iucA基因缺失對尿路致病性大腸埃希菌增殖、黏附、侵襲和定植能力的影響
2022-03-05 08:47:48康嘉樂林梅雙林艷芬
安徽醫科大學學報 2022年2期
關鍵詞:小鼠
鄧 聰,康嘉樂,林梅雙,林艷芬
尿路感染(urinary tract infections,UTI)是臨床上最常見的細菌感染之一[1],其高居不下的復發率、再感染率以及日益增高的多重耐藥菌的檢出率,強調了發展UTI新的治療策略的必要性[2]。約75%~95%的UTI由尿路致病性大腸埃希菌(UropathogenicE.coli,UPEC)所致[3]。鐵攝取系統是UPEC目前已知的毒力因子之一[4-5]。
鐵是微生物生命活動的必要元素。病原菌通過分泌鐵載體與宿主體內的鐵離子結合,形成復合物并通過外膜表面的相應受體攝取。UPEC可以分泌多種鐵載體,包括耶爾森桿菌素、氣桿菌素、腸桿菌素和沙門菌素[6]。其中氣桿菌素已被證實通過促進菌株生長、定植等,在肺炎克雷伯菌、禽致病性大腸埃希菌等的致病過程中發揮重要作用[7-8],但其與UPEC致病之間的關系至今仍不明確。該研究通過對氣桿菌素編碼基因iucA缺失株和回補株的構建,分析iucA基因在UPEC致病中的作用。
1 材料與方法
1.1 實驗材料
1.1.1菌株、質粒和細胞 大腸埃希菌CFT073菌株(臨床UTI分離的大腸埃希菌,由美國南加州大學洛杉磯兒童醫院黃勝和教授惠贈)。pKD46質粒、pKD3質粒、pCP20質粒、pRK415質粒和人膀胱癌上皮細胞株5637由本實驗室保存。
1.1.2實驗動物 24只C57BL/6雌性小鼠購自湖南斯萊克景達實驗動物有限公司。等級:SPF級。體質量:18~24 g。隨機分為3組(CFT073組、ΔiucA組、C-iucA組),每組8只。
1.1.3試劑與儀器 TaqPlus DNA聚合酶、質粒抽提試劑盒和膠回收試劑盒(大連寶生物工程有限公司);L-阿拉伯糖、氯霉素、氨芐青霉素、四環素和慶大霉素(美國Sigma公司)。 PCR擴增儀、電擊儀(美國Bio-Rad公司)、分光光度計(美國Beckman公司)。測序由上海生工生物技術公司完成。
1.2 方法
1.2.1引物設計與合成 本研究所用引物名稱、序列及用途見表1。引物合成由上海生工生物技術公司完成。

表1 引物序列表
1.2.2CFT073iucA基因缺失株的構建 外源性打靶片段的擴增與純化:以pKD3質粒為模板,iucA-F和iucA-R作為引物,擴增含氯霉素的打靶基因。
