納木措水體可培養絲狀真菌優勢種的時空特征研究
2022-02-19 12:28:30薛文凱朱攀德吉郭小芳
生態環境學報 2022年12期
薛文凱,朱攀,德吉,郭小芳
西藏大學理學院,西藏 拉薩 850000
青藏高原是世界上海拔最高、面積最大的高原,其高海拔、低氧、復雜的地貌和脆弱的生態環境形成了獨特的生態系統(Tang et al.,2017)。青藏高原的湖泊系統由1500多個大小不同的湖泊組成,其面積占中國湖泊總面積的49.5%(Wan et al.,2014)。青藏高原具有山脈眾多,太陽輻射強、降水少、蒸發量大等地理和氣候特征,從而形成了以咸水湖和鹽湖為主的湖泊系統(Zhang et al.,2019)。納木措作為青藏高原代表性的咸水湖,海拔高達4720 m,水面積超過500 km2,是地球上最高的湖泊之一(Spencer et al.,2014)。水體中的微生物在有機物和污染物的生物降解或轉化中起著關鍵作用,維持著水體的生態平衡。某些微生物的存在與特定類型的污染密切相關,是作為評估水質和氣候的重要生物指標(Glasl et al.,2017)。
絲狀真菌作為組成水體微生物的重要類群,它們適應力強,廣泛分布于各種生境中,即便在高鹽度的死海中仍能存活較長時間(Kinsey et al.,2003;Kis-Papo et al.,2003)。目前關于青藏高原水體微生物的研究主要包括細菌和真菌的多樣性及影響因子(王艷紅等,2021;鄭艷艷等;2022);自然生態系統條件下細菌群落的多樣性(Wang et al.,2018;Antunes et al.,2022);生態位方面的研究以原生動物為主(安瑞志等,2021;張鵬等,2022)。但用水體絲狀真菌的群落演變特征來反映水文環境變化的研究較少。冰川和高山湖泊生態系統是對氣候變化和人為影響極為敏感的環境。隨著旅游業的發展,人類活動對高原環境的影響日益凸顯,已有研究表明近年來青藏高原周圍地區的大氣污染和黑煙越來越多,導致高原上污染物的沉積增加(Xu et al.,2009;Cong et al.,2013;Spencer et al.,2014)。受全球變暖的影響,降水、冰川和永久凍土的消融增強,導致湖泊儲水量增加和水文條件發生改變(Zhang et al.,2020),高原湖泊正面臨著污染和氣候變化帶來的威脅。
優勢種是群落中占優勢地位的種群,研究優勢種及其生態位對理解群落中各種群間在時空上的相互關系以及對資源的競爭程度具有重要意義(Grinnell,1917;Helaou?t et al.,2009)。同時低溫的湖水環境也有較大幾率分離到耐冷的菌株。因此本研究以納木措春、夏、秋 3個季節的沿岸水體為樣本,分離純化出其中的可培養絲狀真菌,探究絲狀真菌優勢種的時空生態位和季節變化特征以及其群落中物種組成的變化規律,以期為利用微生物物種組成特征作為水質和氣候變化的生物指標提供理論基礎,為低溫環境污水的生物修復提供菌株資源。
