基于CRISPR/Cas9技術對水稻細胞質分裂關鍵基因OsMAP65-3s的編輯
2022-03-11 06:58:58林小麗姜志樹胡嘉敏朱昌蘭周大虎賀浩華
核農學報 2022年4期
關鍵詞:水稻
林小麗 姜志樹 賀 榕 胡嘉敏 朱昌蘭 周大虎 賀浩華 徐 杰
(江西農業大學農學院/作物生理生態與遺傳育種教育部重點實驗室/江西省超級稻工程技術研究中心/雙季稻現代化生產協同中心,江西 南昌 330045)
細胞分裂是植物發育中重要的生理過程,細胞在該階段由一個分裂成兩個子細胞。細胞分裂既調控植物器官的大小,也為植物適應多種環境提供可塑性。細胞質分裂發生在減數分裂或有絲分裂的后期至末期,此過程是細胞一分為二形成子細胞的關鍵,也是高等植物和動物正常生長發育以及繁衍后代的重要環節[1]。細胞分裂后期,紡錘絲將染色體拉向細胞兩極,然后子核開始形成,兩個子核之間增加了許多短的紡錘絲,形成一個密集著紡錘絲的桶狀區域,稱為成膜體。成膜體是體細胞中有絲分裂細胞的一個反平行微管陣列,反映動態微管網絡最顯著的特征之一[2]。當細胞進入到分裂末期,桶狀成膜體外圍的微管聚合,而中間的微管發生解聚,成膜體圍繞著生長的細胞板向外呈放射狀擴張,變成環狀結構[3-4]。
成膜體微管呈現了非常不穩定的動態,這也是成膜體膨脹、細胞板合成所必需的。成膜體微管的排列組裝還需要許多具有特定能力(如微管成核、聚合、成束等)的調節因子參與,而微管成束過程由微管交聯蛋白MAP65家族二聚化介導[5-6]。植物MAP65蛋白最早純化自煙草BY-2細胞,大小為65 kD,該蛋白具有微管結合活性,在整個細胞周期中與BY-2細胞中的微管共定位[7]。Chan等[8]從胡蘿卜細胞骨架中純化出兩種MAP65蛋白,均對細胞微管有修飾作用。在擬南芥基因組中存在9個MAP65家族成員,通過蛋白序列比對,表明上述蛋白的氨基酸水平與煙草MAP65蛋白有28%~79%的同源性[9]。擬南芥(Arabidopsisthaliana)所有MAP65家族成員都可作為微管成束因子,但家族成員的表達模式、細胞定位和靶標微管都存在差異,表明其在細胞內的功能可能不一樣[10-11]。MAP65-1和MAP65-2組成型表達于所有被檢測的細胞[12],而AtMAP65-4定位于分裂后期的紡錘絲中間以及成膜體中,Damme等[13]研究觀察根尖分生組織的分裂過程,發現該蛋白于早前期帶(preprophase band,PPB)時期開始表達,并一直持續到細胞質分裂完成。AtMAP65-6定位于線粒體,可能參與細胞器與微管之間的互作[14]。MAP65-3是細胞質分裂特異的MAP65家族成員,與其他家族成員不同,AtMAP65-3只與成膜體微管結合,并集中于微管的正極,即位于成膜體中線區域[11,15],是調節植物胞質分裂的重要因子。atmap65-3突變體植株矮小,生長發育受到嚴重影響,進一步通過細胞學觀察,發現突變體成膜體中間區域變得更寬,微管束排列松散,且高達40%細胞的細胞質分裂出現問題,突變體根系發育受到影響[15-16]。
水稻(OryzasativaL.)是我國最重要的糧食作物之一,也是單子葉模式植物。系統研究水稻細胞分裂不僅可以補充和豐富植物有絲分裂的分子調控機制,也可以為調控水稻發育提供新的思路。但是,由于細胞分裂對植物發育的重要性(突變易致死)以及家族成員功能冗余(突變無表型),導致水稻細胞分裂突變體難以獲得,相關的研究無法開展。目前,水稻細胞分裂的相關研究較少,尤其是胞質分裂過程的相關研究幾乎空白。由于MAP65-3是調控細胞板的合成及細胞質分裂的關鍵因子,因此水稻中MAP65-3同源蛋白是開展水稻細胞分裂工作的突破口。水稻中包含11個MAP65成員[17],其中LOC_Os01g49200和LOC_Os05g47970與AtMAP65-3同源性最高,其氨基酸同源性分別高達55%和53%,分別命名為OsMAP65-3.1和OsMAP65-3.2。江西農業大學作物生理生態與遺傳育種實驗室前期通過首創的煙草葉片瞬時分裂系統,發現OsMAP65-3.1和OsMAP65-3.2定位于成膜體中線區域,表明水稻中的2個基因可能與擬南芥AtMAP65-3基因功能相似,在胞質分裂中通過調控成膜體微管動態參與水稻生長發育。因此,創制和獲得水稻MAP65-3基因突變體是開展功能研究的首要任務,對闡明水稻細胞分裂和胞質分裂調控機理十分必要。
