30個野生靈芝菌株遺傳多樣性及農藝性狀評價
2022-02-15 01:02:02張彬彬王震劉保衛趙翠敏郭金英王春霞鄭素月
江蘇農業科學 2022年2期
張彬彬 王震 劉保衛 趙翠敏 郭金英 王春霞 鄭素月







摘要:為了給靈芝的生產、馴化及遺傳育種提供種質資源,采用酯酶同工酶技術、分子標記技術(ISSR),結合出菇形態學分析了30個野生靈芝菌株的遺傳多樣性及農藝性狀,構建了聚類樹狀圖。酯酶同工酶分析結果表明,當相似系數在0.709時,可以將30個野生靈芝菌株分為7類;ISSR分析結果表明,當相似系數在0.728時,可以將30個菌株分為6類。出菇試驗結果表明,30個菌株中有25個菌株能出菇,按形態特征,在歐氏距離為3.92時可將25個靈芝菌株分為五大類群,3種方法存在一致性。通過馴化栽培及農藝性狀比較分析篩選出5個產量較高、品相較好的菌株。30個野生靈芝菌株具有較為豐富的遺傳多樣性。
關鍵詞:野生靈芝;酯酶同工酶;ISSR;農藝性狀;多樣性
中圖分類號:S567.3+10.2 ??文獻標志碼: A
文章編號:1002-1302(2022)02-0108-06
收稿日期:2021-05-06
基金項目:河北省現代農業產業技術體系食用菌創新團隊建設專項(編號:HBCT2018050202)。
作者簡介:張彬彬(1988—),女,河北保定人,碩士研究生,主要從事食用菌種質資源研究。E-mail:921940953@qq.com。
通信作者:鄭素月,博士,教授,碩士生導師,主要從事食用菌種質資源研究。E-mail:zhengsuyue@hebeu.edu.cn。
靈芝是我國傳統的中藥材,深受消費者喜愛[1]。現代研究表明,靈芝含有多種活性成分,如靈芝多糖、三萜、麥角甾醇、生物堿等[2],在抗癌、防癌、抗糖尿病、提高免疫力和抗氧化等方面有極高的藥用價值[3-7]。靈芝屬于中高溫栽培種類,野生資源廣泛分布于東亞的暖溫帶和亞熱帶地區,在我國自然分布于河北、山東、河南、安徽、江蘇、浙江、江西、湖北、湖南、四川和云南等地[8]。長期以來,可供人工栽培和加工利用的靈芝品種比較單一,不能滿足多樣化的市場需求。因此,野生靈芝種質資源的開發利用及其人工馴化研究具有重要意義。酯酶同工酶技術和分子標記技術(ISSR)檢測手段簡單、高效,廣泛應用于食用菌的菌種選育、分類鑒定和種質資源評價等研究[9-10]。靈芝屬的多數種類生長在闊葉樹的腐木上,河南省南召縣森林覆蓋率達66.49%,為靈芝提供了良好的生存環境。目前,筆者未見國內外文獻對南召縣靈芝資源的開發利用和系統深入地研究。因此,本試驗對河南省南召縣的野生靈芝進行調查收集,通過生化、分子標記技術和形態學鑒定3種方法對其親緣關系進行初步的研究,通過人工馴化篩選優良品種,以期為靈芝生產栽培和遺傳育種提供種質資源。
