大豆類胡蘿卜素裂解雙加氧酶GmCCD8固氮功能解析
2022-02-26 07:58:40邢馨竹楊占武孔佑賓李文龍杜匯李喜煥張彩英
中國農業科技導報 2022年1期
邢馨竹, 楊占武, 孔佑賓, 李文龍, 杜匯, 李喜煥, 張彩英
(河北農業大學農學院,教育部華北作物種質資源研究與利用重點實驗室,河北 保定 071001)
大豆(Glycine max)是我國種植面積最大的豆科作物,也是需氮較多的作物[1-2]。大豆可與土壤中的根瘤菌共生互作,在固氮酶的作用下進行根瘤固氮,將空氣中游離態的氮轉化為氨,一般根瘤固氮可提供植物所需氮素的50%~60%。共生固氮是一種無污染、可持續、廉價并高效地增施氮肥的方式[3],其過程是土壤中的根瘤菌通過豆科植物根毛或側根杈口侵入其根部形成侵染線,進到根的皮層,刺激宿主皮層細胞分裂進而形成根瘤[4]。根瘤菌在侵染細胞內以類菌體形式存在,由根瘤菌泡囊包圍,并隨類菌體的增多積累聚β-羥基丁酸鹽(poly-β-hydroxybutyrate,PHB)和多磷酸鹽顆粒(polyphosphate particle,PP)等物質[5],豆科根瘤中PHB的積累程度與根瘤固氮和糖代謝能力息息相關[6]。豆科植物結瘤固氮能力與根瘤鮮重、大小及固氮酶活性等顯著關聯。
在植物中,類胡蘿卜素主要存在于葉綠體和液泡中,可作為抗氧化劑防止光氧化損傷,在光合作用中發揮重要作用。類胡蘿卜素可以被裂解為一些醇類、酚類等小分子,如獨腳金內酯、維生素A、脫落酸(abscisic acid,ABA)以及一些有揮發性氣味的芳香類化合物[7-8]。分解類胡蘿卜素的類胡蘿卜素雙加氧酶可分為2類:9-順式環氧類胡蘿卜素雙加氧酶(9-cis-epoxycarotenoid dioxygenases,NCEDs)和類胡蘿卜素裂解雙加氧酶(carotenoid cleavage dioxygenases, CCDs)。NCEDs和CCDs的功能在玉米、水稻、大豆等多種作物中已被報道[9],NCEDs主要與脫落酸形成有關,而CCDs主要參與一些色素類、酚類、萜類物質和獨腳金內酯的合成。玉米中發現的類胡蘿卜素雙加氧酶VP14屬于NCED類,其被證實與ABA合成有關[10];擬南芥中通過同源性比對發現4個CCDs(CCD1、4、7、8),并證明CCDs與NCEDs具有不同的酶活性和催化底物,說明它們具有不同的生物學功能[10-11]。