1 材料與方法
1.1 水樣的采集
在2020年8月以及2021年5月和10月,沿湖區周邊選擇 20 個樣點(91°02′06″—90°15′31″E,30°54′32″—30°30′14″N),各樣點相距 15 km 左右(納木措禁止船只航行;為了在納木措湖周邊均勻設置樣點,在 Google地圖上按納木措湖周長與樣點數量的比值定位標記各采樣點)。用消毒過的水樣采集器,采集離湖岸15 m處的水樣,各樣點采5 L水,盛于2個無菌桶中,用于環境因子的測定和絲狀真菌的分離。在樣點信息記錄中,除 19號樣點為觀景臺,樣點存在地表徑流外,他樣點周邊植被稀疏,明顯特征。
1.2 水體環境因子的測定
測定納木措水體的 8種環境因子,水體溫度(t)、pH、總溶解固體量(TDS)、電導率(EC)和鹽濃度(Salt),以多功能參數儀(Multi-Parameter PCSTestrTM35)進行原位測定,3次重復。銨態氮(NH4+-N)、總氮(TN)以及總磷(TP)的測定,由西藏博源檢測公司完成,其中,銨態氮(NH4+-N)測定采用納氏試劑分光光度法;總氮(TN)采用堿性過硫酸鉀消解紫外分光光度法;總磷(TP)采用鉬酸銨分光光度法(邢殿樓等,2016)。
1.3 絲狀真菌的分離、純化及鑒定
用孔徑0.45 μm、直徑50 mm的無菌醋酸纖維素濾膜,抽濾50 mL和100 mL的湖水,每梯度各抽濾3個重復,抽濾后將濾膜貼于固體RBCH、酸性PDA、酸性YM平板培養基上進行20 ℃的恒溫培養,培養基配比參照亞萍等(2013)。3—5 d后統計平板上的菌落數,挑取單個菌落接種在RBCH斜面培養基上純化,繼續恒溫培養相同時間后觀察并記錄其形態特征(若有雜菌污染則再次純化),將純化好的菌株保存在體積分數為 30%的甘油管中(?20 ℃),用于高原微生物種質資源庫的建設。
分別用點植法和插片培養法培養純化菌種,觀察絲狀真菌的菌落形態和顯微特征,根據真菌的分類鑒定指標對所屬類群做初步鑒定(Ainsworth,2008)。分子鑒定由上海生工完成,其中DNA的提取、PCR的擴增采用CTAB法(孫立夫等,2009),引物為 ITS1(5′-TCCGTAGGTGAACCTGCGG-3′)和 ITS4(5′-TCCTCCGCTTATTGATATGC-3′),用BioEdit軟件校對 DNA序列并去除引物和雙峰序列,然后在NCBI核酸序列數據庫中搜索同源性菌株(選相似度98%以上的模式菌株)(Landeweert et al.,2003),最終確定絲狀真菌的分類地位。
1.4 數據處理與分析
利用R 4.0.2和Excel 2018軟件計算絲狀真菌的多樣性指數、優勢種的更替率(r)、優勢度(Y)、出現頻率(fi)以及生態位寬度(Bi)、生態位重疊(Oik),并對水環境因子的差異性、絲狀真菌的多樣性、優勢種的時空異質性以及優勢種與水環境因子的相關性進行分析。