本研究采用CRISPR/Cas9技術對水稻OsMAP65-3.1 和OsMAP65-3.2進行同時編輯,高效、快速、特異地創制單基因突變體和雙基因突變體。分別針對2個基因設計特異性靶位點,構建至同一個載體,用農桿菌介導的遺傳轉化獲得轉基因植株,測序分析靶位點序列,篩選編輯植株和突變體,旨在為探討OsMAP65-3基因的功能和水稻細胞分裂調控機制奠定基礎。
1 材料與方法
1.1 試驗材料及田間管理
大腸桿菌DH5α,感受態采用CaCl2法,參照《基因工程》[18]的方法制備。農桿菌EHA105,感受態采用Milani等[19]的方法制備。
植物材料為粳稻品種日本晴(Nipponbare),轉基因材料均種于人工氣候室,培養條件如下:28℃光照培養12 h,25℃黑暗培養12 h,光照強度為10 000 lx。
1.2 sgRNA設計
參考CRISPRdirect數據庫(http://crispr.dbcls.jp/)設計OsMAP65-3s的特異靶位點。對OsMAP65-3.1 與OsMAP65-3.2的編碼區序列進行掃描分析,以原間隔子鄰近序列(protospacer adjacent motifs,PAM)前20堿基作為候選位點,日本晴全基因組為參考序列,選擇特異性高的序列為靶位點,靶位點選擇靠近目標基因5′端,旨在提高無功能蛋白的發生概率從而實現目標基因敲除。
1.3 載體構建
以pENTR-sg4作為入門載體(圖1-A),pUbi-Cas9作為目標載體(圖1-B),使用Gateway重組系統構建OsMAP65-3.1與OsMAP65-3.2雙基因敲除載體。首先,針對每一個設計的靶位點,合成兩條反向互補核苷酸單鏈,分別在5′端添加TGTT和AAAC(pENTR-sg4載體BtgZ I消化形成的突出末端)或GTGT和AAAC(pENTR-sg4載體BsaI消化形成的突出末端),退火復性形成帶粘性末端的靶位點雙鏈片段;其次,用BtgZ I線性化pENTR-sg4入門載體,使用DNA連接酶將第一個基因靶位點片段整合,熱激法轉化大腸桿菌,PCR檢測后測序;然后,用BsaI線性化中間載體(已插入第一個基因靶位點的入門載體),通過連接插入第二個基因靶位點片段,轉化大腸桿菌,PCR檢測后測序;最后,利用LR重組酶,將插入2個基因靶位點片段入門載體與pUbi-Cas9目標載體進行重組,轉化大腸桿菌,PCR檢測后測序。
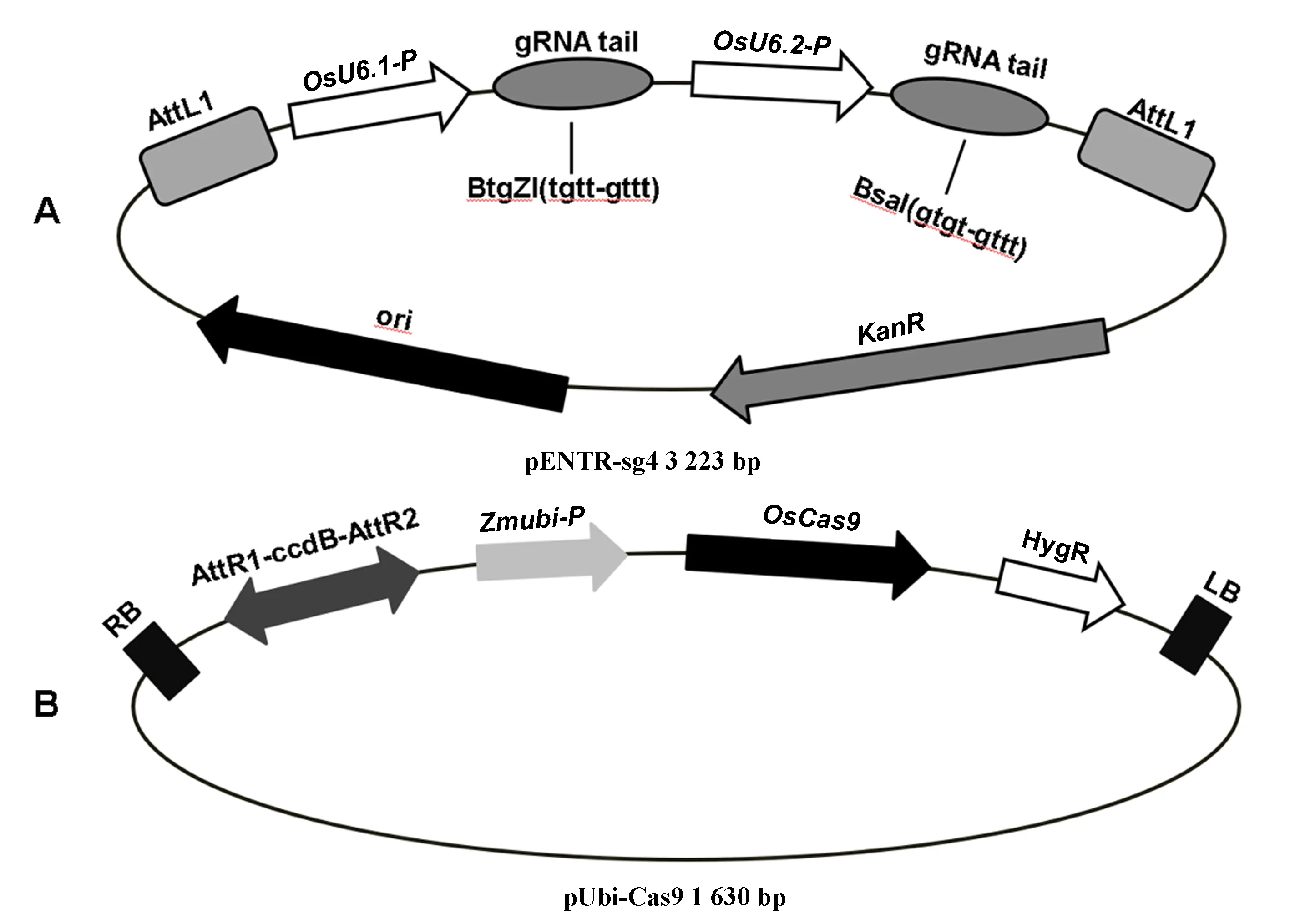
注:A:入門載體PENTR4-sg;B:目標載體pUbi-Cas9。OsU6:水稻U6啟動子;Zmubi-p:玉米Ubi啟動子;LB:Ti質粒左邊界;RB:Ti質粒右邊界;OsCas9:水稻密碼子優化的Cas9基因;KanR:卡那抗性標記;HygR:潮霉素抗性標記。Note: A:The entry vector pENTR4-sg. B:The destination vector pUbi-Cas9. OsU6: Rice U6 promoter. Zmubi-p: Maize Ubi Promoter. LB: Ti vetor left border. LB: Ti vetor right border. OsCas9:Rice codon optimized cas9. KanR: Kanamycin resistant gene. HygR: Hygromycin resistant gene. 圖1 基因編輯載體結構Fig.1 Structure of gene editing vectors