Red重組系統的誘導表達和外源線性打靶片段的電轉化:制備大腸埃希菌CFT073菌株的電轉化感受態細胞。將pKD46質粒電轉化入CFT073細胞,涂布于氨芐青霉素(100 μg/ml)平板,30℃培養過夜。挑選CFT073/pKD46單克隆,制備成電轉化感受態細胞。iucA打靶片段轉化進入CFT073/pKD46感受態細胞。轉化細胞涂布于氯霉素(chloramphenicol,Cm)平板(含34 μg/ml Cm),37℃培養過夜。隨機挑選22個克隆,接種LB液體培養基 (含34 μg/ml Cm),37℃培養過夜。用基因組同源手臂外側的iucA-outF和iucA-outR引物進行PCR檢測,當iucA基因被Cm抗性基因替換后,引物的擴增產物長度由1 921 bp縮短為1 292 bp。選擇替換成功的陽性克隆,命名為CFT073/ΔiucA-Cm。
CFT073/ΔiucA-Cm克隆中Cm抗性基因的刪除:制備CFT073/ΔiucA-Cm的電轉化感受態細胞,將pCP20質粒電轉化入上述細胞。涂布于LB平板(含100 μg/ml氨芐青霉素),30℃培養過夜。挑選單克隆劃線接種LB平板(無抗性),42℃培養過夜。隨機挑取7個克隆,分別接種LB培養基,37 ℃培養過夜。各取0.5 μl菌液用iucA-outF和iucA-outR引物再次進行PCR擴增鑒定,Cm抗性基因消除的克隆產物長度縮短為359 bp。選擇陽性克隆的PCR產物進行測序驗證,iucA被成功敲除,命名為ΔiucA。
1.2.3CFT073iucA基因回補株的構建 采用攜帶四環素抗性基因的pRK415質粒作為大腸埃希菌的回補質粒。利用iucA-comF和iucA-comR引物從大腸埃希菌CFT073原始菌株中擴增iucA基因,經HindIII和EcoRI酶切后克隆入pRK415相應位點,得到回補質粒pRK415-iucA。制備ΔiucA電轉化感受態細胞。通過電轉化將pRK415-iucA回補質粒轉化入ΔiucA菌株,在四環素平板上篩選陽性克隆。經PCR和測序驗證,確定回補質粒轉入目標菌株,選定其中一個克隆為iucA基因回補菌株,命名為C-iucA。
1.2.4CFT073、ΔiucA、C-iucA增殖曲線的測定 CFT073、ΔiucA和C-iucA菌株在LB培養液和無菌尿液中增殖曲線的測定方法見本課題組既往發表的研究[9]。