1.4.1 多樣性指數
通過計算豐富度指數(R)、Shannon-Wiener多樣性指數(H')、Pielou均勻度指數(J')、Simpson多樣性指數(D)來判斷納木措水體絲狀真菌的多樣性。

式中:
S——絲狀真菌物種數;
Pi——i種的個體數與總個體數的比值;
N——某樣點物種的總數。
1.4.2 優勢種的劃分及更替率
確定優勢種采用McNaughton優勢度計算方法(McNaughton,1967;Habib et al.,1997),優勢種的季節更替率計算參考文獻(張皓等,2016;梁淼等,2018)。
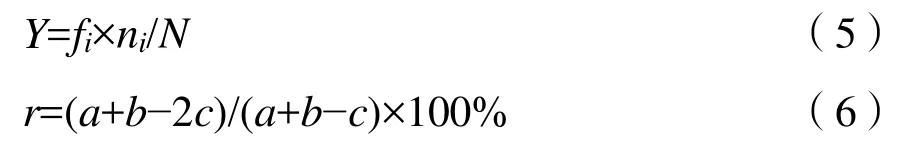
式中:
fi——i種的出現頻率;
N——樣點內的物種總數;
ni——i物種的總數;
a、b——相鄰兩季優勢種的種類數;
c——相鄰兩季共有優勢種的種類數,(Y>0.02為優勢種,Y>0.1為絕對優勢種)。
1.4.3 生態位
生態位寬度值的計算采用Colwell et al.(1971)加權修正的Levins指數。

生態位重疊值的計算采用Pianka指數(1973)。

式中:
Bi——物種i的生態位寬度值;
Pij——空間生態位中代表i種在j樣點的個體數占該樣點總個體數的比例;
r——總樣點數;
Pij——時間生態位中代表i種在j季節的個體數占i種在所有季節總個體數的比值;
r——總季節數;
Oik——物種i與物種k的生態位重疊指數,時空生態位寬度=時間生態位寬度×空間生態位寬度。
2 結果與分析
2.1 納木措不同季節水體環境因子的差異性分析
對納木措3個季節的8種水環境因子做差異性分析(圖1)。pH在春季和夏季、春季和秋季間存在極顯著差異(P<0.01),其中春季高于秋季、夏季;溫度在3個季節間的差異均為極顯著(P<0.01),且夏季>秋季>春季;NH4+-N在春季和秋季、夏季和秋季間呈顯著差異(P<0.05);TP在春季和秋季間呈顯著差異(P<0.05),在夏季和秋季間呈極顯著差異(P<0.01);TN僅在春季和夏季間存在顯著差異(P<0.05),以上3種環境因子均為春季最高,秋季最低,此外,TDS、EC和Salt在3個季節中均無顯著差異。總體而言,湖水中的部分環境因子在季節更替中出現了顯著變化。
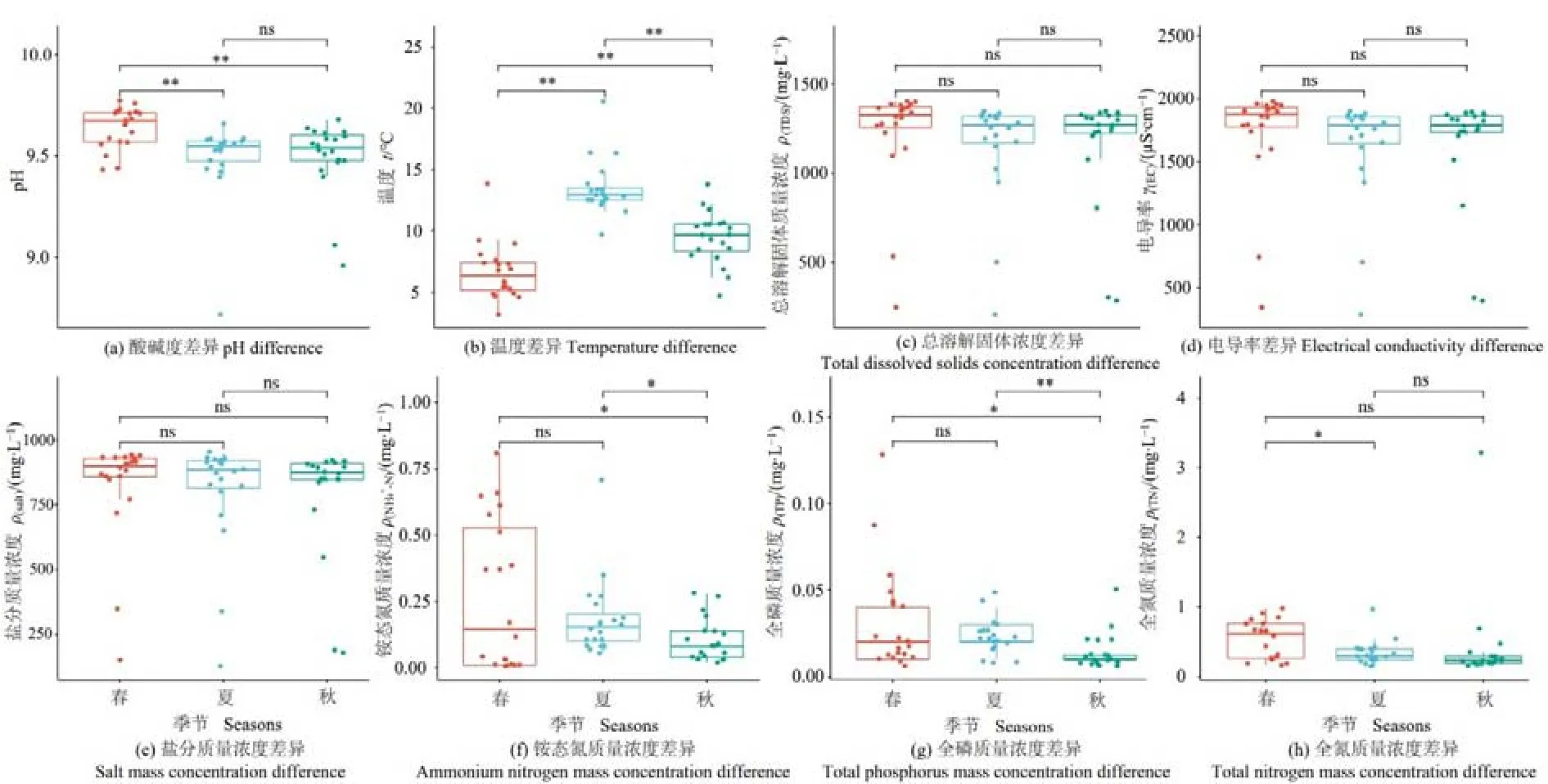
圖1 納木措不同季節水體環境因子差異Figure 1 Differences of water environmental factors in different seasons in Nam Co Lake
2.2 納木措不同季節水體絲狀真菌群落組成與多樣性的比較