1.4 農桿菌轉化
采用凍融法[20]轉化農桿菌。用CaCl2法處理EHA105農桿菌菌株,制備感受態。將5 μg構建好的CSMAP65-3A和CSMAP65-3B質粒分別加入感受態細胞中,冰浴 30 min后液氮迅速冷凍3 min,37℃水浴5 min,加入1 mL農根農桿菌培養基(yeast mannitol medium, YEB)液體培養基,置28℃轉速200 r·min-1搖床恢復培養3 h。取200 μL 培養物,在YEB固體培養基(卡那霉素+利福平)上篩選培養,于28℃培養3 d后,挑取單克隆菌落進行PCR鑒定。
1.5 水稻轉化
選用300粒日本晴種子,去殼后用75%乙醇表面消毒1 min,次氯酸鈉溶液消毒30 min,移至營養肉湯(nutrien broth, NB)培養基(含2 mg·L-1的2,4-二氯苯氧乙酸2,4-D),26℃暗培養2周,挑選生長狀況好的愈傷用作轉化外植體;制備純度高,生活力強的EHA105工程菌液,浸染水稻愈傷后,在25℃暗條件下共培養3 d;脫菌后在含有50 mg·L-1潮霉素的篩選培養基上光照培養14 d左右(光照強度為13 200 lx,溫度為32℃);將預分化的愈傷轉至分化培養基上光照培養1個月左右(光照強度為13 200 lx,溫度為32℃),得到抗性轉基因植株;用1/2 MS培養基生根壯苗獲得T0代植株。
1.6 基因編輯情況分析
用CTAB法[21]提取T0代轉基因植株葉片DNA,通過檢測載體上抗潮霉素基因(HPTⅡ)確定轉基因陽性植株。用HPTⅡ特異引物Hyg-F(T C T C C G A C C T G A T G C A G C T C T CG)和Hyg-R(G C A C T G A C G G T G T C G T C C A T C A C AG)進行PCR檢測。PCR擴增10 μL體系如下:模板DNA 1 μL,10×Buffer 1 μL,dNTP 0.4 μL,引物(F+R)各0.5 μL,Taq酶0.05 μL,ddH2O 6.55 μL。PCR程序如下:94℃預變性5 min;94℃變性30 s,55℃退火30 s,72℃延伸45 s,27個循環;72℃終延伸10 min。
針對靶位點附近序列設計引物,對轉基因陽性植株的OsMAP65-3.1和OsMAP65-3.2進行克隆及測序分析。采用高保真酶KOD-Fx酶(日本東洋紡公司TOYOBO生產)進行PCR反應,KOD酶錯配率低,可提高試驗結果的可靠性。PCR擴增20 μL體系如下:模板DNA 2 μL,2×Buffer 10 μL,dNTP 1 μL,引物(F+R)各1 μL,KOD酶0.5 μL,ddH2O 4.5 μL。PCR程序如下:94℃預變性5 min;94℃變性30 s,60℃退火30 s,68℃延伸1 min,30個循環;68℃終延伸10 min。
2 結果與分析
2.1 水稻MAP65-3s同源蛋白分析
水稻中MAP65-3基因的來源用AtMAP65-3氨基酸序列在美國國家生物技術信息中心(National Center for Biotechnology Information,NCBI)數據庫進行比對,證實水稻中的2個基因(LOC_Os01g49200和LOC_Os05g47970)與AtMAP65-3同源性最高,同源性分別高達55%和53%(圖2),且2個基因之間的同源性高達74%。LOC_Os01g49200(OsMAP65-3.1)基因包含13個外顯子,蛋白質編碼區(coding sequence, CDS)全長2 070 bp,編碼689個氨基酸;而LOC_Os05g47970(OsMAP65-3.2)同樣包含13個外顯子,CDS全長1 989 bp, 編碼662個氨基酸。