據報道,類胡蘿卜素裂解雙加氧酶CCD1參與了多種植物果實成熟過程,同時與植物生殖生長及根系形成有關[12-13];CCD1在樹莓、玫瑰等植物成熟過程中參與揮發性芳香化合物形成[14];CCD4參與多種植物花和果實芳香類化合物合成和果實成熟過程,在花和其他生殖組織中高水平表達[15-18];CCD7和 CCD8 參與激素獨腳金內酯(strigolactones,SLs)合成[19],其中CCD7被證實與水稻分蘗有關[20],擬南芥AtCCD7主要在根、莖、葉、花等的維管束中特異表達[21-22];CCD8可通過調控SLs影響馬鈴薯主枝側枝和匍匐莖的形成[23];但CCDs對根瘤形成和生長的影響鮮見報道。
本研究通過分析根瘤不同發育時期的轉錄組數據發現,隨著根瘤的生長發育,GmCCD8表達量遞增,且接種根瘤菌28 d后達到最大值,暗示GmCCD8可能參與大豆根瘤的生長發育。為此,本研究通過基因克隆、實時定量、亞細胞定位、啟動子表達分析、過表達和基因干擾等技術,進一步驗證GmCCD8在大豆根瘤固氮中的生物學功能,以期為大豆根瘤固氮相關基因功能解析提供理論支撐,為大豆高效固氮分子育種奠定物質基礎。
1 材料與方法
1.1 材料與菌種
大豆品種中黃15和野生型擬南芥(Columbia)由河北農業大學大豆課題組提供;大腸桿菌TOP10感受態細胞購自北京博邁德基因技術有限公司;發根農桿菌K599購自上海唯地生物技術有限公司;慢生根瘤菌Bradyrhizobium diazoefficiensUSDA110為中國科學院遺傳與發育生物學研究所提供。
將整齊一致、顆粒飽滿的大豆種子用氯氣消毒10~12 h,置于鋪有濕潤濾紙的培養皿中28℃催芽3 d,待芽長至5 cm左右播種于裝滿蛭石的花盆中,每盆1粒,共60盆,生長條件為16 h光照、8 h黑暗,光照強度10 000 lx,溫度28℃,濕度60%,生長期間所澆灌低氮營養液(2.5 mmol·L?1K2SO4、2 mmol·L?1MgSO4·7H2O、0.5 mmol·L?1KH2PO4、0.15 mol·L?1FeCl2、0.1mol·L?1NH4NO3、0.046 mmol·L?1H3BO3、0.004 2 mmol·L?1MnCl2·4H2O、0.000 8 mmol·L?1ZnSO4、0.000 5 mmol·L?1CuSO4、0.001 mmol·L?1MoO3、0.000 1 mmol·L?1CoCl2·6H2O)。