在納木措3個季節中共鑒定出3359株絲狀真菌,如圖2a所示,絲狀真菌總豐度在夏季與春季、夏季與秋季間呈極顯著差異(P<0.01),且夏季最多,為1412株,其次為秋季(1026株),春季最少(921株)。絲狀真菌物種數在秋季與夏季、秋季與春季間呈極顯著差異(P<0.005),其中春季包括 20屬62種,優勢種9個,夏季包括22屬47種,優勢種13個,秋季包括13屬47種,優勢種7個。絲狀真菌的多樣性指數(圖2b),除Shannon-Wiener多樣性指數、Simpson多樣性指數在春季與夏季間無顯著差異外,其他季節間的各指數均存在顯著(P<0.05)和極顯著差異(P<0.01),其中秋季的Shannon-Wiener多樣性指數、Pielou均勻度指數均為最高,春季的Simpson多樣性指數最高。
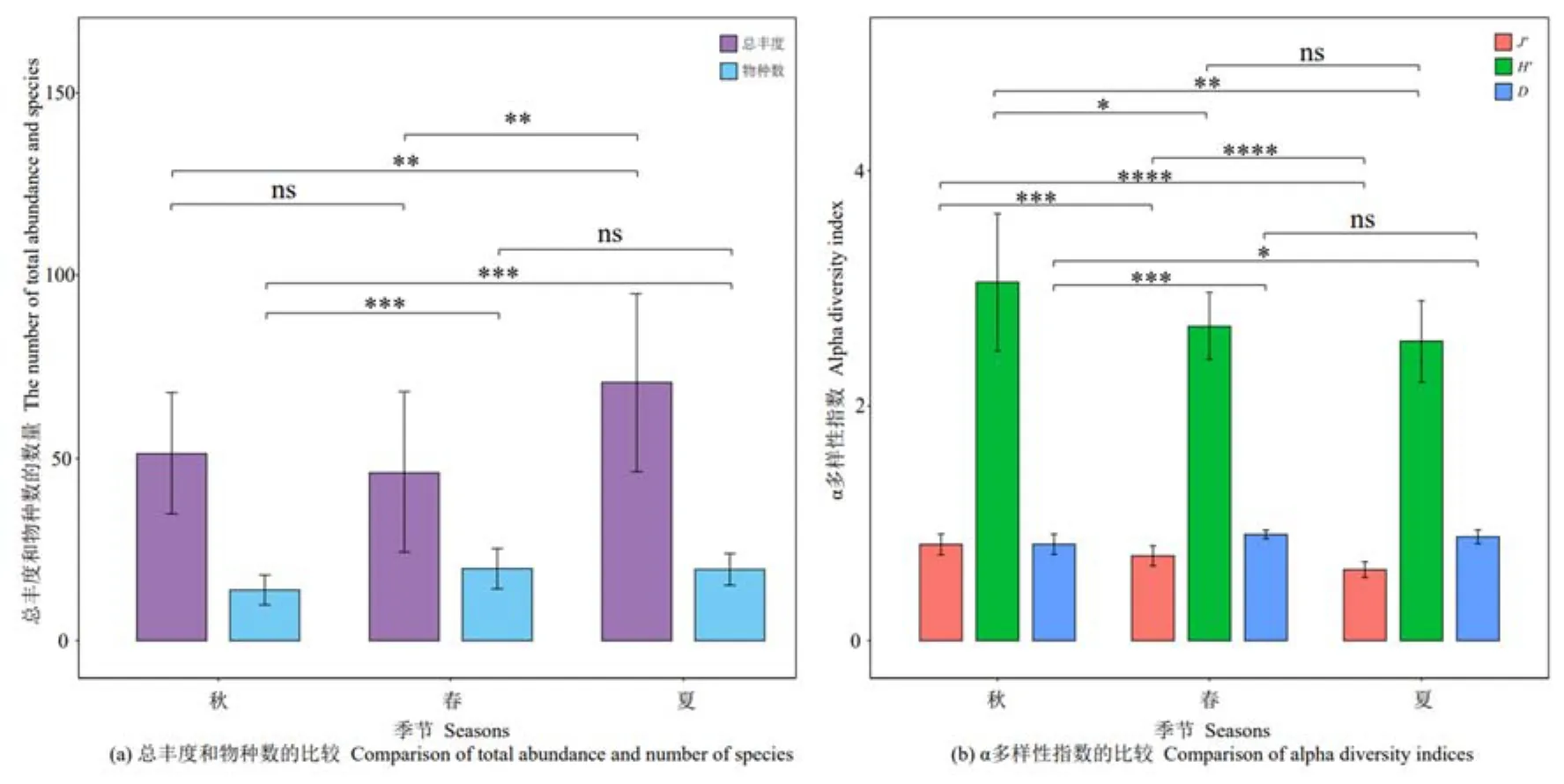
圖2 納木措不同季節水體絲狀真菌多樣性指數的比較Figure 2 Comparison of diversity indices of filamentous fungi in different seasons in Nam Co Lake
為進一步分析納木措各季節水體絲狀真菌優勢種的時空異質性(圖 3),采用 Bray-Curtis距離的非度量多維尺度(NMDS)分析,按季節分為 3組,發現秋季的2號和15號以及夏季的14號樣點中優勢種群落結構與本季節其他樣點差異較大,其余各樣點的優勢種群落結構在同一季節內的差異較小。ANOSIM 結果顯示,R=0.939,P<0.01,說明各季節間的差異顯著大于季節內樣點間的差異。
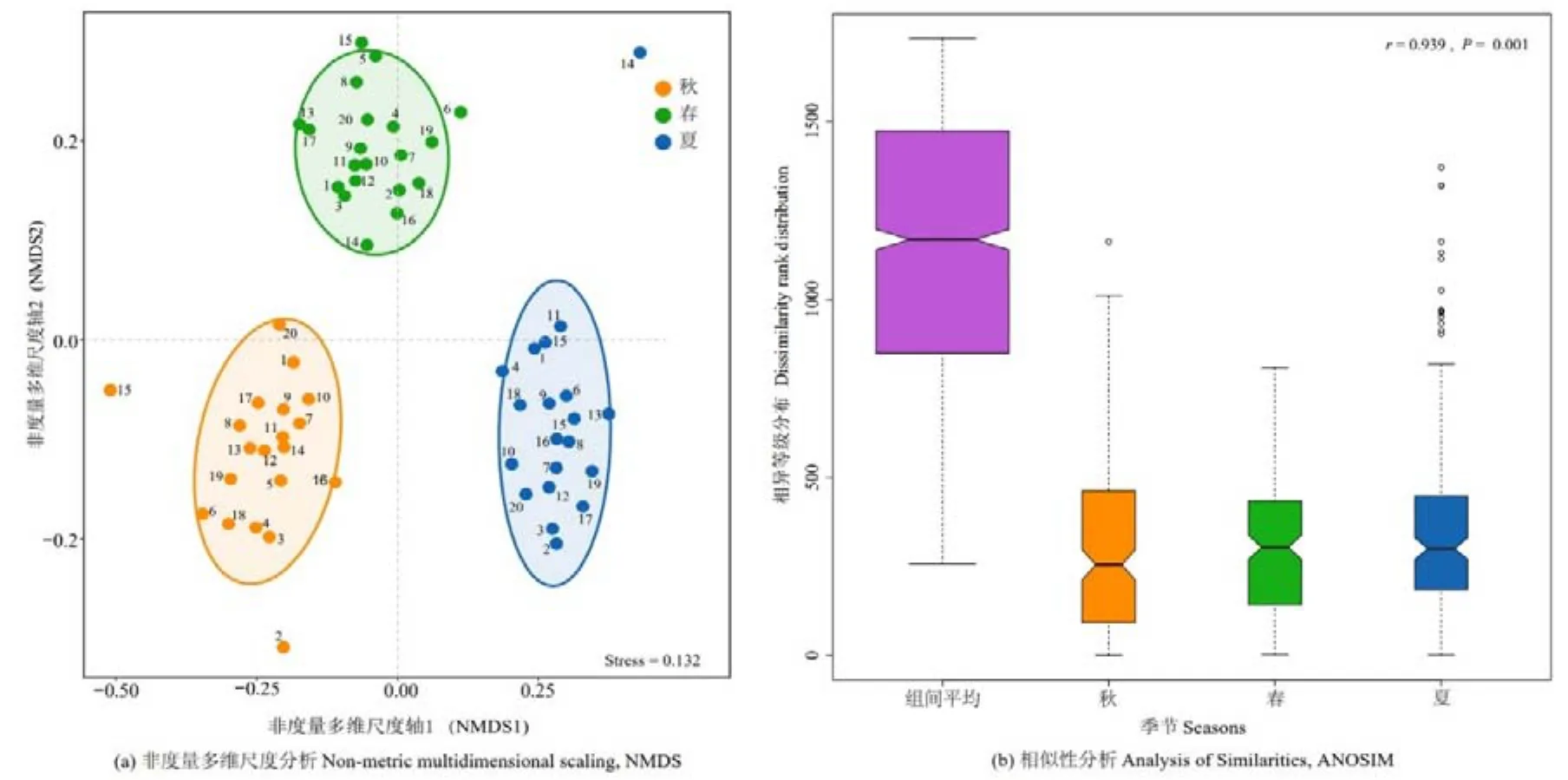
圖3 納木措不同季節水體絲狀真菌優勢種的空間變異分析Figure 3 Spatial variation analysis of the dominant species of filamentous fungi in different seasons in Nam Co Lake
2.3 納木措不同季節水體絲狀真菌優勢種和更替率
對納木措3個季節的水體絲狀真菌優勢種進行劃分,共得到優勢種23種,如表1所示,普通青霉Penicillium commune為3個季節的共有優勢種,酒色青霉Penicillium vinaceum、凍土毛霉Mucor hiemalis為春季和夏季的共有優勢種,殼青霉Penicillium crustosum、灰玫瑰青霉Penicillium griseoroseum為春季和秋季的共有優勢種。春季和秋季有獨有優勢種4個,夏季有獨有優勢種10個。由春季進入夏季時,優勢屬Penicillium的占比下降,優勢種的數量增加;由夏季進入秋季時,優勢屬Penicillium的占比上升,優勢種的數量下降。經計算,夏季與秋季優勢種的更替率最高,為95%,春季與夏季優勢種的更替率次之,為84%,春季與秋季優勢種的更替率最低,為77%,說明納木措水體絲狀真菌優勢種的季節性變化較大。