植物MAP65蛋白包含一個相對保守的N端和一個保守性較差的C端,C端又分為C1和C2區,靠近C端的C2區保守性最差。N端是蛋白質形成二聚體所必需的功能結構域,C端是微管結合結構域[21]。進一步的比對結果也證明OsMAP65-3.1、OsMAP65-3.2與AtMAP65-3在N端(均為61%)和C1區同源性較高(分別64%和65%),而在C2區同源性相對較低(42%和35%)。
2.2 OsMAP65-3s sgRNA靶位點設計
對OsMAP65-3.1和OsMAP65-3.2編輯,依據靠近5′端和高特異性的原則,各設計了2個20 bp的靶位點,分別為CS3.1-1和CS3.1-2、CS3.2-1和CS3.2-2,序列信息見表1。CS3.1-1和CS3.1-2分別位于OsMAP65-3.1基因CDS的42~61和161~180;CS3.2-1和CS3.2-2分別位于OsMAP65-3.2基因CDS的221~240和468~487。4個靶位點均在對應基因的N端區域,編輯易產生無功能蛋白。此外,這些靶位點的特異性非常高,每一個靶位點的20個堿基序列在水稻基因組中都是唯一的;為考慮脫靶因素產生其他編輯,僅用12堿基作為靶序列進行基因組比對分析,每個靶位點也是唯一的;以8堿基作為靶序列的位點也較少,都在700之下。分析結果表明本研究所設計的靶位點特異性極高,脫靶概率極低,可特異編輯并產生無功能的OsMAP65-3.1和OsMAP65-3.2。
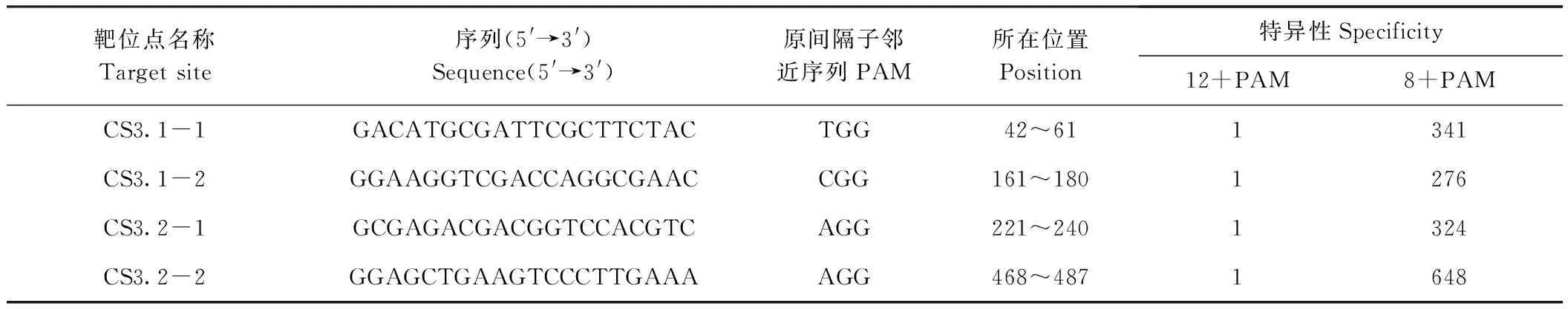
表1 OsMAP65-3s靶位點相關信息Table 1 The information of target sites for OsMAP65-3s
2.3 OsMAP65-3.1和OsMAP65-3.2雙基因編輯載體構建
由于OsMAP65-3.1和OsMAP65-3.2同源性較高,并均定位于成膜體中線,可能具有相似的功能。為避免兩者的功能冗余現象,將兩個基因的sgRNA構建至同一載體上,以獲得單基因和雙基因同時編輯的植株。為構建雙基因編輯載體,根據入門載體pENTR-4sg特征,利用單鏈合成和退火復性形成雙鏈,將靶位點CS3.1-1 和CS3.2-1 序列帶上BtgZ I酶切后的粘性末端序列,CS3.1-2和CS3.2-2序列帶上BsaI酶切后的粘性末端序列(圖3-A、B)。通過酶切連接法先將CS3.2-1和CS3.1-1分別整合到PENTR4-sg 載體BtgZ I位點,再將CS3.1-2和CS3.2-2 分別整合到對應中間載體的BsaI位點,構建入門載體PENTR4-sgMAP65-3A和PENTR4-sgMAP65-3B。PENTR4-sgMAP65-3A包含靶向CS3.2-1和CS3.1-2的sgRNA,PENTR4-sgMAP65-3B則包含靶向CS3.1-1 和CS3.2-2 的sgRNA。采用PCR技術對兩個入門載體陽性克隆檢測,前者用OsMAP65-3.2-pENTR-sg4-OsMAP65-3.1-2-R1作為引物,后者用OsMAP65-3.1-1-F1/OsMAP65-3.2-2-R1作引物(表2)。結果表明所檢測的菌落均能擴增出片段,說明兩個靶位點均已同時整合到載體上,陽性菌落測序結果也證實了入門載體構建成功。通過LR重組反應將對應片段整合到目標載體pUbi-Cas9,形成兩個雙基因編輯載體,分別為CSMAP65-3A和CSMAP65-3B。雙基因編輯載體中包含OsU6.1和OsU6.2啟動子指導的OsMAP65-3.1sgRNA和OsMAP65-3.2sgRNA,以及玉米Ubi啟動子指導的Cas9基因等重要結構(圖2-C、D),確保OsMAP65-3.1sgRNA、OsMAP65-3.2sgRNA和Cas9在水稻中表達。

