甘藍(lán)型油菜黃化轉(zhuǎn)綠型突變系的光合特性
2022-03-17 08:09:52張耀文關(guān)周博李少欽侯君利董育紅張文學(xué)田建華
西北農(nóng)業(yè)學(xué)報 2022年3期
張耀文,關(guān)周博,李少欽,侯君利,董育紅,張文學(xué),田建華
(陜西省雜交油菜研究中心,國家油料作物改良中心 陜西油菜分中心,陜西楊凌 712100)
根據(jù)葉色變異后能否轉(zhuǎn)綠可將作物的葉色突變體分為轉(zhuǎn)綠和非轉(zhuǎn)綠兩種類型[1],轉(zhuǎn)綠型突變體又可分為白化轉(zhuǎn)綠型和黃化轉(zhuǎn)綠型[1-2],其中黃化轉(zhuǎn)綠型突變體對研究作物的葉綠體發(fā)育機(jī)制、光合機(jī)理及進(jìn)行高光效育種均具有重要價值。在水稻[3-12]、小麥[13-14]、大豆[15-16]、大麥[17-18]、棉花[19-22]等作物上均已發(fā)現(xiàn)黃化轉(zhuǎn)綠型突變體。前人的研究表明,不同作物(來源)的黃化轉(zhuǎn)綠型突變體在光合色素含量及組成、光合能力、葉綠素?zé)晒鈪?shù)等方面表現(xiàn)出不同甚至相反的結(jié)果:突變體的葉綠素a含量降低[4-5,16],葉綠素b含量降低[5,8,16]甚至不存在[7],突變體的葉綠素a/b比值升高[20]、降低[21],突變體的凈光合效率降低[6,8,10,14]、升高[4,20]或無差異[9],突變體的葉綠素?zé)晒鈪?shù)降低[5,10,12,16]、升高[4]。因此尚難以全面理解葉色突變體的生理生化特性及對光合特性影響的機(jī)制。
相對于水稻、小麥等作物,目前油菜中發(fā)現(xiàn)的黃化轉(zhuǎn)綠型葉色突變體較少,僅董遵等[23]、趙云等[24]、肖華貴等[25]、張澤斌等[26]在甘藍(lán)型油菜、李瑋等[27]在芥菜型油菜中發(fā)現(xiàn)了黃化轉(zhuǎn)綠型突變體。因葉色突變的來源、研究方法不同所得結(jié)論有較大差異,尚需發(fā)現(xiàn)更多的突變體進(jìn)行廣泛深入的研究。同時,前人多是在黃化期間的某個時間節(jié)點對油菜黃化材料進(jìn)行研究,所得結(jié)論也不能全面解析黃化轉(zhuǎn)綠型突變系的光合特性的動態(tài)變化。筆者研究團(tuán)隊于2013年在田間發(fā)現(xiàn)甘藍(lán)型油菜可轉(zhuǎn)綠型黃化突變株,具體表型性狀表現(xiàn)為:子葉綠色,3種類型真葉(長柄葉、短柄葉、無柄葉)初生的幼嫩葉均為黃色,隨著發(fā)育由葉緣向葉柄處逐漸轉(zhuǎn)綠最終恢復(fù)正常葉色。新生的主莖、花蕾、角果也不同程度黃化隨后逐漸復(fù)綠。其表型有別于前人的報道,是一種甘藍(lán)型油菜的新型葉色變異材料。