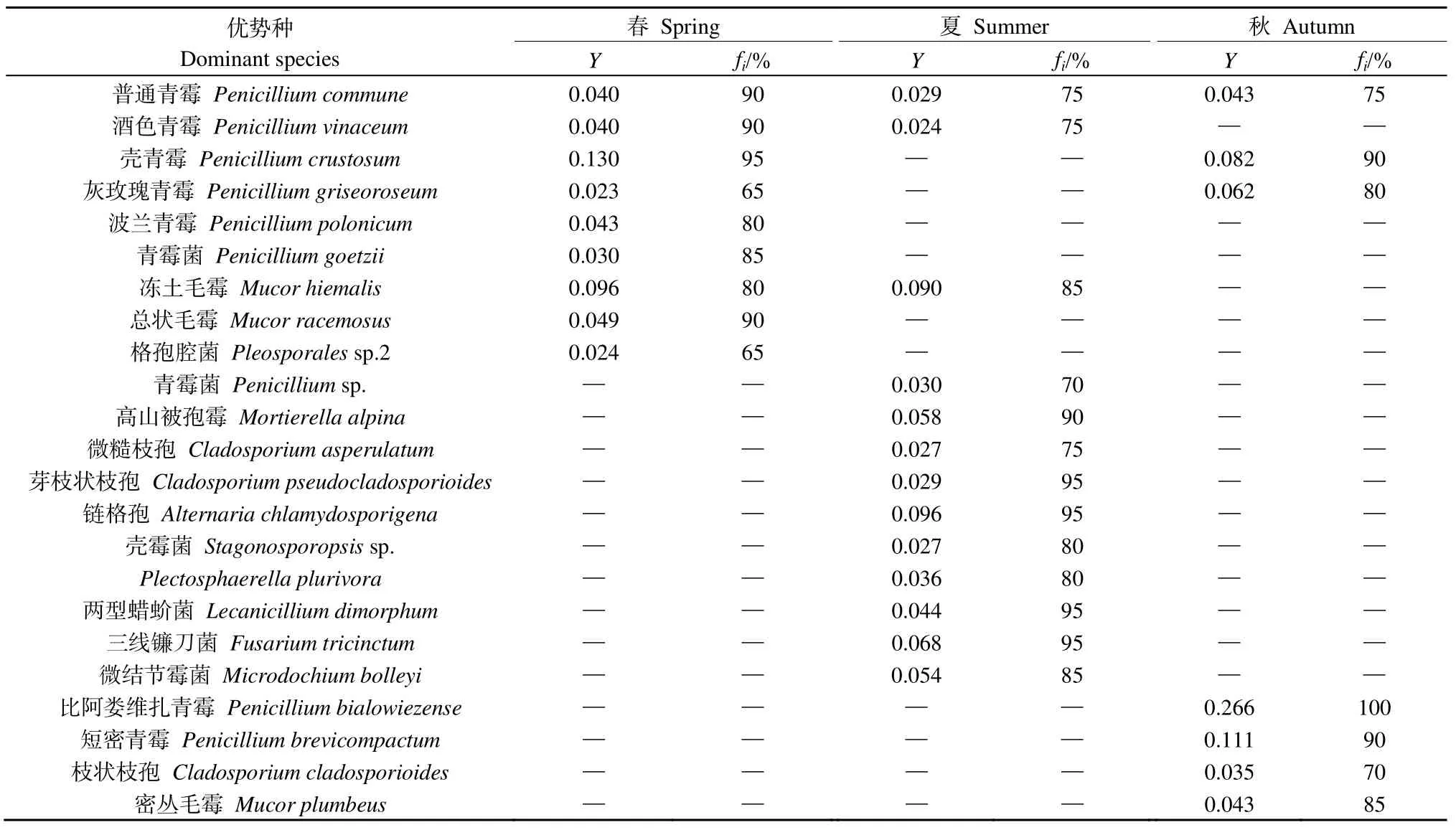
表1 納木措水體絲狀真菌優勢種在不同季節的優勢度(Y)與出現頻率(fi)Table 1 Dominance (Y) and frequency (fi) of dominant species of filamentous fungi in different seasons in Nam Co Lake
2.4 納木措不同季節水體絲狀真菌優勢種的生態位
如表2所示,納木措水體絲狀真菌優勢種的空間生態位寬度值區間在 0.017—0.942,其中殼霉菌Stagonosporopsissp.的空間生態位寬度值最高,比阿婁維扎青霉Penicillium bialowiezense的空間生態位寬度值最低。時間生態位寬度值區間在 0.333—0.979,其中P.commune的時間生態位寬度值最高,波蘭青霉Penicillium polonicum等19個菌種的時間生態位寬度值均較低。絲狀真菌優勢種的時空生態位寬度值均較小,區間在 0.006—0.314,其中P.bialowiezense的時空生態位寬度值最低,Stagonosporopsissp.的時空生態位寬度值最高,且處于窄生態位(0≤Bi<0.15)的菌種占比較大,這表明納木措水體絲狀真菌優勢種易受時空變化的影響。
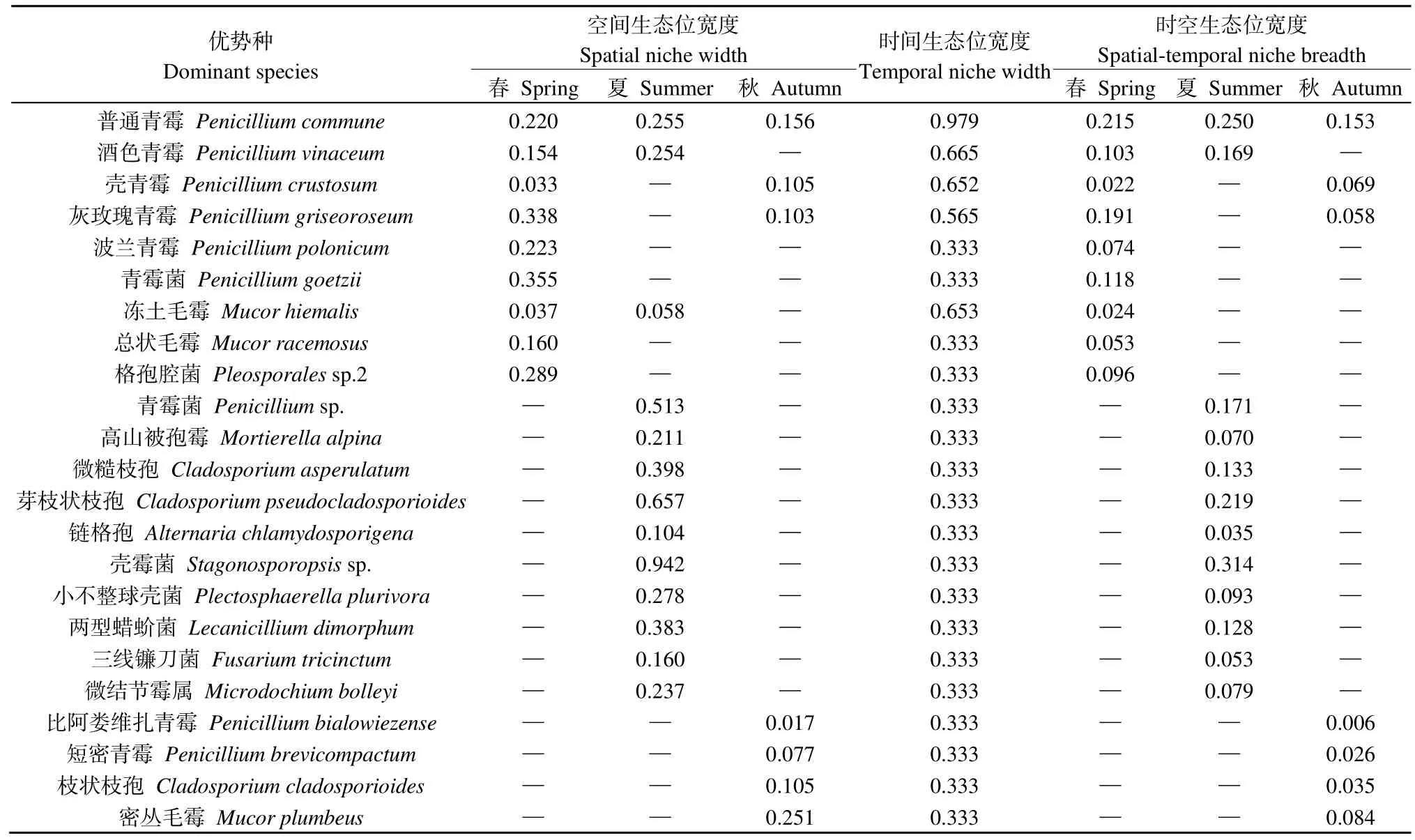
表2 納木措水體絲狀真菌優勢種在不同季節的生態位寬度(Bi)Table 2 Niche width of dominant species of filamentous fungi in different seasons in Nam Co Lake
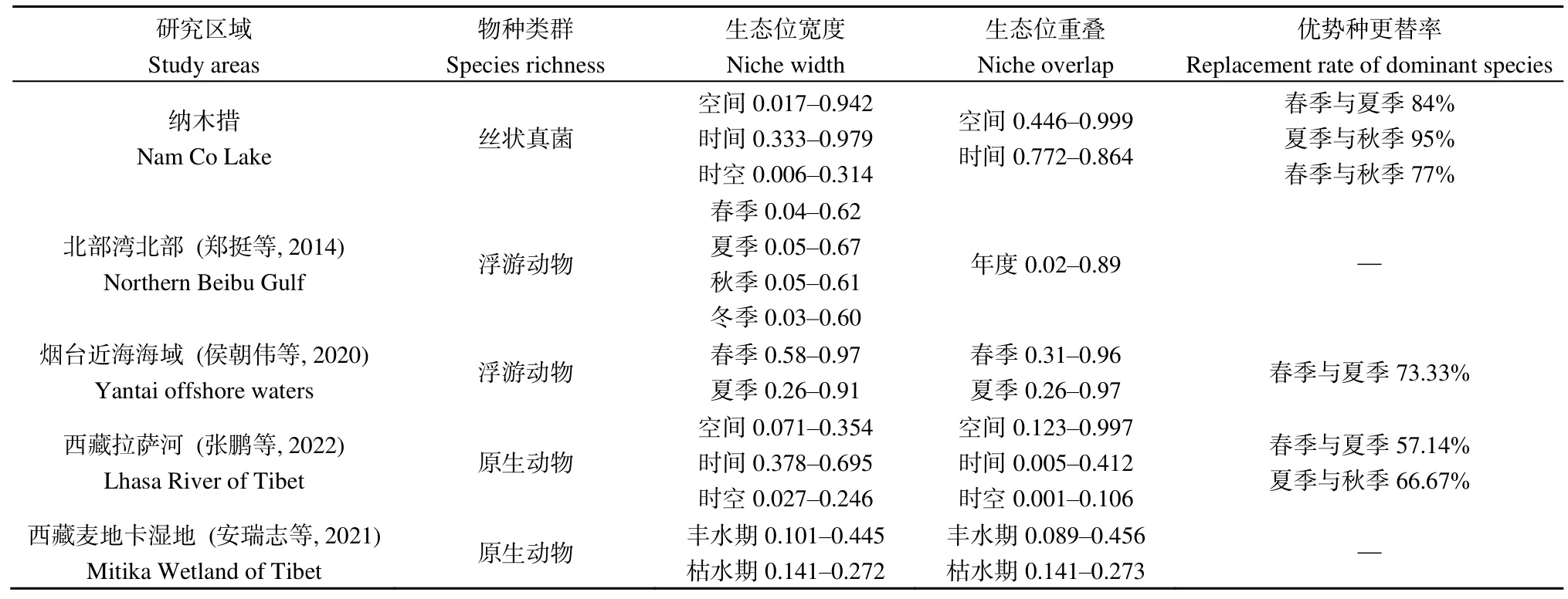
在空間維度上(表3),納木措各樣點絲狀真菌優勢種所組成的群落間的生態位重疊度較高,其中絲狀真菌群落在160對樣點間高度重疊(Oik>0.8),占總對數的 84%;在 24對樣點間中度重疊(0.6 表3 納木措各樣點水體絲狀真菌群落的生態位重疊(Oik)Table 3 Niche overlap of filamentous fungal communities at different sites in Nam Co Lake 表4 納木錯各季節水體絲狀真菌群落的生態位重疊(Oik)Table 4 Niche overlap of filamentous fungal communities in different seasons in Nam Co Lake 分析納木措不同季節的水環境因子,發現總溶解固體量、電導率和鹽濃度在3個季節中均無顯著差異,其他 5種環境因子在各季間存在顯著(P<0.05)或極顯著差異(P<0.01)。