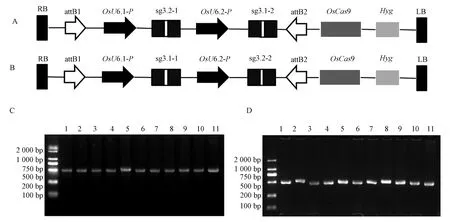
注: A: CSMAP65-3A載體結構圖;B: CSMAP65-3B載體結構圖;C:CSMAP65-3A載體菌落檢測圖; D: CSMAP65-3B載體菌落檢測圖。OsU6:水稻U6啟動子; LB:Ti質粒左邊界;RB:Ti質粒右邊界;OsCas9:水稻密碼子優化的Cas9基因; Hyg:潮霉素抗性標記。Note: A: Structure of CSMAP65-3A. B: Structure of CSMAP65-3B. C: Positive clone selection for CSMAP65-3A by PCR. D: Positive clone selection for CSMAP65-3B by PCR. OsU6: Rice U6 promoter. LB: Ti vetor left border. RB: Ti vetor right border. OsCas9: Rice codon optimized Cas9. Hyg: Hygromycin resistant gene.圖3 OsMAP65-3s雙基因編輯載體構建Fig.3 Construction of double-gene-editing vectors for OsMAP65-3s
2.4 轉基因植株鑒定
用含有OsMAP65-3s雙基因編輯載體(CSMAP65-3A、CSMAP65-3B)的EHA105菌株制備的工程菌液轉化水稻,分別獲得17和21株轉基因植株。用篩選標記抗潮霉素基因HPTII特異引物Hyg-F和Hyg-R對轉基因植株進行PCR鑒定,以載體質粒為模板做陽性對照,野生型日本晴DNA為模板做陰性對照。其中CSMAP65-3A載體獲得16株陽性轉基因植株,僅有#5株系為陰性植株(圖4-A);而CSMAP65-3B載體也僅有#5植株表現為陰性,其他20株均為陽性植株(圖4-B)。鑒定結果表明兩個載體轉基因植株的轉化效率均較高,可獲得足夠的轉基因株系供后續基因編輯分析。
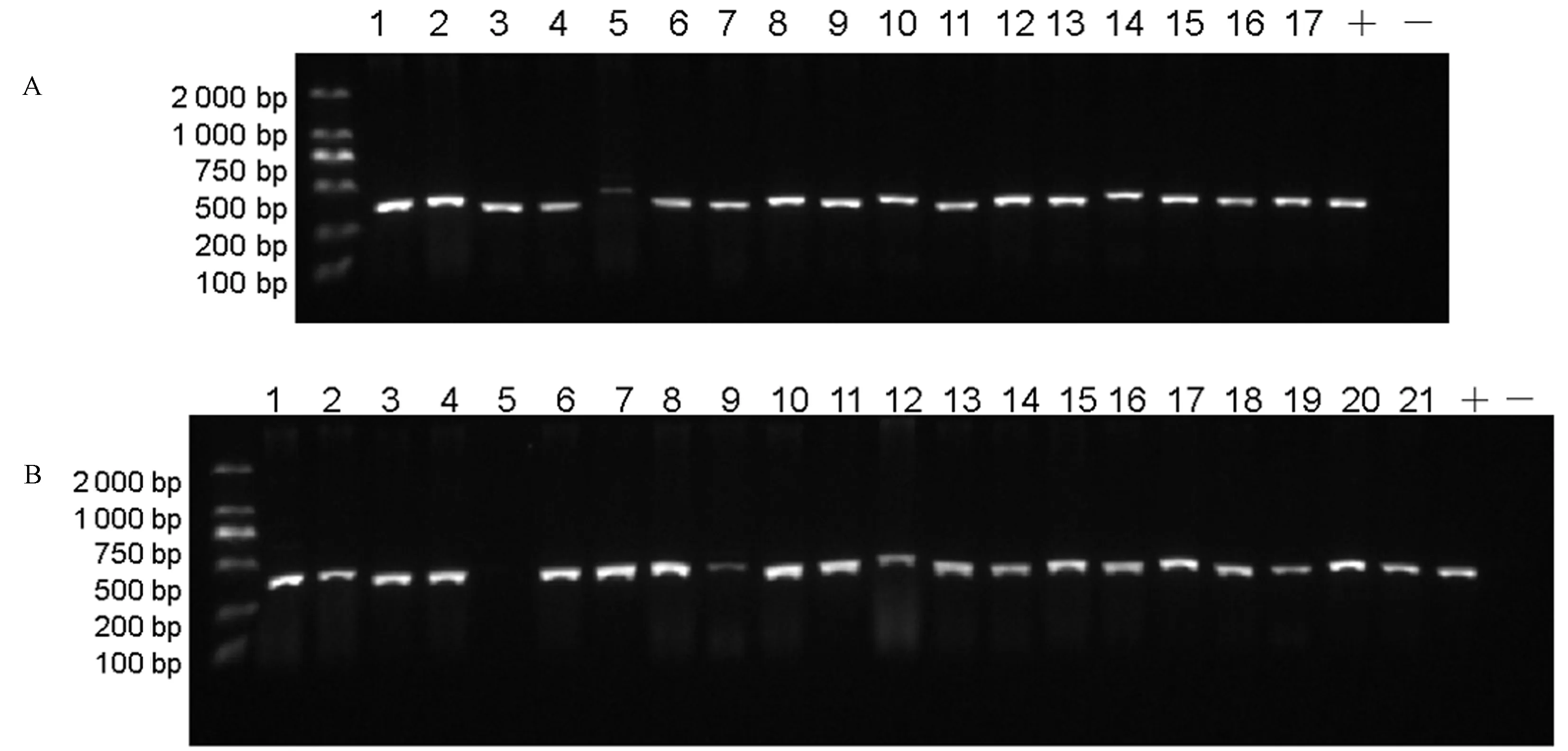
注:A: 轉CSMAP65-3A植株HPT Ⅱ鑒定圖,1-17泳道代表CSMAP65-3A對應的17株轉基因植株; B:轉CSMAP65-3B植株HPT Ⅱ鑒定圖,1-21代表CSMAP65-3B對應的21株轉基因植株。“+”代表以載體質粒為模板的陽性對照,“-”代表以野生型日本晴為模板的陰性對照。Note: A: HPT Ⅱ identification in CSMAP65-3A transgenic plants, 1-17 lane represents 17 transgenic plants corresponding to CSMAP65-3A. B: HPT Ⅱ identification in CSMAP65-3B transgenic plants. 1-21 represents 21 transgenic plants corresponding to CSMAP65-3B. ‘+’ represents positive control with vector plasmid as template. ‘-’ represents negative control with wild-type Nipponbare as template.圖4 轉基因株系潮霉素抗性基因HPT ⅡPCR鑒定Fig.4 Identification of HPT Ⅱ in transgenic lines by PCR
2.5 靶位點序列克隆