將慢生根瘤菌 USDA110 在 YMB液體培養基(北京酷來博科技有限公司)中28℃培養2~3 d,待大豆第1片真葉展開時,用注射器吸取蒸餾水稀釋的根瘤菌液(OD600=0.08)澆于大豆植株根部,每株30 mL。接種根瘤菌后10、17、28 d分別為根瘤發育初期、中期和成熟期,每個時期選取15株大豆植株上符合發育時期的大小一致的根瘤進行取樣,樣品于液氮速凍,?80℃保存備用。
1.2 試驗方法
1.2.1GmCCD8的克隆 使用天根生化科技有限公司的RNA多糖多酚植物總RNA提取試劑盒提取28 d根瘤樣品的RNA;使用反轉錄試劑盒(PrimeScript? 1stStrand cDNA Synthesis Kit,TaKaRa)反轉錄cDNA;利用NCBI中的Primer Blast設計特異引物(表1)擴增GmCCD8ORF。擴增體系參照大連TaKaRa公司PrimeSTAR@Max DNA Polymerase說明書,采用推薦三步擴增法:98℃3 min;98 ℃ 10 s,58 ℃ 15 s,72 ℃ 30 s,34個循環;72℃5 min;12℃保溫)。擴增后的PCR產物經瓊脂糖凝膠電泳鑒定、回收、純化、連接轉化等過程將目的片段連接到測序載體,篩選到的陽性克隆送蘇州金唯智有限公司進行測序分析。
1.2.2GmCCD8表達模式分析 提取10、17、28 d根瘤樣品總RNA;使用大連TaKaRa公司的反轉錄試劑盒PrimeScript?RT reagent Kit with gDNA Eraser(Perfert Real Time)反轉錄cDNA,通過BIORAD CFX96 Real-Time PCR儀,采用2×SYBR@染料法進行qRT-PCR分析(引物見表1,以actin11為內參對照)。qRT-PCR體系為:SYBR 10μL,cDNA 1μL,前引物和后引物各1μL,H2O 7μL。反應程序為:95 ℃ 30 s;95 ℃ 15 s,58 ℃ 15 s,72 ℃15 s,30個循環;10℃保溫。