水環境因子與絲狀真菌優勢種的Spearman等級相關性分析(圖4)顯示,在春季,優勢種P.vinaceum、M.hiemalis分別與全磷、銨態氮呈極顯著負相關和極顯著正相關(P<0.01)。在夏季,優勢種Stagonosporopsissp.、高山被孢霉Mortierella alpina、M.hiemalis與電導率、鹽濃度、總溶解固體量呈顯著(P<0.05)或極顯著負相關(P<0.01);優勢種兩型蠟蚧菌Lecanicillium dimorphum與溫度、pH分別呈顯著正相關和顯著負相關(P<0.05),溫度又與小不整球殼菌Plectosphaerella plurivora呈顯著正相關(P<0.05),pH與微結節霉菌Microdochium bolleyi呈顯著負相關(P<0.05);優勢種微糙枝孢Cladosporium asperulatum與全氮、銨態氮呈顯著負相關(P<0.05),同樣全氮與鏈格孢Alternaria chlamydosporigena呈顯著負相關(P<0.05);優勢種M.hiemalis與全磷呈顯著正相關(P<0.05),與銨態氮呈極顯著正相關(P<0.01)。秋季,優勢種枝狀枝孢Cladosporium cladosporioides與鹽濃度、電導率、總溶解固體量呈顯著正相關(P<0.05),P.bialowiezense與銨態氮、pH以及P.griseoroseum與全磷均呈顯著負相關(P<0.05)。 圖4 納木措不同季節水體環境因子與絲狀真菌優勢種豐度的相關性Figure 4 Correlation between water environmental factors and the abundance of dominant species of filamentous fungi in different seasons in Nam Co Lake 納木措3個季節的水體中共鑒定出3359株絲狀真菌,各季節的多樣性均較高,且α多樣性指數間大多存在顯著差異(P<0.05)。時空異質性的結果表明,絲狀真菌優勢種在各季節間的差異顯著大于季節內各樣點的差異。有相似研究表明,生境差異能使湖泊中細菌群落的多樣性和組成產生異質性(Jiao et al.,2020;Zhang et al.,2021);納木措不同季節間絲狀真菌的多樣性和群落結構存在顯著的時間異質性,且時間異質性大于空間異質性,各季節間環境因子的差異形成生境的差異,可能是造成這一結果的重要原因。絲狀真菌優勢種的數量和物種組成隨著季節的改變發生顯著變化,各季節優勢種的更替率也較高,如其中的優勢屬Penicillium,由春季進入夏季,夏季進入秋季,其占比先下降后上升,這可能與Penicillium在低溫環境中的優勢更強有關(Cantrell et al.,2011)。結合不同季節間環境因子的變化幅度來看,環境的變化越大可能是群落越不穩定,更替率越大的重要原因。 