1 材料和方法
1.1 試驗材料
1.1.1 供試菌株
供試野生靈芝菌株30個,均于2018年8月從河南省南召縣采集,由河北工程大學食用菌實驗室分離和保藏,菌株編號見表1。
1.1.2 供試培養基
(1)馬鈴薯葡萄糖瓊脂(PDA)培養基。
(2)木屑培養基:木屑(煮汁) 200 g、豆粕(煮汁)30 g、葡萄糖20 g、瓊脂粉20 g、蛋白胨4 g、硫酸鎂1 g、磷酸二氫鉀3 g。
(3)原種培養基:小麥95.5%、石灰2%、石膏1%、磷酸二氫鉀1%、硫酸鎂0.5%。
(4)栽培種培養基:棉籽殼86%、麥麩12%、石灰1%、石膏1%。
1.2 試驗方法
試驗于2019年11月至2020年12月在河北工程大學食用菌實驗室完成。酯酶同工酶分析采用垂直板聚丙烯酰胺活性凝膠電泳方法[11]。ISSR分子引物從28個常用引物[均由生工生物工程(上海)有限公司合成]中篩選合適的引物,對30個靈芝菌株進行聚合酶鏈式反應(PCR)擴增,PCR擴增體和電泳條件參照文獻[12-13]所述方法。形態學鑒定將活化好的菌種定量(5 mm2)接種到PDA綜合培養基上,置于25 ℃恒溫培養箱培養,觀察菌絲形態,記錄菌絲生長性狀。出菇形態采用熟料栽培,常規出菇試驗,觀察子實體形態, 記錄子實體形態特征,測量菌蓋直徑、菌蓋厚度、菌柄長度、生物轉化率等。
1.3 數據處理
利用NTSYSpc分析軟件構建各菌株的親緣關系樹狀圖。按易騰飛等的方法[14]計算遺傳多樣性指數(H′),利用DPS 7.05軟件進行聚類分析。
2 結果與分析
2.1 酯酶同工酶結果分析
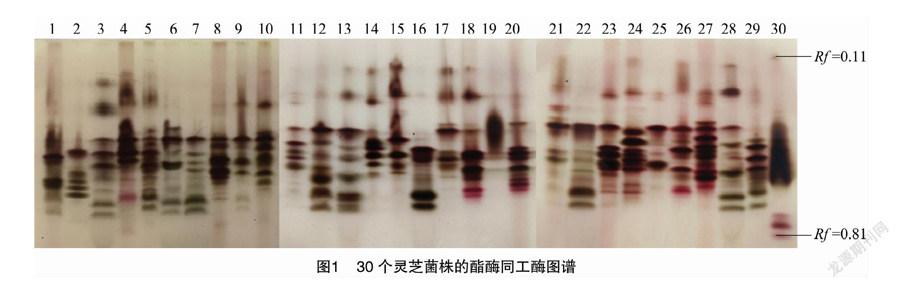
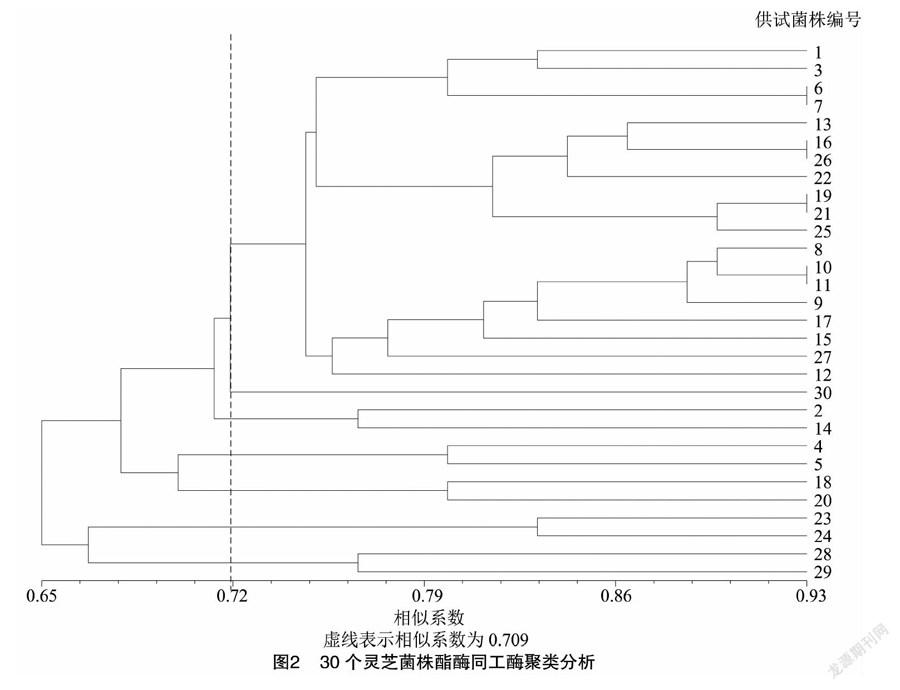
如圖1所示,菌株的酶譜遷移率(Rf)主要介于0.11~0.81之間,30個菌株共擴增出30條不同酶帶,多態性較高。對其電泳圖譜進行相似性聚類分析,由圖2可知,30個靈芝菌株兩兩間相似系數的分布范圍是0.65~0.93,當相似系數在0.709時,可以將30個野生靈芝菌株分為7類:第1類包括20個菌株,分別為YL-15、YL-8、YL-42、YL-12、YL-72、NZ-33、YL-53、YL-79、NZ-27-1、YL-37、YL-82、YL-77、YL-68、YL-83、YL-49、YL-51、YL-40、YL-31、YL-61;第2類為 YL-18;第三類為YL-90、YL-65;第4類為YL-57和YL-93;第5類為YL-88和YL-89;第6類為 NZ-28 和YL-76;第7類為YL-81和YL-14。