經(jīng)過多代篩選已獲得可穩(wěn)定遺傳的黃化轉(zhuǎn)綠型突變系(Yellow green-revertible line,Ygr)和正常葉色的近等系(Near isogenic line,Nil)。油菜葉色近等系因與突變系遺傳背景的高度一致,是研究油菜葉色變異光合特性的理想材料。本試驗以黃化轉(zhuǎn)綠型突變系(Ygr)和正常葉色的近等系(Nil)為材料,對多個光合器官的多種光合特性的變化過程進(jìn)行比較,以期能夠為研究油菜的光合特性機(jī)理提供有益的參考。
1 材料與方法
1.1 試驗材料
陜西省雜交油菜研究中心選育的甘藍(lán)型油菜黃化轉(zhuǎn)綠型突變系(Ygr)和正常葉色的近等系(Nil)。試驗于2018-2019和2019-2020年在陜西省雜交油菜研究中心試驗田進(jìn)行,每個材料種植12行;田間管理按本中心統(tǒng)一要求進(jìn)行。
在五葉期(2019-10-10)、抽薹期(2020-03-23)、選擇剛展開第5葉位長柄葉(Longpetiole leaf,LPL)、倒1葉位的短柄葉(Shortpetiole leaf,SPL);初花期(2020-03-28)選擇主莖上的花蕾(Flower bud,F(xiàn)B)、盛花期(2020-04-02) 選擇綠色莖稈(Main stalk skin,MSS),角果期(2020-04-14)選擇主莖第2~5位角果(Silique skin,SS)進(jìn)行測定。
1.2 測定項目及方法
1.2.1 光合色素 葉片葉綠素含量變化的測定:在五葉期、抽薹期分別標(biāo)記剛展開的長柄葉、短柄葉,每個材料標(biāo)記10株;用SPAD502葉綠素測定儀對已標(biāo)記的葉片進(jìn)行定點測量(葉綠素含量用SPAD值表示),此后每隔3 d重復(fù)測定一遍,取平均數(shù)進(jìn)行統(tǒng)計。按照新葉抽出時間劃分為前期(A1d-A6d)、中期(A9d-A12d)、后期(A12d以后)3個階段進(jìn)行比較分析。
光合色素含量變化:在五葉期、抽薹期、初花期、角果期對長柄葉、短柄葉、花蕾、綠色莖稈、主莖進(jìn)行取樣,剪碎混勻后稱取1 g樣品,按照張耀文等[28]測定的方法進(jìn)行測定。分別在5 d、10 d、15 d后在同樣部位取樣進(jìn)行測定。
1.2.2 光合面積 單葉片、角果面積生長變化的測定:在五葉期、抽薹期標(biāo)記剛展開的長柄葉、短柄葉,Ygr和Nil各標(biāo)記10株。對已標(biāo)記的葉片在固定部位進(jìn)行長、寬測定,按照曹櫟等[29]的系數(shù)回歸法計算單葉面積。此后每隔3 d重復(fù)測定一遍。葉片的生長階段劃分同上。在2019-03-25對Ygr和Nil主莖上所開的花進(jìn)行標(biāo)記(每個材料標(biāo)記10株),從2019-03-30開始測量角果的長、寬,按照張耀文等[30]的方法計算單個角果的面積。同上,每隔3 d重復(fù)測定一遍。將角果生長階段按照開花后日期劃分為前期(A1d-A10d)、中期(A13d-A16d)、后期(19 d以后)3個階段進(jìn)行比較。