生態位寬度是反映種群對環境適應能力和資源利用能力的重要指標(李德志等,2006;彭松耀等,2015)。在空間維度上,Stagonosporopsissp.的空間生態位寬度值最高,受空間資源的限制較小;P.bialowiezense的空間生態位寬度值最低,受空間資源的限制較大(何雄波等,2018)。在時間維度上,P.commune的時間生態位寬度值最高,受季節變化的限制較小;P.polonicum等19個菌種的時間生態位寬度值均較低,受季節變化的限制較大(黃保宏等,2005;馬玲等,2012)。絲狀真菌優勢種的時空生態位寬度值均較小,且窄生態位的菌種占比較大,總體上納木措水體絲狀真菌優勢種易受時空變化的影響。生態位重疊值反映著種群間對資源的競爭強弱(求錦津等,2018),納木措各樣點絲狀真菌優勢種所組成的群落間的生態位重疊度較高,在空間維度上,各樣點絲狀真菌優勢種組成的群落高度重疊的占84%;在時間維度上,3個季節的絲狀真菌優勢種所組成的群落間的生態位重疊度同樣較高(77.2%—86.4%)。表明各絲狀真菌群落的競爭較為激烈,且群落結構不穩定正處于發展的狀態。雖然不同季節的絲狀真菌群落間不會直接產生競爭關系,但它們之間較高的生態位重疊正是構建各個季節不同群落結構的重要動力。總體而言,不同水域或不同季節塑造了水體中絲狀真菌不同的群落結構,不同微生物種群對各種環境因子有著不同的耐受度和適應力(Williams et al.,2008),競爭優勢隨著環境變化而改變,群落發生演替,表現為各季節絲狀真菌的多樣性和物種組成產生異質性以及優勢種在各季節間較高的更替率。 微生物群落的多樣性和物種組成受到CO2、有機物、氮、磷和溫度等各種環境因素的影響(Zhang et al.,2014;Li et al.,2019;Xia et al.,2020)。時空變化對絲狀真菌群落結構的影響本質上是不同水域和不同季節的水環境因子與絲狀真菌各種群相互作用的體現,組成微生物群落的各物種彼此之間對資源存在廣泛的競爭(Esser et al.,2015;Hassani et al.,2018),環境的變化能直接影響它們的競爭力,因此,環境條件的變化造成了微生物群落的演替。相關研究表明水體真菌的組成和分布等受環境因子的影響(王艷紅等,2021),本研究中,不同季節的優勢種與水環境因子的相關性差別較大,如夏季的兩個優勢種與溫度具有顯著的正相關(P<0.05),且夏季湖水的溫度顯著高于春季和秋季,說明溫度的升高可能是造成二者逐漸成為優勢種的原因之一。 對環境響應敏感,受水體環境限制較強的某些原生動物類群作為環境指示種被認為有著可靠的生態學意義(汪志聰等,2010)。通過與其他學者相似的研究比較發現(表5),本研究中的絲狀真菌優勢類群對水環境的變化有著更敏感的響應,在季節變化中,群落的更替率和構成群落的物種變化更大。