為分析轉基因植株中OsMAP65-3.1和OsMAP65-3.2基因編輯情況,用高保真酶克隆靶位點附近片段。由于OsMAP65-3.1基因的2個靶位點(CS3.1-1和CS3.1-2)在該基因DNA序列位置距離僅99 bp,可共用一對引物克隆目標片段,即SQ-OsMAP65-3.1-F/SQ-OsMAP65-3.1-R;而OsMAP65-3.2 2個靶位點(CS3.2-1和CS3.2-2)在染色體位置相差857 bp,克隆需設計不同引物,分別為SQ-OsMAP65-3.2-1-F/SQ-OsMAP65-3.2-1-R和SQ-OsMAP65-3.2-2-F/SQ-OsMAP65-3.2-2-R(表3)。提取CSMAP65-3A和CSMAP65-3B轉基因株系DNA,用對應引物對靶位點序列進行克隆,通過PCR反應獲得所有株系對應位置的OsMAP65-3.1和OsMAP65-3.2片段,用于測序及編輯情況分析。
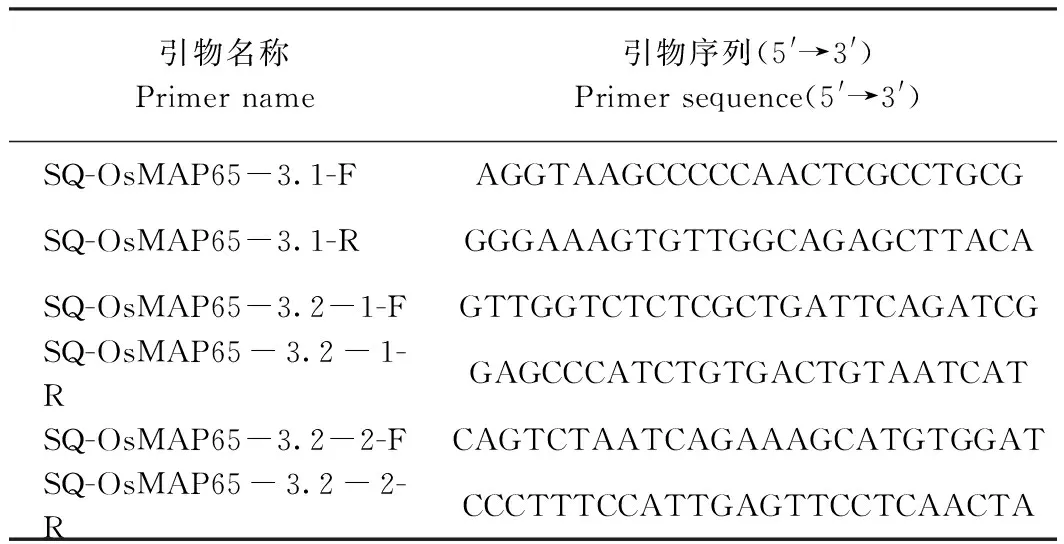
表3 OsMAP65-3s各靶位點片段克隆引物Table 3 Primers of target sites for OsMAP65-3s
2.6 OsMAP65-3s編輯情況分析
以野生型序列作為參考,通過測序峰圖讀取,可分析出每條染色體被編輯的情況。對CSMAP65-3A、CSMAP65-3B轉基因株系中各靶位點附近序列的測序峰圖進行分析,重點考查正常峰圖結果(單峰)是否存在個別位點的套峰(雜合)以及雙染色純合編輯情況,雙峰結果是一條染色體被編輯還是兩條染色體都被編輯(不同編輯形式)。如CSMAP65-3A轉基因#3植株和CSMAP65-3B轉基因#3植株,其OsMAP65-3.1與OsMAP65-3.2相應靶位點附近序列測序結果均表現為雙峰(圖6)。在CSMAP65-3A-#3中,OsMAP65-3.1兩條染色體均被編輯,其中一條缺失2個堿基,氨基酸發生移碼并提前終止,而另外一條染色體發生單堿基替換,導致其保守區氨基酸發生改變,可能會影響蛋白功能,需后期繼續研究;OsMAP65-3.2同樣有兩條染色體均被編輯,其中一條插入1個堿基,導致氨基酸移碼并提前終止,另外一條染色體發生3堿基缺失,在較不保守區域缺失1氨基酸,推測可能不影響蛋白功能。在CSMAP65-3B-#3株系的OsMAP65-3.1與OsMAP65-3.2基因在兩條染色體均被編輯,兩者編輯形式分別是缺失3堿基/插入1堿基和插入2堿基/缺失4堿基、插入1堿基(圖6)。
注:CSMAP65-3A 17株轉基因植株中OsMAP65-3.2-1靶位點區域片段克隆(A)和OsMAP65-3.1-2靶位點區域片段克隆(B);CSMAP65-3B 21株轉基因植株中OsMAP65-3.1-1靶位點區域片段克隆(C)和OsMAP65-3.2-2靶位點區域片段克隆(D)。Note:Cloning of OsMAP65-3.2-1 target region fragment (A) and cloning of OsMAP65-3.1-2 target region fragment (B) in 17 CSMAP65-3A transgenic plants. Cloning of OsMAP65-3.1-1 target region fragment (C) and cloning of OsMAP65-3.1-2 target region fragment (D) in 21 CSMAP65-3A transgenic plants. 圖5 轉基因植株中OsMAP65-3s靶位點區域片段克隆Fig.5 Cloning of target sites for OsMAP65-3s in transgenic plants
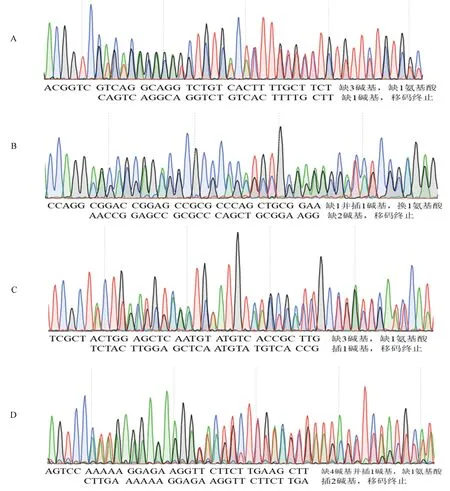