單株綠葉面積、角果皮面積測定:在不同生長時期,Ygr和Nil各選取15株均勻一致的植株,按照張耀文等[28]的方法分別測定單株綠葉面積、角果皮面積。
1.2.3 氣體交換參數(shù) 不同光照條件下氣體交換參數(shù)的測定:在2019-10-16、2020-03-29、2020-04-12分別選擇第5葉位長柄葉(LPL)、倒1葉位短柄葉(SPL)、主序上的角果(第6~8位)用Li-6400便攜式光合作用測定系統(tǒng)進(jìn)行測定,測定方法同參考文獻(xiàn)[30]。
不同CO2濃度下氣體交換參數(shù)測定:在2019-10-17、2020-03-30、2020-04-13選擇分別選擇第5葉位LPL、倒1葉位SPL、主序上第6~8位的角果,用Li-6400便攜式光合作用測定系統(tǒng)進(jìn)行測定,測定方法同參考文獻(xiàn)[30]。
1.2.4 葉綠素?zé)晒鈪?shù) 在2019-10-16、2020-03-29、2020-04-13分別選擇第5葉位LPL、倒1葉位SPL、主序上第6~8位角果;應(yīng)用暗適應(yīng)夾子將葉片(角果)暗適應(yīng)2 h后,用便攜式調(diào)制葉綠素?zé)晒鈨xPAM-2500的熒光誘導(dǎo)動力學(xué)曲線進(jìn)行測定。
1.2.5 農(nóng)藝性狀和產(chǎn)量性狀 分別在苗期、苔期、花期、成熟期取10株代表性植株,按照DUS標(biāo)準(zhǔn)[31]測定葉片數(shù)、分枝數(shù)、株高、生物學(xué)產(chǎn)量等農(nóng)藝性狀,主花序長度、主花序角果數(shù)、總角果數(shù)、角粒數(shù)、千粒質(zhì)量、單株產(chǎn)量等產(chǎn)量性狀;取2年結(jié)果的平均值進(jìn)行比較。
1.3 數(shù)據(jù)處理
用Excel 2009初步整理后進(jìn)行統(tǒng)計分析。
2 結(jié)果與分析
2.1 光合色素的比較
2.1.1 葉片葉綠素含量變化 從圖1可看出,Ygr與Nil的葉綠素含量均隨時間變化而升高。在出葉后1~16 d,Ygr的2類葉片(LPL、SPL)葉綠素含量的增長幅度(31.01、 44.03)均大于Nil(10.87、11.83)。在不同時間點與Nil相比較,Ygr的2類葉片在A1d~A13d均低于Nil,且2者間差異的大小均為A1d(-70.34%、 -60.40%)>A4d(-53.01%、-50.86%)>A7d (-33.35%、 -27.96%)>A10d (-16.86%、 -10.58%)>A13d(-5.11%、-3.45%)>A16d(0.07%、 0.17%),Ygr的2類葉片的葉綠素含量在前期(A1d~A7d)分別平均低52.23%、48.74%,在中期(A10d~A13d)分別平均低 10.99%、6.82%。