2.2 30個野生靈芝菌株ISSR分子鑒定結果與分析
通過對28個常用引物進行ISSR分析,篩選出7個對靈芝菌株DNA擴增重復性好、條帶清晰且有明顯多態性片段的引物,引物序列見表2。
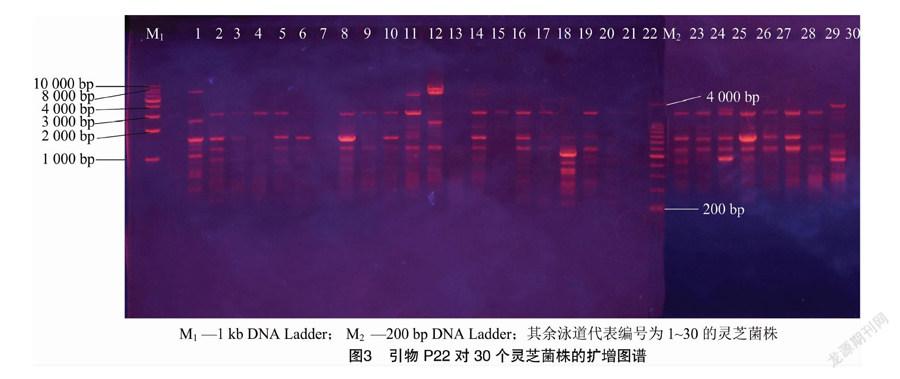
用這7個引物對30個靈芝菌株的基因組進行擴增,可獲得多態性強、穩定性好且具有一定特異性的DNA指紋圖譜,部分引物擴增結果見圖3。經統計,7個引物共擴增出85條清晰的多態性片段,DNA片段的大小介于300~8 000 bp之間,平均每個引物對野生靈芝菌株基因組擴增的條帶為12.1條。將30個野生靈芝菌株的ISSR圖譜轉換為數字矩陣,用NTSYS軟件進行聚類分析,得到供試菌株的親緣關系樹狀圖(圖4),30個靈芝菌株遺傳系數在0.62~0.89之間。相似系數在0.728時,30個菌株可以分為6類:第1類包括23個菌株,分別為YL-15、YL-57、YL-93、YL-42、YL-49、YL-12、YL-68、NZ-33、YL-79、YL-37、YL-90、YL-8、YL-40、YL-88、YL-89、YL-31、NZ-28、YL-76、YL-81、YL-77、YL-14、YL-82和YL-53;第2類為YL-83、YL-51 和YL-61;第3類為YL-18;第4類為YL-72;第5類為NZ-27-1;第6類為YL-65。說明這30個野生靈芝菌株具有豐富的遺傳多樣性。
2.3 形態學鑒定
2.3.1 菌絲形態
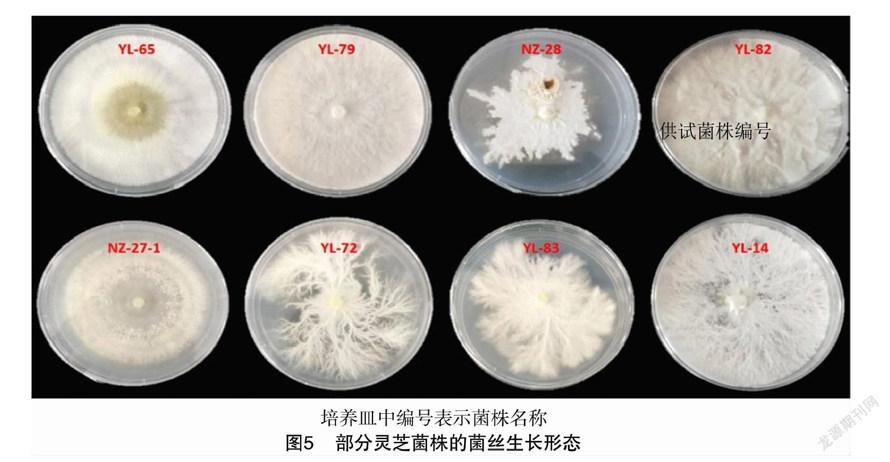
不同靈芝菌株的菌絲形態差異較大,可將30個菌株的形態類型分為四大類,部分代表菌株形態如圖5所示,分類結果和描述見表3。
2.3.2 子實體農藝性狀
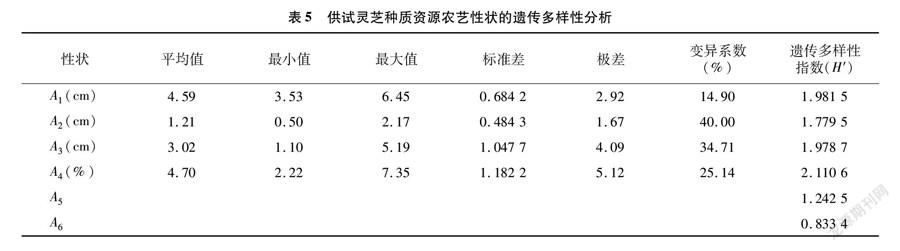
對30個野生靈芝進行馴化栽培,成功得到25個靈芝菌株的子實體,部分子實體形態見圖6。對其4個數量性狀和2個質量性狀進行記錄,結果見表4。多樣性分析結果(表5)表明,數量性狀遺傳多樣性指數以生物轉化率(A4)最高,為2.110 6,其次依次為菌蓋直徑(A1,1.981 5)、菌柄長度(A3,1.978 7)、菌蓋厚度(A2,1.779 5);質量性狀中顏色的遺傳多樣性指數(1.242 5)高于形狀的遺傳多樣性指數(0.833 4)。
如表5所示,不同菌株間變異系數存在很大差異,菌蓋厚度的變異系數最大,為40%,變幅為 0.50~2.17 cm;變異系數最小的是菌蓋直徑,為14.9%,變幅為3.53~6.45 cm。
2.3.3 農藝性狀的聚類分析