目前已有研究表明,青藏高原的變暖率遠高于全球平均水平(Liu et al.,2000),全球變暖造成冰川消融增加,冰川融水的大量輸入改變了高原湖泊的鹽度、養分和生物多樣性(Milner et al.,2009),這些水文條件的變化可能對湖泊生態系統的結構和功能產生嚴重影響(Lin et al.,2017)。且微生物對環境變化的高度敏感性,使得用生物方法評判水質和氣候的細微變化成為可能,雖然微生物檢測法需要在統一的標準下(如采樣方法、培養基的種類、稀釋倍數、培養環境等)才有可比性,但隨著技術的發展和分離方法的完善,利用微生物的多樣性、群落結構、生態位等信息作為環境變化的指標在未來具有巨大的應用潛力。 表5 納木措微生物與其他區域微生物的生態位研究對比Table 5 Comparison of niche studies of microorganisms in Nam Co Lake with those in other regions (1)納木措不同季節間絲狀真菌的多樣性和群落結構存在顯著的異質性,時間資源比空間資源的影響更顯著,各季節環境因子的差異以及不同時期環境因子與絲狀真菌之間相關性的差異是驅動其群落構建的主要動力。 (2)在時空生態位寬度以及生態位重疊上,大多優勢種處于窄生態位,且群落間重疊度較高,絲狀真菌的群落結構不穩定,在不同季節的環境條件下,一些菌種的競爭優勢隨之改變,群落發生演替,塑造了各季節絲狀真菌的多樣性和組成差異,表現為優勢種在各季節間較高的更替率。 (3)相比于其他類型的水體微生物生態位,絲狀真菌優勢類群對水環境的變化有著更敏感的響應,這種高度敏感性,可以用來評估水質和氣候的微小變化,具有較大的應用潛力。
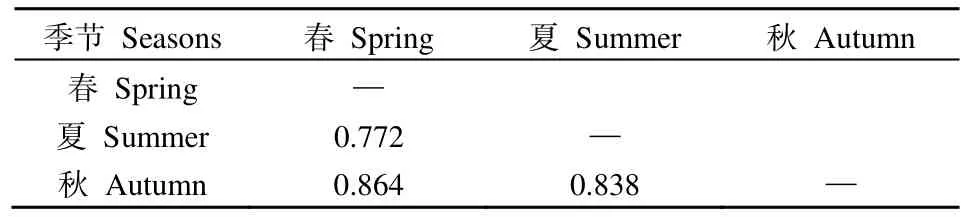
2.5 納木措不同季節水體環境因子與絲狀真菌優勢種的相關性分析
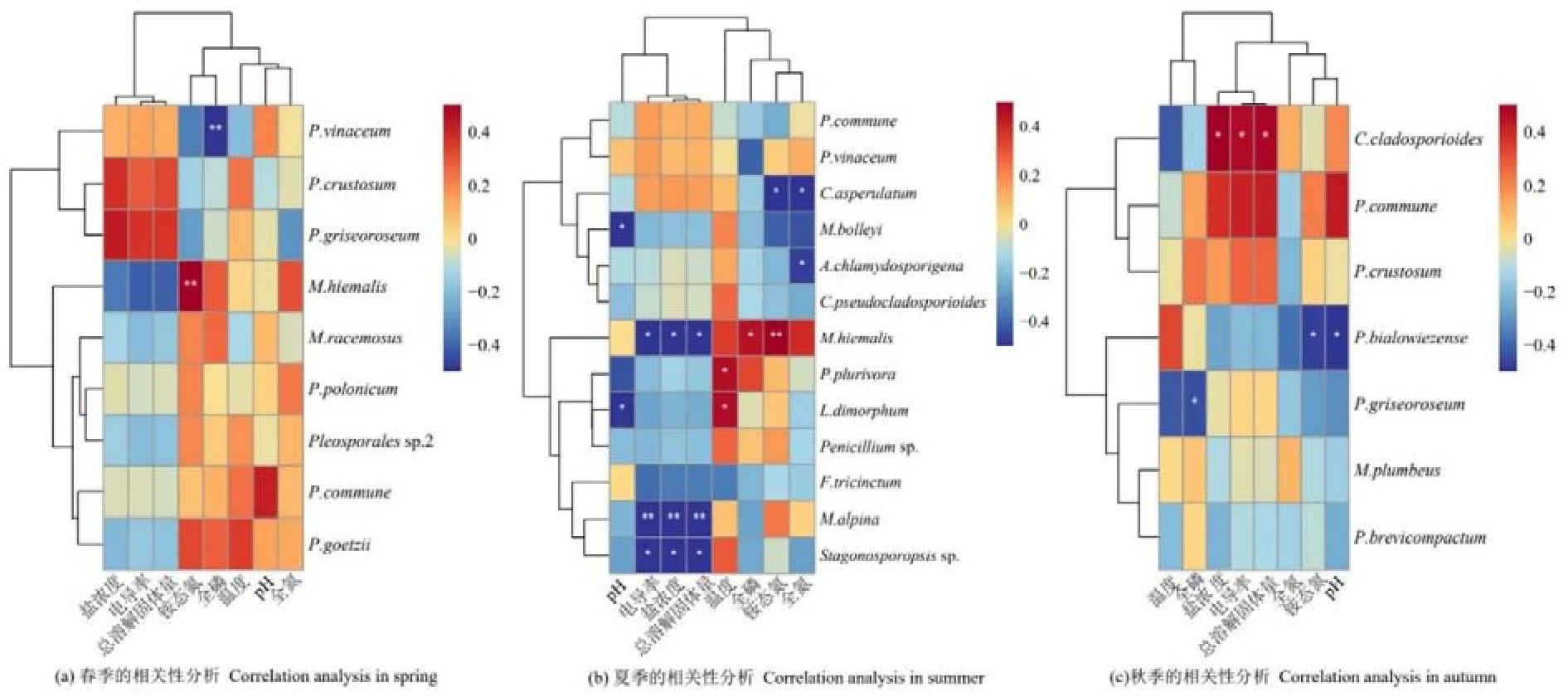
3 討論
4 結論
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
保健醫苑(2021年7期)2021-08-13 08:48:02
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
趣味(語文)(2020年3期)2020-07-27 01:42:46
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
中國生殖健康(2020年6期)2020-02-01 06:28:50
中國生殖健康(2019年11期)2019-01-07 01:28:02
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
領導文萃(2015年4期)2015-02-28 09:19:05