圖柱上的大、小寫字母分別表示不同柱間差異達(dá)到0.01或0.05的顯著水平,下同
從圖2可看出,Ygr的2類葉片(LPL、SPL)葉綠素含量的增長速率為“先升高再降低”而Nil則為“逐步降低”。與Nil的相比較,Ygr的2類葉片葉綠素含量增長速率在A1d~A16d分別高156.62%~727.03%、118.72%~4 900.00%。在不同時間階段相比較,Ygr與Nil的2類葉片間的差異均為A13d~A16d(727.03%、 4 900.00%)最大、A1d~A4d(156.62%、 118.72%)最小;在其余階段Ygr與Nil間的差異大小,LPL為A10d~A13d(396.38%)>A7d~A10d (350.67%)>A4d~A7d(252.34%)而SPL為A7d~A10d(727.10%)>A4d~A7d(360.65%)>A10d~A13d(221.09%)。
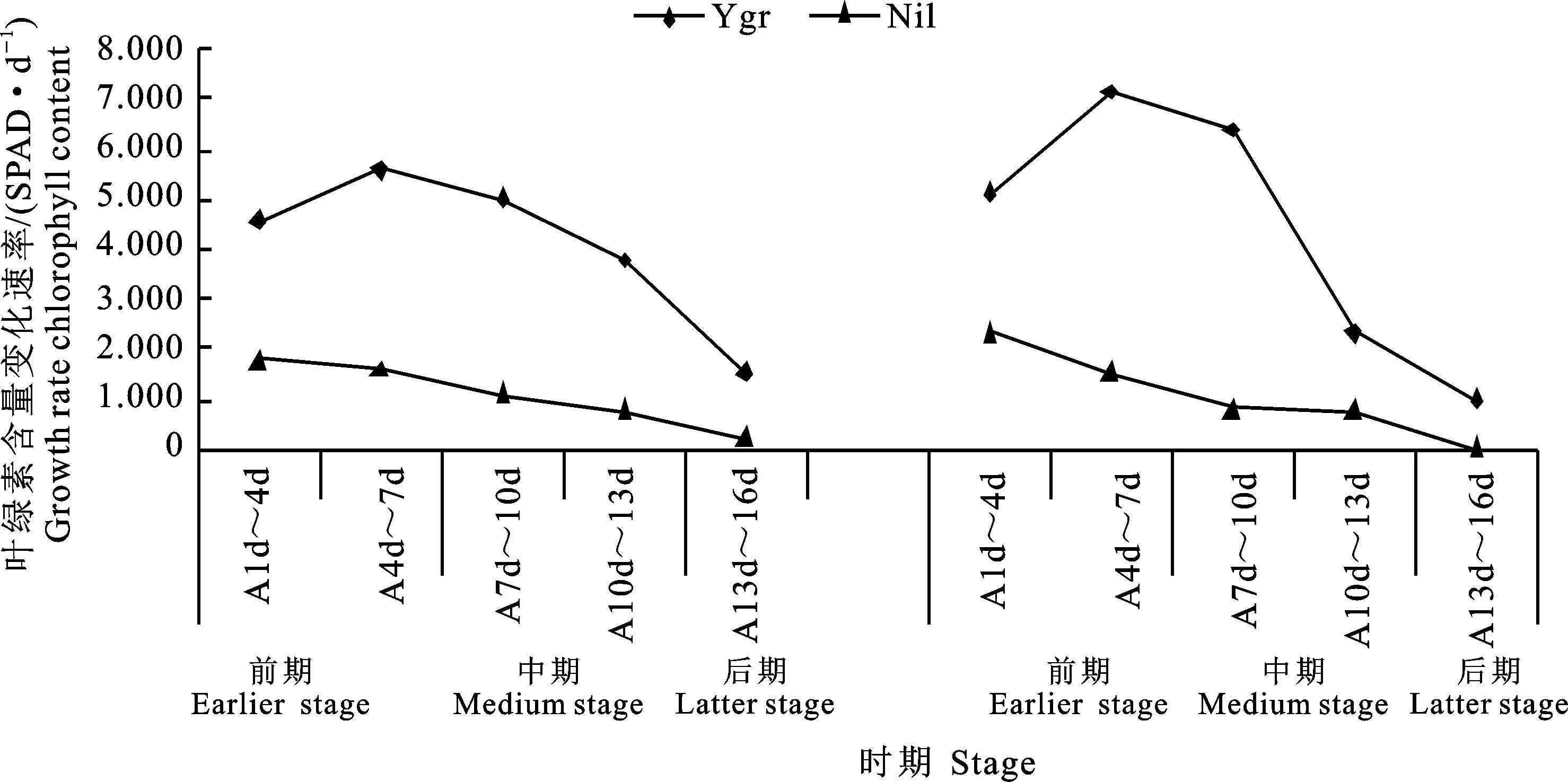
圖2 Ygr與Nil長柄葉、短柄葉(單葉)葉綠素含量增長速率變化的比較Fig.2 Comparison of growth rate of chlorophyll content changes of Ygr and Nil single leaf
2.1.2 光合色素含量的變化 從圖3可看出,在不同時間點與Nil相比較,在新葉展開后第1天(A1d),Ygr的5類光合器官的(LPL、SPL、MSS、FB、SS)的光合色素組分中Chla/b、Chla/(a+b)分別高5.81%~43.06%、1.50%~9.71%,而Chl(a+b)/Car、Chlb/(a+b)則分別低 12.50%~23.99%、4.07%~22.75%;在新葉展開后第5天(A5d),Ygr的2類葉片(LPL、SPL)的Chla/b、Chla/(a+b)分別高22.66%、 25.56%,5.87%、6.46%,而Chl(a+b)/Car、Chlb/(a+b)則分別低17.35%、5.68%, 13.69%、5.21%;在新葉展開后第10天(A10d)、15天(A15d),Ygr與Nil光合器官間的光合色素含量則無顯著差異。

圖3 不同時期不同光合器官光合色素含量的比較Fig.3 Comparison of photosynthetic pigments content in different photosynthetic organs