1.2.5體外黏附、侵襲實驗 凍存的細胞復蘇后至少傳代兩次,將5637細胞接種至24孔板,待長滿至細胞形成單層。按照細菌:細胞為100:1的比例將細菌接種到單層細胞。37 ℃,5% CO2孵育2 h。黏附實驗中,孵育2 h后,用PBS清洗5遍,加入0.5%Triton X-100 裂解細胞,吸出每孔樣品梯度稀釋涂板計數,作為黏附到細胞的細菌數。侵襲實驗中,每孔加入終濃度為200 mg/L的慶大霉素孵育殺死細胞外細菌,PBS清洗3遍,加入0.5%Triton X-100 裂解細胞,吸出每孔樣品梯度稀釋涂板計數,作為侵襲到細胞內的細菌數。黏附率和侵襲率計算公式如下:
黏附率=黏附到細胞的細菌數/接種總菌數×100%
侵襲率=侵襲到細胞內的細菌數/接種總菌數×100%
1.2.6體內定植能力檢測 C57B/L6小鼠尿路感染模型的建立方法及CFT073、ΔiucA和C-iucA菌株在膀胱定植能力檢測見本課題組既往發表的研究[10]。

2 結果
2.1 CFT073、ΔiucA和C-iucA菌株增殖曲線測定將CFT073、ΔiucA和C-iucA菌株分別接種于LB液體培養基和無菌尿液后,每隔2 h取樣測定A600,繪制增殖曲線顯示,在LB液體培養基中CFT073、ΔiucA和C-iucA菌株具有相似的增殖速率(F=2.613,P=0.153)(圖1)。而在無菌尿液中,三者增殖速率差異有統計學意義(F=18.927,P=0.003),其中ΔiucA的增殖速率低于CFT073菌株(P=0.001),C-iucA的增殖速率則高于ΔiucA(P=0.005),C-iucA與CFT073菌株增殖速率比較,差異無統計學意義。見圖2。
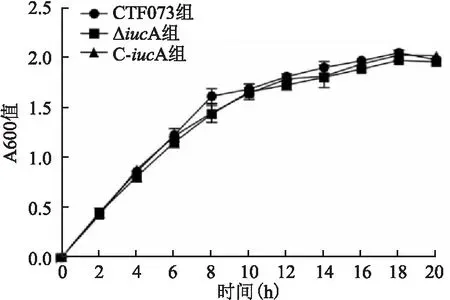
圖1 CFT073、ΔiucA和C-iucA菌株在LB液體培養基中的增殖曲線
2.2 體外黏附和侵襲能力比較比較CFT073、ΔiucA、C-iucA菌株對人膀胱癌上皮細胞株5637的黏附率和侵襲率顯示,ΔiucA組黏附率和侵襲率均低于CFT073組(P=0.007、 0.002)(圖3、4)。而C-iucA組黏附率和侵襲率與ΔiucA組相比均有所上升(P=0.046、 0.037)(圖3、4)。C-iucA組黏附率接近CFT073組,兩組之間差異無統計學意義(P=0.177)(圖3),侵襲率則仍低于CFT073組,差異有統計學意義(P=0.040)(圖4)。
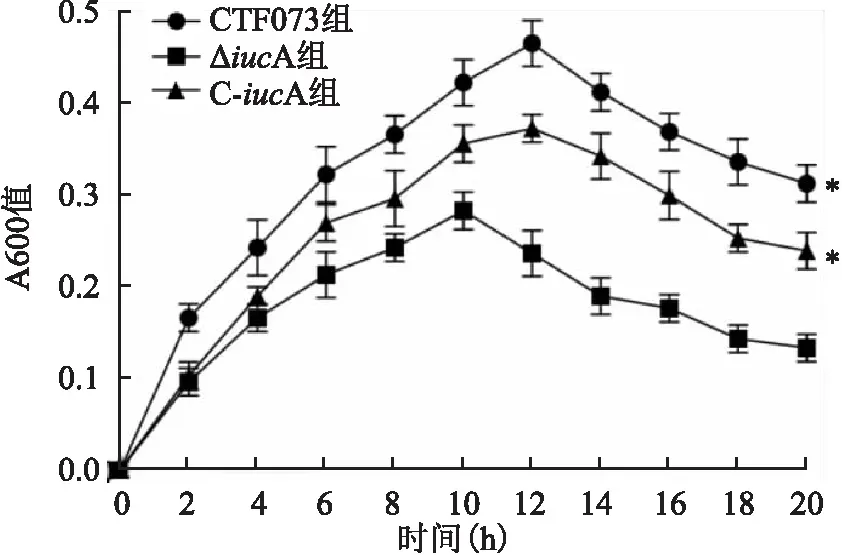
圖2 CFT073、ΔiucA和C-iucA菌株在無菌尿液中的增殖曲線與ΔiucA組比較:*P<0.05