表1 本研究所用引物Table 1 Primers used in this study
1.2.3 GmCCD8亞細胞定位 利用WoLF PSORT(http://wolfpsort.seq.cbrc.jp/)預測GmCCD8的亞細胞定位,利用限制性核酸內切酶XbaⅠ和BamHⅠ(TaKaRa)在37℃條件下分別雙酶切326-GFP載體和GmCCD8PCR產物30 min,然后使用T4 DNA Ligase(TaKaRa)在 16 ℃連接 16 h,轉化TOP10感受態細胞。篩選陽性克隆,凱杰生物質粒大提試劑盒提取質粒,?20℃保存。野生型擬南芥種植于光照培養室,生長條件為16 h光照、8 h黑暗,溫度23℃,濕度70%,定期澆Hoagland營養液。取20片生長4周左右的擬南芥葉片,用刀片切割后轉移到10 mL酶解液中,室溫避光酶解3 h,用血球計數板在顯微鏡下計算原生質體數量,達到約105cell·100 μL?1;吸取10 μg 326-GFPGmCCD8質粒轉化擬南芥原生質體,以35S-GFP為陽性對照,培養16 h后利用FV10i激光共聚焦顯微鏡(Olympus,日本)觀察GFP發光情況。
1.2.4GmCCD8基因表達模式分析 將GmCCD8起始密碼子上游2 000 bp序列克隆到PHY107載體,構建PGmCCD8-GUS融合表達載體,轉化TOP10感受態細胞,提取陽性克隆質粒,轉化發根農桿菌K599,誘導轉GmCCD8啟動子的毛狀根:利用注射器針頭劃傷并侵染萌發3~4 d的大豆下胚軸,種植在蛭石中,14 d后利用GUS染色技術對劃傷處生長的毛狀根進行篩選。對染色成功的陽性毛狀根接種根瘤菌,分別取接菌后10、17、28 d的根和根瘤進行GUS染色,觀察GmCCD8啟動子驅動基因的表達情況。
1.2.5GmCCD8生物學功能分析 將GmCCD8ORF利用限制性核酸內切酶XbaⅠ和BamHⅠ雙酶切30 min,T4 DNA Ligase連接16 h,克隆到帶有GUS報告基因的PCAMBIA1304過表達載體;將GmCCD8220 bp特異片段分別用KpnⅠ和SpeⅠ、SacⅠ和BamHⅠ酶切,并連接構建到帶有GUS報告基因的PTCK303 RNAi載體。