從圖4可看出,隨時間變化,Ygr的5類光合器官的4種光合色素[葉綠素a(Chla)、葉綠素b(Chlb)、葉綠素a+b (Chl a+b)、類胡蘿卜素(Car)]含量與Nil之間的差異均逐步變小,在第15d則無顯著差異。與Nil相比,Ygr的4種光合色素間的降低程度為Chlb>Chl(a+b)>Chla>Car, 5類光合器官的降低程度為LPL>SPL>MSS>FB>SS。在不同時間點與Nil相比,Ygr的2類葉片(LPL、SPL)的4種光合色素含量的降低量為A1d(52.44%~72.66%、47.90%~ 66.84%)>A5d(41.10%~56.46%、18.72%~35.27%)>A10d(14.00%~18.00%、4.79%~6.47%)>A15(0.02%~0.71%、0.49%~ 0.72%)。與Nil相比,Ygr莖稈(MSS)的4種光合色素含量在A1d低26.02%~55.09%,在A5d僅Chlb、Chl(a+b)含量低8.45%、5.74%。與Nil相比,Ygr花蕾(FB)、角果(SS)的4種光合色素含量僅在A1d分別低17.43%~34.00%、 10.05%~24.50%,其余時間則無顯著差異。

圖4 不同光合器官光合色素含量的差異Fig.4 Differences of photosynthetic pigments content in different photosynthetic organs
2.2 氣體交換參數(shù)的比較
2.2.1 光響應(yīng)曲線參數(shù) 從圖5可看出,Ygr與Nil的3類光合器官(LPL、SPL、SS)的凈光合速率(Pn)均隨光照強(qiáng)度增加而提高。光照強(qiáng)度由0 μmol·mL-1提高到2 200 μmol·mL-1,Ygr的3類光合器官Pn的提高幅度 (19.26 μmol·m-2·s-1、22.19 μmol·m-2·s-1、 17.80 μmol·m-2·s-1)分別比Nil低17.17%、 18.79%、0.81%。在不同光強(qiáng)點的與Nil相比較,Ygr葉片(LPL、SPL)的Pn值在光照強(qiáng)度為0 μmol·mL-1、20 μmol·mL-1平均高 335.04%、318.40%,在50 μmol·mL-1~ 2 200 μmol·mL-1范圍內(nèi)平均低23.91%、28.95%。在不同光強(qiáng)點與Ni相比較,在0~100 μmol·mL-1、150 μmol·mL-1~2 200 μmol·mL-1范圍內(nèi)Ygr角果的Pn值分別平均低13.82%、 7.93%。
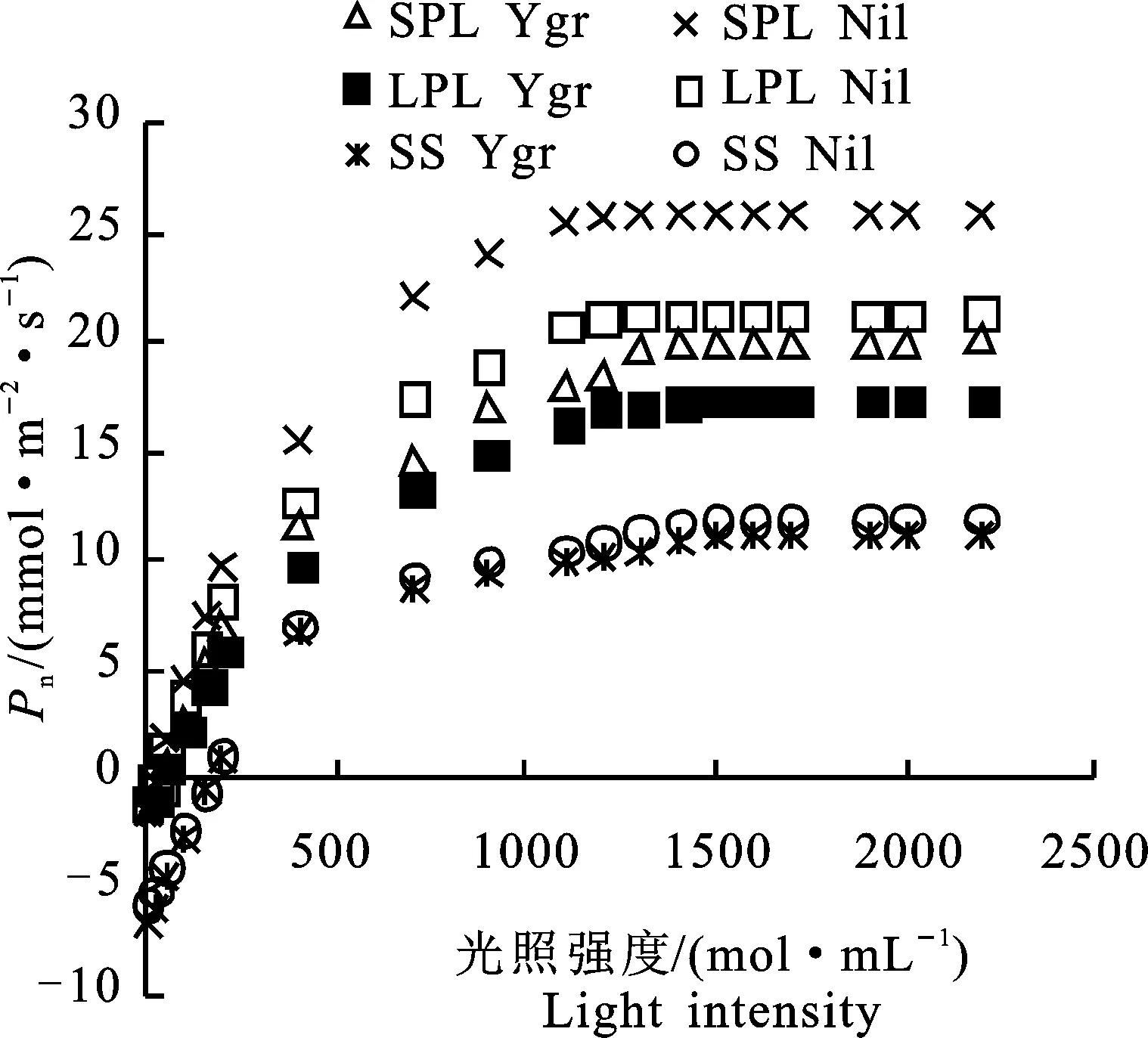
圖5 Ygr與Nil不同光合器官光曲線的比較Fig.5 Comparison of automatic light curves of different photosynthetic organs in Ygr and Nil