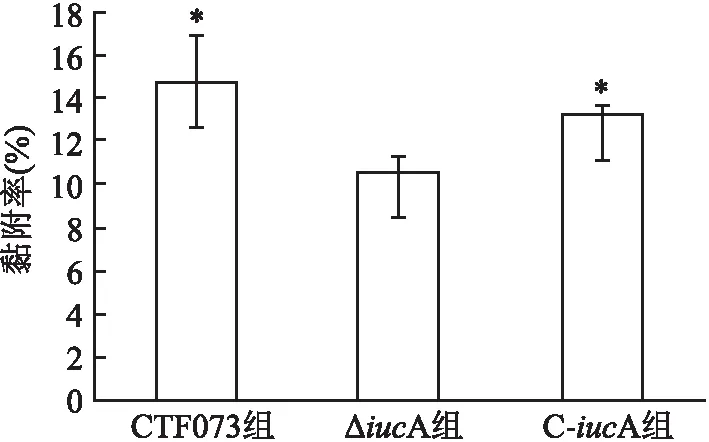
圖3 CFT073、ΔiucA和C-iucA菌株對人膀胱癌上皮細胞5637的黏附率與ΔiucA組比較:*P<0.05
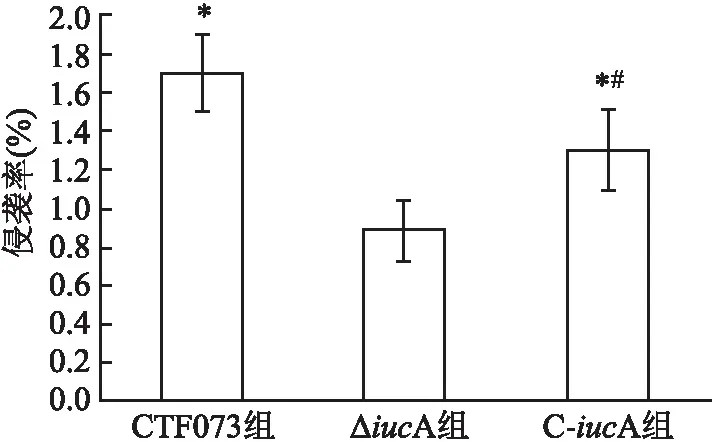
圖4 CFT073、ΔiucA和C-iucA菌株對人膀胱癌上皮細胞5637的侵襲率與ΔiucA組比較:*P<0.05;與CFT073組比較:#P<0.05
2.3 膀胱定植能力檢測成功構建C57B/L6小鼠UTI模型后,CFT073、ΔiucA和C-iucA菌株通過尿道灌注的方法感染小鼠,取膀胱組織勻漿涂平板培養計算定植細菌數。采用多個獨立樣本比較的秩和檢驗分析顯示,CFT073組侵襲到膀胱定植的細菌數平均秩次為17.75,ΔiucA組為6.00,C-iucA組為13.75,各組之間定植細菌數差異有統計學意義(Kruskal-Wallis秩和檢驗卡方值為11.545,P=0.003)。各組之間兩兩比較顯示,ΔiucA組與CFT073組,ΔiucA組與C-iucA組之間膀胱定植細菌數比較差異均有統計學意義(P=0.002、0.017), CFT073組與C-iucA組相比,定植細菌數差異無統計學意義。見圖5。
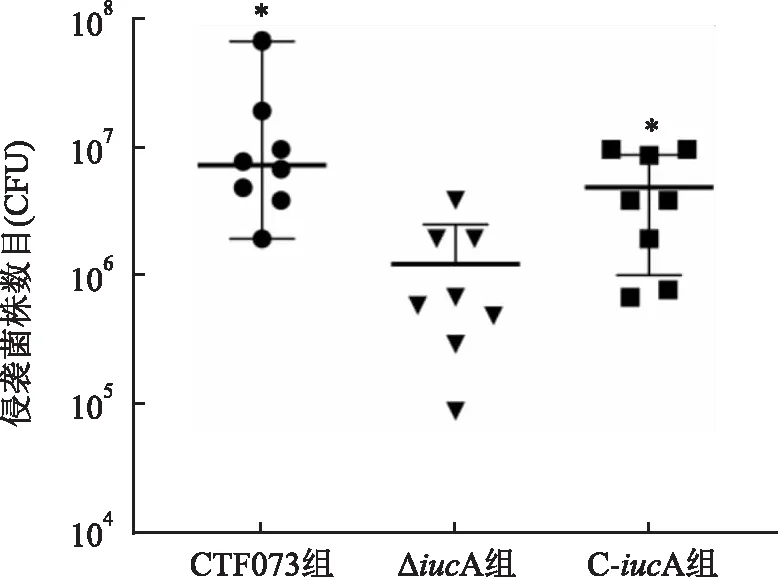
圖5 CFT073、ΔiucA和C-iucA菌株在小鼠膀胱的定植細菌數與ΔiucA組比較:*P<0.05
3 討論
鐵載體作為致病過程中重要的毒力因子和可能的治療靶標已經在多種致病菌中被證實[11]。本課題組既往研究已經證實鐵攝取相關基因fyuA在UPEC致病中的作用[9-10]。本研究繼續探究鐵載體氣桿菌素編碼基因iucA與UPEC致病之間的關系。
既往研究中,Rosso et al[12]通過分別構建高毒力肺炎克雷伯菌(hypervirulentKlebsiellapneumoniae, hvKP)的氣桿菌素、沙門菌素、腸桿菌素和耶爾森桿菌素等鐵載體的合成缺失株,發現與野生株相比,hvKP1ΔiucA菌株(氣桿菌素合成缺失株)在腹水和血清中的存活受到影響,生長速率減低,并且在小鼠動物模型中hvKP1ΔiucA菌株表現為毒力減低。而沙門菌素、腸桿菌素和耶爾森桿菌素的單獨或聯合合成缺失株均不影響菌株在體外生長/存活以及在小鼠體內毒力。提示iucA影響菌株生長繁殖,是hvKP的關鍵毒力因子。Khasheii et al[13]的研究也表明,與合成其他鐵載體的菌株相比,合成耶爾森桿菌素和氣桿菌素的UPEC表現出最高的生長速率。