注:CSMAP65-3A轉基因3號株系OsMAP65-3.2-1位點編輯情況(A)和OsMAP65-3.1-2位點編輯情況(B);CSMAP65-3B轉基因3號株系OsMAP65-3.1-1位點編輯情況(C)和OsMAP65-3.2-2位點編輯情況(D)。Note:The edition of OsMAP65-3.2-1 site (A) and OsMAP65-3.1-2 site (B) in CSMAP65-3A transgenic plants (#3). The edition of OsMAP65-3.1-1 site (C) and OsMAP65-3.2-2 site (D) in CSMAP65-3B transgenic plants (#3).圖6 部分轉基因植株中OsMAP65-3s靶位點編輯分析Fig.6 The edition of target sites for OsMAP65-3s in transgenic plants
對全部轉基因陽性植株進行分析(表4、5),發現在16個CSMAP65-3A陽性轉基因株系中,有3個株系發生了基因編輯,其中2株的兩個基因均發生編輯;其中2個單株中OsMAP65-3.1-2位點被編輯(3個編輯事件,2株都有一條染色體為移碼終止),編輯效率為9.38%;3個株系OsMAP65-3.2-1位點被編輯,其中2株兩條染色體均被編輯,包含5個編輯事件,4個為移碼終止,編輯效率為15.62%。在20株CSMAP65-3B的轉基因陽性材料中,9株發生編輯,其中4株的OsMAP65-3.1-1位點被編輯,其中3株兩條染色體同時被編輯,共7個編輯事件,4個移碼終止,編輯效率為17.50%;而OsMAP65-3.2-2發生編輯的9個株系全部為兩條染色體同時編輯,移碼終止的編輯事件高達13次,編輯效率為45.00%。

表4 CSMAP65-3A轉基因植株中OsMAP65-3s編輯情況Table 4 The edition of OsMAP65-3s in CSMAP65-3A transgenic plants

表5 CSMAP65-3B轉基因植株中OsMAP65-3s編輯情況Table 5 The edition of OsMAP65-3s in CSMAP65-3B transgenic plants
3 討論
OsMAP65-3.1和OsMAP65-3.2是水稻中細胞質分裂關鍵因子MAP65-3的2個同源蛋白,本研究團隊用煙草葉片瞬時分裂系統已經證明OsMAP65-3.1和OsMAP65-3.2均定位于成膜體中線[22],可能均參與調控水稻細胞質分裂,并可能存在功能冗余。本研究通過CRISPR/Cas9技術對水稻OsMAP65-3.1和OsMAP65-3.2進行編輯實現定點突變,創制osmap65-3s突變體,獲取雙基因突變體是分析其功能的重要前提,有利于闡明水稻有絲分裂調控機理。創制雙基因突變體有兩種方案:第一種方案需先分別獲得單突變植株,然后進行雜交以獲得雙突植株。如擬南芥中的CPK10/CPK30雙突變體是以純合單突變體作為親本進行人工雜交,繁衍至F2代從群體中分離出的純合雙突植株[23]。而水稻突變體資源相對較少,用該方案創制雙突變體周期較長。需先用基因編輯技術創制兩個基因的單突變體,再通過雜交將兩個突變基因聚合到F1,自交F2中篩選到雙突變體,經歷3~6個世代才可獲得雙突變體。第二種方案是將兩個基因的靶位點對應的sgRNA構建到同一個載體上,可對兩個基因進行編輯。由于單個位點的編輯具有一定的概率,轉化植株會出現不同的編輯組合,因此可獲得單基因突變體和雙基因突變體。該方案節省大量的時間,僅需1~2個世代就可獲得雙突變體。如水稻中酯酶家族成員同源性較高的成員OsGELP110與OsGELP115,將二者sgRNA構建到同一載體轉化后,迅速獲得osgelp110單突變體、osgelp115單突變體和osgelp110/osgelp115雙突變體,此外單獨敲除OsGELP110或OsGELP115無明顯表型,而雙突變體花粉外壁結構會發生變化,導致雄性不育[24]。