由表1可看出,與Nil相比較,Ygr2類葉片(LPL、SPL)的光飽和點(LSP)、光補(bǔ)償點(LCP)、光量子效率(LQE)、光呼吸速率(PR)高16.73%~95.93%。在光飽和點以上與Nil的平均氣體交換參數(shù)相比較,Ygr葉片的Pn、氣孔導(dǎo)度(Cond)、氣孔限制值(LS)、水分利用效率(WUE)低10.78%~34.58%,而胞間CO2濃度(Ci)、蒸騰速率(Trm)則高8.49%~18.30%。與Nil相比較,Ygr角果(SS)的光飽和點(LSP)、光量子效率(LQE)、光呼吸速率(PR)高4.52%~ 14.71%。在光飽和點以上與Nil的平均氣體交換參數(shù)相比較,Ygr角果的氣孔導(dǎo)度(Cond)、水分利用效率(WUE)低8.57%、9.15%。
表1 不同光合器官在不同光照條件下氣體交換參數(shù)的比較Table 1 Comparison of the gas exchanges photosynthetic parameters of different photosynthetic organs under different light conditions between Ygr and Nil
2.2.2 A-Ci曲線參數(shù) 從圖6可看出,Ygr與Nil的3類光合器官(LPL、SPL、SS)的Pn值隨CO2濃度的增加而提高。CO2濃度由0 μmol·mL-1升高到2 200 μmol·mL-1,Ygr 3類光合器官Pn的提高幅度(43.62 μmol·m-2·s-1、 46.20 μmol·m-2·s-1、40.45 μmol·m-2·s-1)分別比Nil低11.16%、16.61%、5.43%。在整個CO2濃度(10~ 2 200 μmol·mL-1)變化范圍內(nèi)與Nil相比較,Ygr的3類光合器官的Pn平均分別低17.57%、17.37%、5.26%。
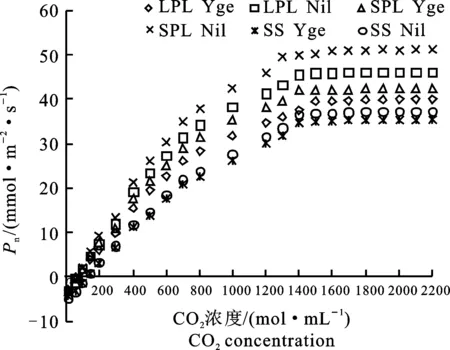
圖6 Ygr與Nil不同光合器官A-Ci曲線的比較Fig.6 Comparison of A-Ci curve of different photosynthetic organs in Ygr and Nil
由表2可看出,與Nil相比較,Ygr葉片(LPL、SPL)的CO2飽和點(CSP)、CO2補(bǔ)償點(CCP)、暗呼吸速率(DR)高5.09%~13.45%,Ygr角果(SS)的CSP、DR分別高4.59%、 8.31%,Ygr的3類光合器官(LPL、SPL、SS)的羧化效率(CE)則低6.82%~14.29%。在CO2飽和點以上與Nil的平均氣體交換參數(shù)相比較,Ygr的3類光合器官的Pn、Cond、Ls、WUE低 4.93%~50.00%,而Ci、Trm則高4.87%~ 20.32%。