本研究采用最長距離法對不同靈芝菌株的歐氏距離進行聚類分析,繪制聚類樹狀圖。如圖7所示,在歐氏距離為3.92時可將25個靈芝菌株分為五大類群:第1類包括11個菌株,這一類多為紫褐色腎形或扇形靈芝,產量較低,生物轉化率僅為2.22%~5.03%;第2類有4個菌株,這一類多為黃褐色扇形靈芝;第3類有6個菌株,包括紅褐色扇形靈芝,產量最高高,生物轉化率最高可到7.35%;第4類包括YL-42、YL-40 和YL-81等3個菌株,為黃褐色靈芝,產量較高;第5類為1株黑褐色的鹿角靈芝。
3 結果與討論
本研究以南召縣30個野生靈芝菌株為供試材料,從生化、ISSR分子標記和形態學3個方面研究了其親緣關系。菌株的酯酶同工酶譜Rf值主要介于0.11~0.81之間,30個靈芝菌株兩兩間相似系數的分布范圍是0.65~0.93,當相似系數在0.709時,可以將30個野生靈芝菌株分為7類。ISSR篩選出的7個引物共擴增出85條清晰的多態性片段,DNA片段的大小介于300~8 000 bp之間,30個靈芝菌株遺傳相似性在0.62~0.89之間,相似系數在0.728時,30個菌株可以分為6類。通過6個主要農藝性狀對25個馴化成功的靈芝菌株進行聚類,結果顯示,當歐氏距離為3.92時,可將25個靈芝菌株分為五大類群。本研究中,酯酶聚類(圖2)、ISSR聚類(圖4)和實體形態分類(圖7)等3種方法都把YL-15、YL-8、YL-12、YL-68、YL-49、YL-53和YL-77這7個菌株聚為一類,同時也把鹿角靈芝YL-18分為一類。為增加結果的準確性,可以聯合使用3種分類方法對野生靈芝菌株親緣關系進行鑒定。通過馴化栽培及農藝性狀比較分析,篩選出5個產量較高、品相較好的菌株NZ-28、YL-31、YL-89、YL-51和YL-18。
本研究有5個菌株未馴化成功,其中NZ-33有原基產生,但是后期未分化成子實體;其余4個菌株YL-65、YL-90、NZ-27-1和YL-79未出現原基。除NZ-27-1外,其余4個菌株菌絲生長性狀良好,未出菇原因有待進一步研究。本研究為河南省南召縣野生靈芝資源的保護和合理開發利用提供了理論依據,同時也為建立野生靈芝種質資源數據庫奠定了基礎。
參考文獻:
[1]李欽艷,鐘瑩瑩,李 忠,等. 不同生長階段靈芝子實體和孢子粉活性成分分析[J]. 中國農學通報,2021,37(1):49-54.
[2]張瑞婷,周 濤,宋瀟瀟,等. 靈芝活性成分及其藥理作用的研究進展[J]. 安徽農業科學,2018,46(3):18-19,22.
[3]Bai J H,Xu J,Zhao J,et al. Ganoderma lucidum polysaccharide enzymatic hydrolysate suppresses the growth of human colon cancer cells via inducing apoptosis[J]. Cell Transplantation,2020,29(1):1-9.