利用發根農桿菌K599介導的毛狀根轉化法獲得轉GmCCD8過表達(overexpression,OX)和RNAi干擾的復合陽性植株(方法同1.2.4)。利用GUS染色技術對劃傷處生長的毛狀根染色,篩選染色成功的陽性毛狀根進行根瘤菌接種,28 d后鑒定表型,野生型根作為陰性對照。采用乙炔還原法[24]測定固氮酶活性。每處理選取30株帶有根瘤的陽性根分別放于10 mL真空管中,用注射器抽出2 mL空氣,再打進制備好的乙炔氣體2 mL,將該真空管放入28℃搖床,3~4 h后抽取1 mL氣體,用氣相色譜儀檢測固氮酶活性。色譜條件為:色譜柱為毛細管柱,載氣速度1 mL·min?1,進樣量5 μL,進樣口溫度130℃,檢測器溫度230℃,柱溫80℃。制備完乙烯氣體之后,分別摘取真空管中的所有根瘤,將根瘤稱重后,根據氣相色譜結果計算固氮酶活性[24]。
1.2.6GmCCD8RNAi根瘤透射電鏡分析 將接菌28 d的野生型和RNAi根瘤取下,用刀片切取觀察部位,將樣品浸入混合固定液中進行前固定,放置4℃冰箱2 h左右;用PBS沖洗3次,10 min后進行脫水、置換,用丙酮和812包埋劑包埋,聚合后切片干燥,用2%醋酸鈾飽和酒精溶液染色,隨后用HT7800透射電鏡(Hitachi,日本)進行掃描,觀察野生型和RNAi根瘤的內部結構[25]。
2 結果與分析
2.1 GmCCD8的序列分析
以接種根瘤菌28 d的大豆根瘤cDNA為模板,利用GmCCD8ORF特異引物擴增目的序列,得到與目的序列大小相似的擴增片段,經測序發現該基因序列長度為1 692 bp,與Phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)已公布的Glyma.06G085800序列相同。該基因組序列全長3 888 bp,包含5個內含子和6個外顯子;cDNA長度為2 046 bp,除1 692 bp的ORF外,還包含134 bp的5’非翻譯區(5’-UTR)和220 bp的3’非翻譯區(3’-UTR),編碼563個氨基酸。
2.2 不同發育時期根瘤GmCCD8表達分析