表2 不同光合器官在不同CO2濃度條件下光合參數(shù)的比較Table 2 Comparison of photosynthetic characteristics of different photosynthetic organs under different CO2 concentration conditions in Ygr and Nil
2.3 葉綠素?zé)晒鈪?shù)的比較
從圖7、8可看出,與Nil的相比較,Ygr葉片(LPL、SPL)的初始熒光(F0)、PSⅡ調(diào)節(jié)性能量耗散的量子產(chǎn)額(YNPQ)、非光化學(xué)猝滅系數(shù)(NPQ)分別高16.39%、16.45%,10.00%、 20.27%,17.59%、21.67%;PSⅡ潛在活性 (Fv/Fm)、PSⅡ中光化學(xué)能量轉(zhuǎn)化的有效量子產(chǎn)量(YⅡ)、光化學(xué)猝滅系數(shù)(Qp)、表觀電子傳遞速率(ETR) 則分別低5.30%、4.61%, 14.29%、 12.77%,8.06%、7.29%,13.52%、 12.02%。Ygr角果(SS)的主要葉綠素?zé)晒鈪?shù)中僅YNPQ比Nil高10.64%。

圖7 Ygr與Nil不同光合器官A-Ci曲線的比較Fig.7 Comparison of chlorophyll fluorescence parameter of different photosynthetic organs in Ygr and Nil
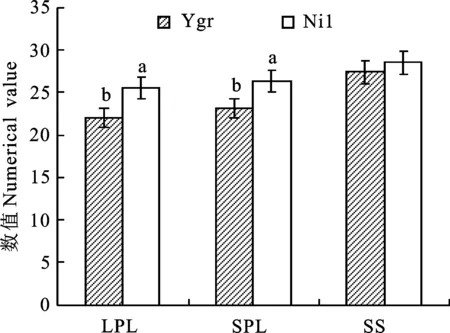
圖8 Ygr與Nil不同光合器官電子傳遞速率的比較Fig.8 Comparison of electron transport rate of different photosynthetic organs in Ygr and Nil
2.4 光合面積變化的比較
2.4.1 單葉面積變化 從圖9可看出,Ygr與Nil的LPL、SPL的單葉面積均隨時間變化而逐步升高,且均在展開后22 d(A22d)達(dá)到最大值(19.53、19.89、13.77、13.65 cm2)。與Nil相比較,Ygr的LPL、SPL在前期(A1d~A10d)分別平均低25.85%、24.23%。與Nil間的差異,LPL在第7天(39.68%)、SPL在第10天(37.62%)分別達(dá)到最大。與Nil相比較,Ygr在中期(A13d~A16d)平均低13.03%、 8.04%,在后期(A19d~A22d)平均低2.77%、 0.07%。