[4]徐 錦,汪雯翰,楊 妍,等. 乙醇濃度對提取靈芝三萜含量的影響及提取物抗前列腺癌細胞LNCaP的活性[J]. 菌物學報,2020,39(1):155-163.
[5]Chiu C H,Wang R,Zhuang S,et al. Biotransformation of mogrosides from Siraitia grosvenorii by Ganoderma lucidum mycelium and the purification of mogroside Ⅲ E by macroporous resins[J]. Journal of Food and Drug Analysis,2019,28(1):74-83.
[6]Liu H Z,Qiu F F,Wang Y Y,et al. A recombinant protein rLZ-8,originally extracted from Ganoderma lucidum,ameliorates OVA-inducedlunginflammationbyregulatingTh17/Tregbalance[J]. Journal of Leukocyte Biology, 2020,108(2):531-545.
[7]田淑雨,鹿士峰,吳楊洋,等. 超聲破碎輔助提取靈芝多糖工藝優化及抗氧化活性研究[J]. 食品研究與開發,2019,40(8):101-107.
[8]戴玉成,曹 云,周麗偉,等. 中國靈芝學名之管見[J]. 菌物學報,2013,32(6):947-952.
[9]孔祥會,姚方杰,王 鵬. 食用菌種質資源評價方法及在品種選育上的應用實踐[J]. 中國食用菌,2019,38(12):8-10.
[10]孟 虎,孫國琴,睢 韡,等. ISSR技術在食用菌研究上的應用[J]. 北方園藝,2016(5):207-210.
[11]郭金英,宋彥龍,李 超,等. 四十個野生香菇菌株遺傳多樣性分析[J]. 北方園藝,2018(10):157-160.
[12]李輝平. ISSR在食用菌遺傳多樣性研究中的應用[D]. 北京:中國農業科學院,2007:26-27
[13]中華人民共和國農業部.食用菌菌種真實性鑒定 ISSR法:NY/T 1730—2009[S].北京:中華人民共和國農業部,2009.
[14]易騰飛,李珊珊,李嘉豪,等. 261份小麥品種基于農藝性狀的遺傳多樣性分析[J]. 河北農業大學學報,2018,41(2):7-13.