對不同發育時期根瘤的GmCCD8進行qRTPCR分析,結果表明,隨著根瘤的生長發育GmCCD8表達量持續上升,且接種后28 d的表達量與根瘤生長初期和中期相比極顯著提高,此時根瘤已成熟,并具有旺盛的固氮能力。因此,推測GmCCD8的高效表達與根瘤的生長發育及固氮能力相關(圖1)。
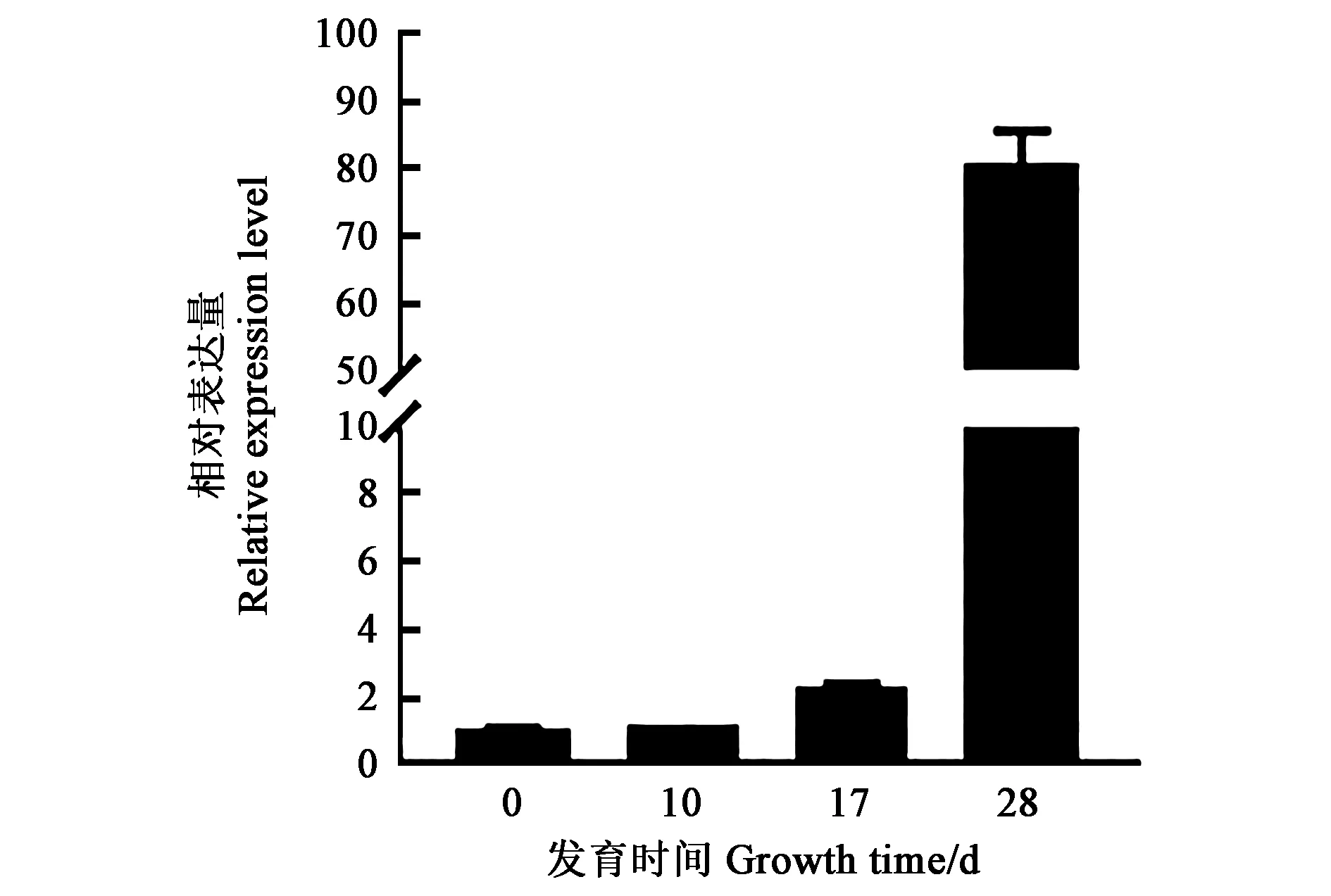
圖1 GmCCD8根瘤表達量Fig.1 Relative expression of GmCCD8 in nodules
2.3 GmCCD8亞細胞定位分析
WoLF PSORT預測GmCCD8定位于細胞質。為了對其進一步驗證,本研究利用構建的亞細胞定位載體326-GFP-GmCCD8和35S-GFP陽性對照載體轉化擬南芥原生質體,培養室培養16 h后觀察GFP熒光信號,結果顯示GmCCD8定位于細胞質(圖2),與預測結果相同,說明GmCCD8在細胞質中發揮作用。
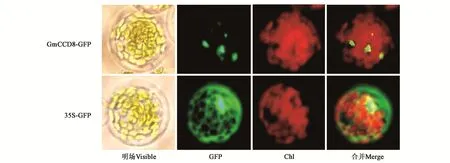
圖2 GmCCD8在擬南芥原生質體中亞細胞定位Fig.2 Subcellular localization of GmCCD8 in Arabidopsis protoplasts
2.4 GmCCD8表達模式分析
轉基因毛狀根中的GUS染色結果顯示,GmCCD8在根和根瘤中均有表達,且隨著根瘤的生長發育GUS染色逐漸加深,成熟期著色最深(圖3),與實時定量結果一致,進一步暗示GmCCD8參與根瘤的生長發育,并且主要在根瘤成熟期發揮作用。

圖3 轉GmCCD8啟動子的陽性根瘤不同發育時期GUS染色Fig.3 Histochemical GUS staining of nodules with GmCCD8 promoter in different developmental stages
2.5 GmCCD8在根瘤中的生物學功能分析
將構建好的PCAMGIA1304-GmCCD8過表達載體、PTCK303-GmCCD8RNAi載體轉化發根農桿菌并侵染毛狀根,通過GUS染色篩選陽性毛根,分析其GmCCD8表達量、根瘤鮮重和固氮酶活,結果顯示,GmCCD8在過表達陽性毛根(OX)的表達量比對照組提升2.3倍,根瘤鮮重提升36%,固氮酶活活性提高33%;與對照組相比,GmCCD8在RNAi陽性毛根(RNAi)的表達量下降了2.5倍,隨著GmCCD8表達量的下降,根瘤鮮重與固氮酶活分別降低40%和30%(圖4)。根瘤鮮重和固氮酶活是根瘤固氮能力的重要指標,由此推測GmCCD8在根瘤發育和固氮中具有重要的作用。
