻中基因與擬南芥中基因同源,與為等位基因,是編碼SLs 合成酶的關鍵基因,抑制分枝(Zou et al.,2005,2006);水稻中基因與基因同源,參與SLs 的生物合成,抑制分枝(Zou et al.,2006;Wang &Li,2011)。水稻中3 個等位基因、、在SLs 信號轉導中發揮作用,是水稻分蘗的負調控因子(Arite et al.,2007,2009;Brady et al.2007;Zhou et al.,2013)。而是SLs 信號轉導中的抑制子,正調控水稻分枝(Jiang et al.,2014)。玉米可負調控分蘗抑制基因和/-的表達,并與轉錄輔阻遏蛋白TOPLESS 互作,從而促進分蘗(Studer et al.,2011;Zhang et al.,2019)。為獲得影響側芽伸長發育調控基因的類型,對相關研究文獻進行了總結(表3)。

表3 側芽伸長發育調控基因
3.3 側芽起始和伸長發育協同調控基因
在AM 起始和側芽發育都起作用的基因是和(),它們是同源基因,基因主要在葉腋處強烈表達,控制IAA 的合成來調控擬南芥分枝(Reintanz et al.,2001)。擬南芥突變體特征是在葉腋中形成的分生組織數量增加以及側芽的停滯釋放,導致側芽大量繁殖,從而形成了可重復的分枝(Tantikanjana et al.,2001)。也有研究表明,基因通過加速CTK 的氧化降解來降低葉腋處的CTK 的濃度,使得AM 的起始和腋芽的發育都受到抑制,導致擬南芥分枝減少(Grbic &Bleecker,2000;Tantikanjana et al.,2001)。
4 植物分枝性在蔬菜育種中的應用及展望
我國蔬菜產值約2.2 萬億元,已經超過糧食作物成為種植業第一大產業。番茄和黃瓜等茄果類和瓜類蔬菜是我國設施栽培的主要蔬菜作物,且以搭架栽培為主。由于分枝較多,在茄果類和瓜類蔬菜生產中需要頻繁地進行整枝、打杈、吊蔓等耗費人工的操作,增加了人工成本;其次,分枝較多還容易造成植株養分分配不均,易發生病害,從而降低了蔬菜作物的產量和品質。隨著人工成本的逐漸增加,培育具備理想株型(側枝少或弱)、適宜輕簡化栽培的茄果類和瓜類蔬菜品種是我國蔬菜產業亟待解決的問題之一。在生產過程中,除了選擇理想株型品種之外,也可根據上述研究結果對茄果類和瓜類蔬菜的分枝性進行調控。首先,可通過改變生長環境條件來調節植株分枝。例如,可在栽培環境中適當增加遠紅光、降低溫度或增施氮肥來減少分枝,也可通過減少鉀肥和糖類物質來減少分枝。第二,可利用植物激素調節分枝。通過噴施IAA、SLs、GA、ABA、BRs 等減少分枝,同時CTK 抑制劑處理亦可減少分枝。第三,利用已鑒定的植物分枝調控基因開發分子標記,通過分子標記輔助育種培育理想株型新品種。閆立英等(2015)通過分子標記輔助選擇技術等育種手段選育出了適合棚室省力化栽培且分枝性強的優質豐產旱黃瓜品種“綠島7 號”。第四,利用已克隆的植物分枝調控基因,可以通過基因編輯技術獲得不同分枝類型新種質。Heidi(2017)對番茄基因進行編輯,獲得了有分枝且花果大小合適的番茄種質材料。在黃瓜方面,中國農業大學張小蘭教授團隊克隆了調控黃瓜分枝發育的關鍵基因,由于中國的黃瓜種植多傾向于選擇側枝數目少、長度短、萌發時間晚的品種,因此可以利用基因信息,開發分子標記,或者通過基因編輯技術鑒定、創制不同分枝類型黃瓜種質資源(Shen et al.,2019)。隨著人們對于植物分枝調控理解的不斷深入,將會有效解決茄果類和瓜類蔬菜生產中分枝較多的問題,創新種質資源,并培育出符合育種目標的優質、高產蔬菜新品種,有效解決蔬菜生產過程中用工難、用工貴等難題,降低蔬菜生產成本,提高蔬菜生產輕簡化栽培程度。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