提示氣桿菌素可能有助于菌株的生長繁殖。本研究也顯示,盡管在LB液體培養基中野生株、缺失株和回補株表現為相似的增殖曲線,但是在無菌尿液中,iucA缺失株的生長速率低于野生株,而iucA基因的回補顯著回復菌株的生長繁殖能力,提示當UPEC處于鐵含量較低的環境(無菌尿液)時,iucA基因對促進UPEC生長繁殖具有重要作用。推測iucA基因這種對菌株生長繁殖的促進作用與其促進鐵離子的攝取作用有關,當處于營養豐富的環境(LB培養液)中時,盡管缺失了iucA基因,不能合成氣桿菌素,UPEC仍可以通過其它鐵載體介導鐵離子的攝取滿足菌株生長繁殖所需,而當UPEC處于鐵含量較低的無菌尿液中時,iucA基因的缺失進一步削弱了菌株對鐵離子的利用能力,導致生長繁殖受限。其具體機制尚需要進一步實驗證實。
黏附和侵襲是病原菌致病的第一步,強大的黏附能力可以幫助尿路致病菌抵抗尿液的沖刷,黏附在上皮細胞后再侵襲進入胞內發揮致病作用。在本研究中,通過野生株、缺失株和回補株分別與人膀胱癌上皮細胞株5637相互作用顯示,iucA缺失株的黏附率和侵襲率均低于野生株, 而在回補了iucA基因后,黏附率和侵襲率與缺失株相比均有所上升。提示iucA基因在促進菌株黏附和侵襲方面同樣具有重要作用。為進一步明確iucA基因這種介導菌株黏附和侵襲的作用是否有助于菌株在靶器官的定植,課題組建立了小鼠尿路感染的動物模型。3種菌株分別感染C57B/L6小鼠后,缺失株在小鼠膀胱的定植數目低于野生株,而回補株可以回復其在膀胱組織的定植能力,提示iucA基因有助于增強UPEC在靶器官的定植能力。這與Ling et al[8]發現缺失iucA基因的禽致病性大腸埃希菌在肝臟、腎臟、脾臟、心臟和肺臟等的定植能力大幅減低的研究結論相似。iucA基因增強UPEC在靶器官的定植能力的機制,可能部分與其促進菌株的生長繁殖以及增加菌株的黏附和侵襲能力有關。
本研究通過iucA基因的缺失和回補,明確了鐵載體氣桿菌素在促進UPEC生長繁殖、體外黏附和侵襲以及體內靶器官定植中的作用。這有助于篩選與UPEC致病密切相關的鐵載體,可以為UTI抗菌藥物作用靶點、疫苗候選抗原選擇奠定基礎,并且對其他腸桿菌科細菌的致病機制研究具有借鑒意義。iucA基因介導UPEC生長、黏附和侵襲的具體作用機制未來還需要通過進一步的實驗研究明確。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