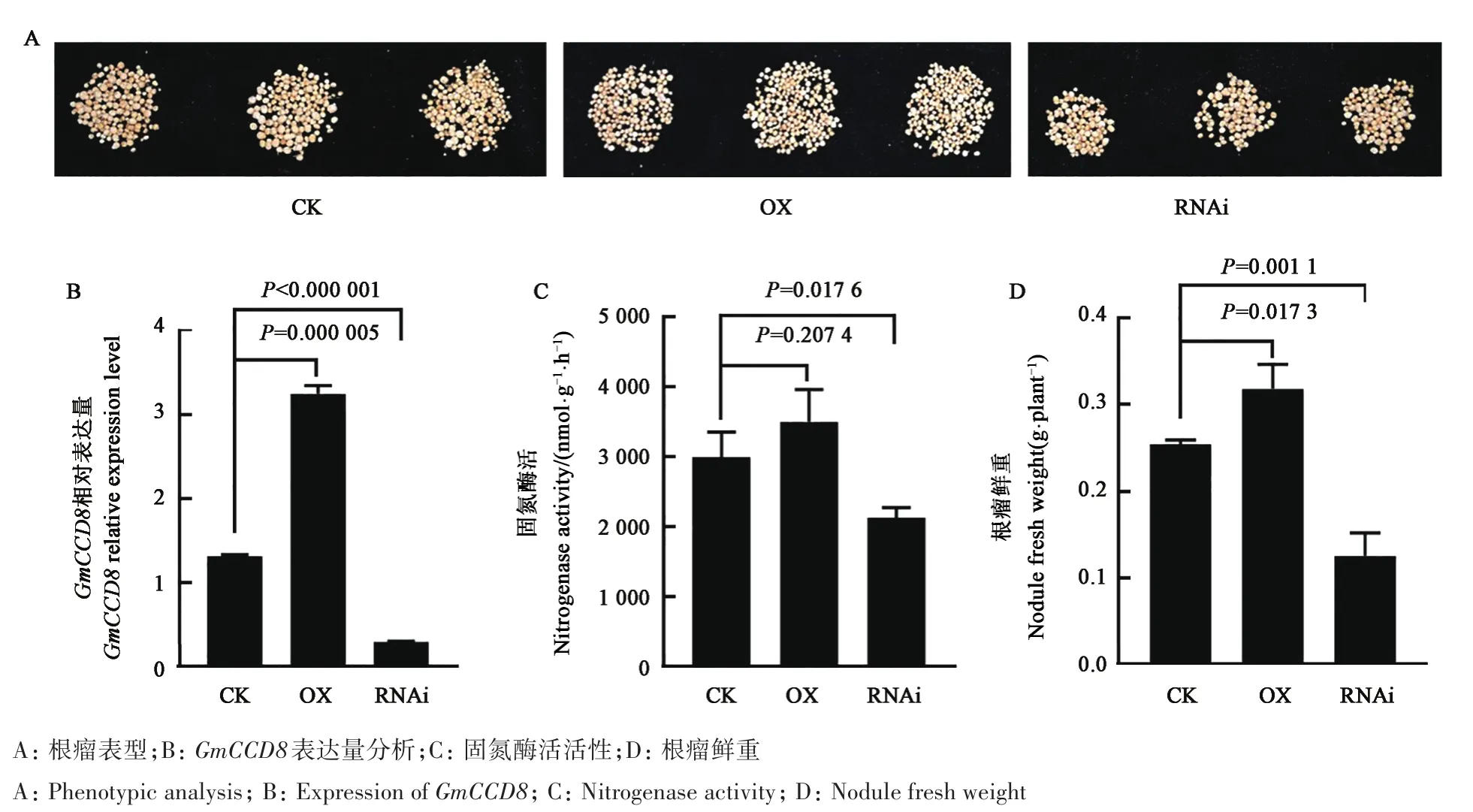
圖4 GmCCD8轉基因復合植株根瘤表型及指標分析Fig.4 Index and phenotypic of nodules in GmCCD8 transgenic composite plants
2.6 GmCCD8干擾根瘤透射電鏡分析
為了進一步驗證GmCCD8在根瘤發育中的作用,本研究對根瘤內部結構進行了觀察。透射電鏡觀察結果顯示,野生型根瘤具有正常生長的共生體,根瘤內部的根瘤菌在泡囊中分裂增殖,泡囊不斷伸長隨之根瘤也不斷加大,并且積累了大量聚β-羥基丁酸鹽(PHB);然而GmCCD8干擾后,根瘤內泡囊受損或整體變成長橢圓形且邊緣不規則化,泡囊內類菌體減少且PHB積累降低(圖5)。說明GmCCD8嚴重影響根瘤內部結構和根瘤菌與大豆根瘤的共生互作,推斷GmCCD8對根瘤發育具有重要作用。
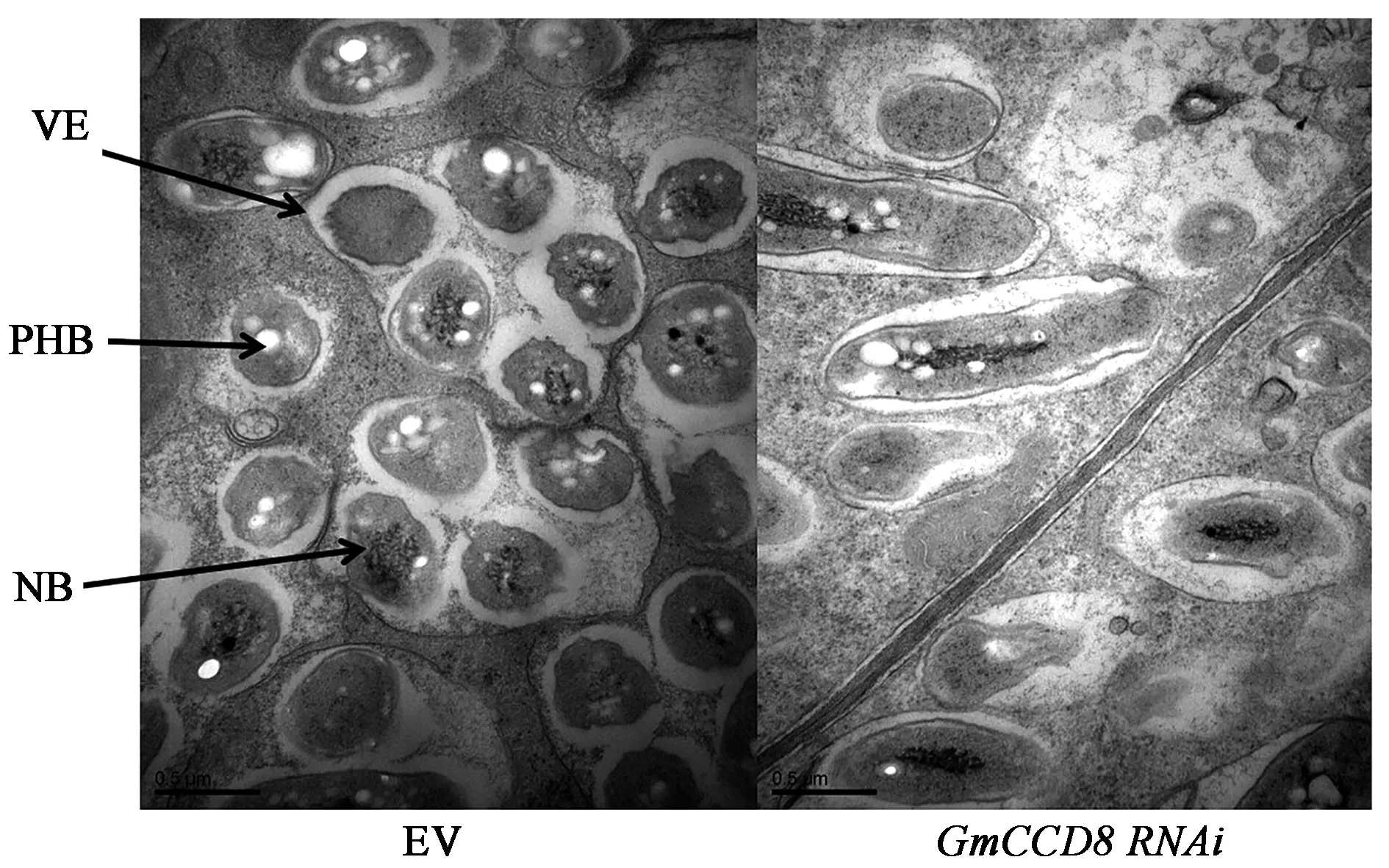
圖5 GmCCD8 RNAi 28 d根瘤顯微結構Fig.5 Nodule ultrastructure of GmCCD8 RNAi at 28 d
3 討論
諸多研究證明,類胡蘿卜素雙加氧酶在植物生長過程中發揮著重要作用,不同的CCDs表達部位不同,說明它們的功能不同。Kim等[26]研究發現,GmCCD8在根瘤中表達量較高,并且在瘤成熟期時表達量最高。本研究在根瘤不同發育時期對GmCCD8進行了qRT-PCR分析,結果顯示,隨著根瘤的生長,其表達量逐漸增加,在生長28 d的成熟根瘤中表達量與其他時期相比顯著提高,GmCCD8啟動子-GUS表達模式研究表明,隨著根瘤的生長,GUS在毛狀根和根瘤中的著色逐漸加深。上述結果說明,GmCCD8在根瘤生長中發揮著重要作用。