本研究將OsMAP65-3.1和OsMAP65-3.2的不同靶位點整合成兩個不同的雙基因編輯載體CSMAP65-3A和CSMAP65-3B,轉化后獲得的T0代轉基因材料中,已經獲得多個osmap65-3.2、osmap65-3.2/OsMAP65-3.1(+/-)和OsMAP65-3.2(+/-)/OsMAP65-3.1(+/-)植株(表4、5),可通過對應株系自交T1代獲得osmap65-3.1和osmap65-3.1/osmap65-3.2。此外,本研究中雙基因編輯植株比例較高,16個CSMAP65-3A陽性植株中有2株,比例12.5%,而20個CSMAP65-3B的轉基因株系中有4株兩個基因同時被編輯,比例高達20%。而李星坤等[25]用雙基因編輯載體對擬南芥中糖基轉移酶同工酶基因UGT84A1和UGT84A2同時進行編輯,雙突植株比例僅為4.76%。因此,本研究選用靶位點和編輯載體可快速、高效地創制OsMAP65-3s雙突變體材料。
本研究針對OsMAP65-3.1和OsMAP65-3.2均設計了2個不同的靶位點,且都獲得了水稻編輯植株。其中OsMAP65-3.1-1位點編輯效率為17.50%,OsMAP65-3.1-2位點編輯效率為9.38%;OsMAP65-3.2-1位點編輯效率為15.62%,OsMAP65-3.2-2編輯效率為45.00%。表明同一基因的不同靶位點會影響編輯效率,與已報道的蛋白激酶MPK5一致,載體的編輯效率與不同的靶序列相關性較大[26]。因此,為確保目標基因的高效編輯,應選用2個及以上的靶位點。但是,相比OsMAP65-3.2以及前人報道的水稻OsGRAS39基因[27],本研究中OsMAP65-3.1 2個靶位點編輯效率均較低,一定程度上可排除靶位點選擇的因素。由于OsMAP65-3.1基因在水稻中的功能非常重要,該基因被編輯的細胞在水稻轉化過程中難以存活,導致成苗率低,且大部分編輯事件無法檢測到。因此,OsMAP65-3.1編輯植株難以獲得。MAP65-3是調控植物細胞板的合成及細胞質分裂的關鍵因子[22,28]。MAP65-3通過二聚化及磷酸化調控其蛋白活性決定了成膜體中間區域空間的寬窄和成膜體的擴張;該基因的擬南芥突變體成膜體中間區域變的更寬,細胞質分裂失敗的頻率明顯增加[29-30]。因此,水稻OsMAP65-3s異常可能會影響細胞分裂及轉化苗的形成。
4 結論
本研究通過CRISPR/Cas9技術,對水稻中2個與細胞質分裂相關的OsMAP65-3基因進行敲除創制突變體材料。為避免基因功能冗余,本試驗在一個載體中同時設計了OsMAP65-3.1與OsMAP65-3.2敲除靶位點,并且設計了2個雙敲載體來提高編輯效率。在獲得的2個轉基因群體中,陽性植株比例均高于90%,所設計的4個靶位點都出現編輯事件,其中OsMAP65-3.1的2個靶位點的編輯效率分別為17.50%和9.38%,OsMAP65-3.2編輯效率分別為15.62%和45.00%。T0代已經獲得4種突變體材料,分別是osmap65-3.2、osmap65-3.2/OsMAP65-3.1(+/-)和OsMAP65-3.2(+/-)/OsMAP65-3.1(+/-), 后面仍需通過自交獲得osmap65-3.1單突變體及osmap65-3.1/osmap65-3.2 雙突變體。此外,在獲得的編輯植株中,OsMAP65-3.2在T0代已經出現2條染色體均被提前終止的突變體,而OsMAP65-3.1只存在突變雜合,表明OsMAP65-3.1與OsMAP65-3.2的功能可能存在差異,且OsMAP65-3.1在水稻有絲分裂過程中可能更為重要。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