獨腳金內酯(SLs)及其衍生物作為新型內源性植物激素可調控植物根系發育過程,如主根伸長、側根形成、不定根及根毛生長等[21]。目前明確的SLs合成途徑為:all-trans-β-類胡蘿卜素在類胡蘿卜素異構酶催化作用下生成9-cis-β-類胡蘿卜素,隨后9-cis-β-類胡蘿卜素在CCD7裂解作用下生成 9-cis-β-apo-10’-胡蘿卜醛,繼而在CCD8的作用下生成己內酯,己內酯在己內酯氧化酶作用下生成5-脫氧獨腳金醇,再經過一系列酶促反應轉變為獨腳金內酯[21,27]。近期研究表明,SLs在根瘤的生長發育中也發揮著很重要的作用[28]。GR24是SL的人工合成類似物,在苜蓿中通過外施GR24,首次發現SLs參與根瘤菌與豆科植物共生結瘤過程,能夠促進根瘤的形成[29];豌豆CCD8突變體rms1根瘤數較野生型降低40%,同時通過對CCD8突變體rms1外施GR24可增加其結瘤數[30];苜蓿根瘤感染區中同時檢測到SLs生物合成基因MtD27、MtCCD7和MtCCD8的表達[31];在番茄中,CCD8被敲除后,列當醇和茄酚含量下降,與野生型相比主根長度變短,側根減少,列當醇是番茄根中分泌的獨腳金內酯,其含量下降說明CCD8基因的表達量影響了獨腳金內酯的含量及根系的生長[32];百脈根LjCCD7基因沉默后,根瘤數目比野生型減少20%[33]。
本研究利用毛根轉化法獲得GmCCD8過表達和RNAi轉基因根瘤,GmCCD8過表達后根瘤鮮重和固氮酶活性與對照相比分別提高36%和33%,而GmCCD8干擾后根瘤鮮重與固氮酶活性分別下降40%和30%,與前人結果一致,表明GmCCD8參與大豆根瘤的生長發育及生物固氮。
前人研究表明,根瘤中糖酵解生成的丙酮酸在進入TCA循環的同時也形成PHB,在土壤低氧環境中TCA循環會受到NADH/NAD+氧化還原狀態的限制,PHB合成會減緩這些氧化還原產物的限制來維持TCA循環,大豆固氮所需的能量主要由TCA循環提供,所以PHB的含量直接影響了根瘤的固氮能力[5,34]。本研究對GmCCD8RNAi陽性根瘤的透射電鏡觀察結果分析,GmCCD8的沉默影響了PHB的含量及類菌體與泡囊的形成,由此推測,GmCCD8會影響根瘤菌與根瘤的互作從而影響根瘤的固氮能力。
猜你喜歡
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