從圖10可看出,Ygr與Nil的單葉面積的增長速率均在前期逐步加大而在中-后期則逐步縮小。LPL單葉面積的增長速率,Ygr與Nil均在第7~10天的達(dá)到最大(2.20、3.20 cm2·d-1)。SPL單葉面積的增長速率,Ygr在第10~13天的達(dá)到最大(1.60 cm2·d-1),而NiL在第7~10天的達(dá)到最大(2.27cm2·d-1)。與Nil相比較,Ygr葉片(LPL、SPL)單葉面積的增長速率在前期(A1d~A10d)平均低48.48%、50.49%,在中、后期(A13d~A16d、A19d~A22d)則平均高 354.83%、549.68%。
2.4.2 單個角果面積變化 從圖9可看出,與Nil相比,Ygr的角果面積在前期(A1d~A10d)平均低14.15%,在中期(A13d~A16d)平均低 6.36%,在后期(A19d~A22d)則無顯著差異。Ygr與Nil間的差異在前期逐步加大(由 -3.83%~-20.12%)且在第10天達(dá)到最大 (-20.12%),在中、后期則逐步縮小(由 -9.06%~0.15%)。
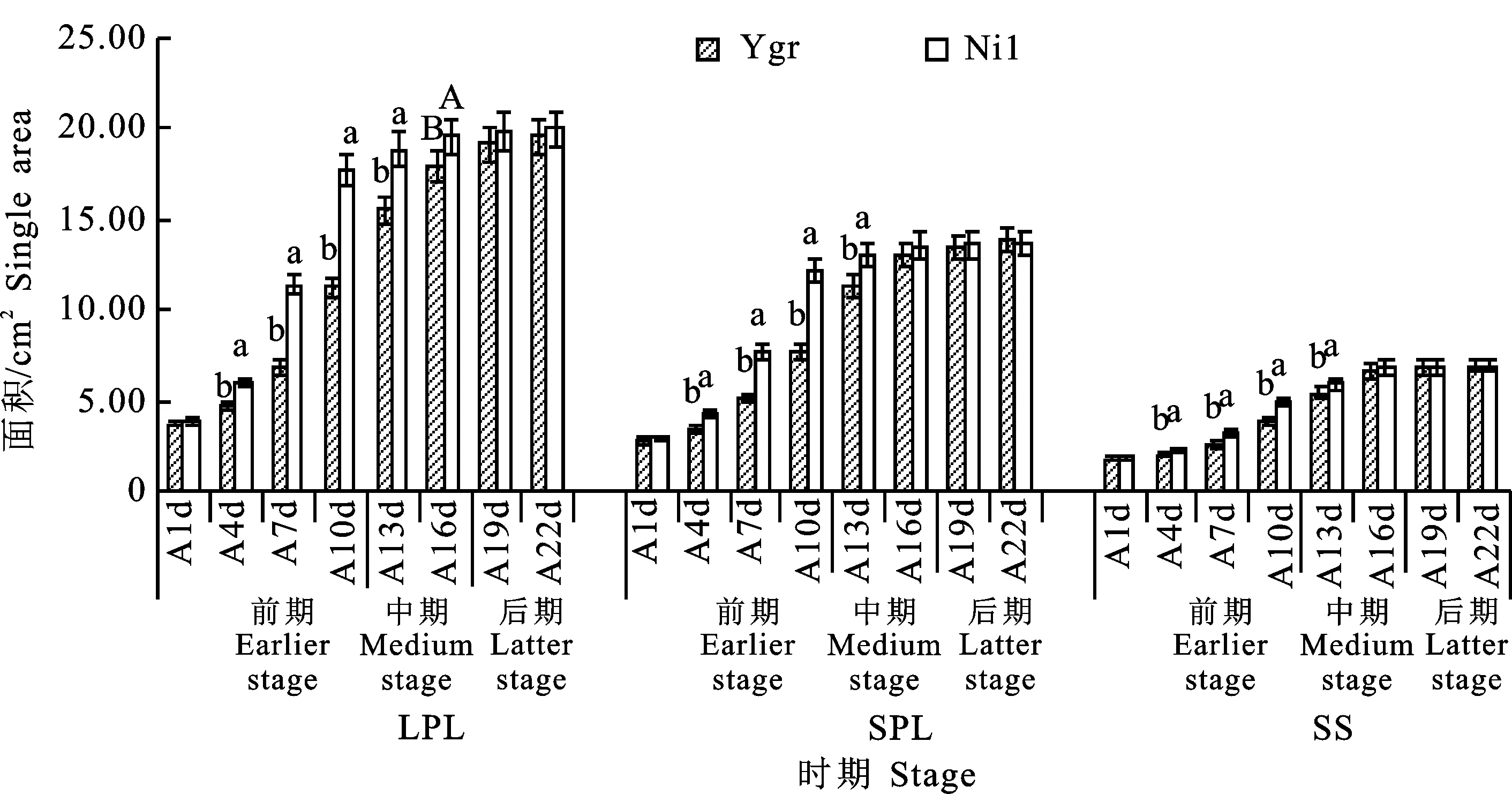
圖9 不同時期Ygr與Nil單葉面積和單個角果面積變化的比較Fig.9 Comparison of single changes of green leaf and single silique area per plant of Ygr and Nil at different stages
從圖10可看出,Ygr與Nil角果面積的增長速率均在前期(A1d~A10d)逐步加大(0.13~ 0.66 cm2·d-1、0.24~0.83 cm2·d-1)而在后期(A19d~A22d)均逐步縮小(0.17~0.04 cm2·d-1、0.02~0.01 cm2·d-1)。角果面積的增長速率,Ygr在A10d~13d(0.77cm2·d-1)、Nil在第7~10天的達(dá)到最大(0.83 cm2·d-1)分別達(dá)到最大。與Nil相比較,Ygr的角果面積增長速率在前期(A1d~A10d)平均低35.22%,而在中、后期(A13d~A16d、A19d~A22d)則分別平均高 30.98%、800%。
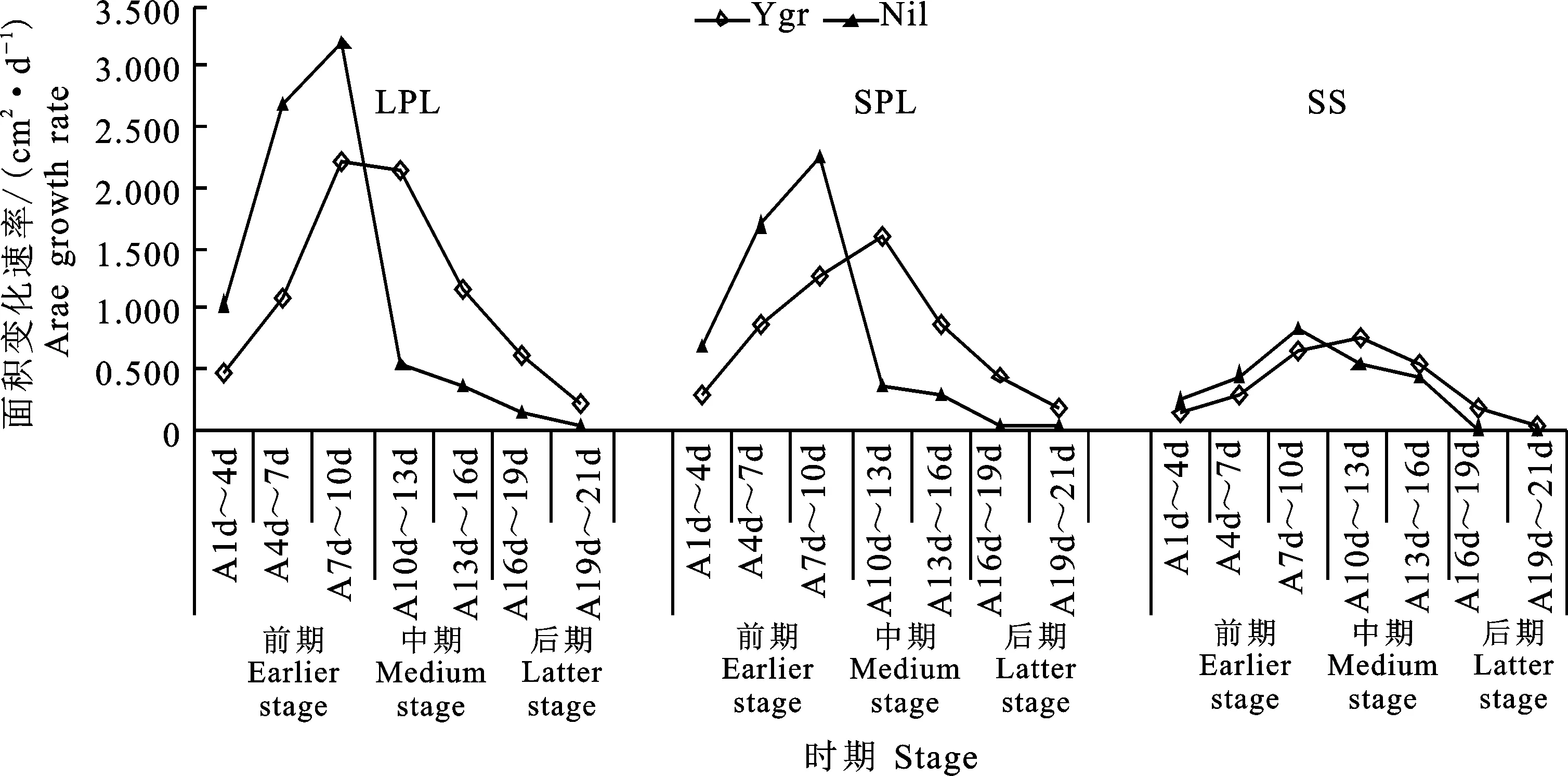
圖10 不同時期Ygr與Nil單葉面積和單個角果面積變化速率的比較Fig.10 Comparison of chang rates of single green leaf area and single of Ygr and Nil at different stages
2.4.3 單株綠葉面積和角果面積變化 從圖10可看出,Ygr與Nil的單株綠葉面積(Leaf area LA)的變化為“雙峰曲線”:兩者均在12月6日達(dá)到第1峰(分別為958.76、910.54 cm2);Nil在4月4日(2 080.53 cm2)、Ygr在4月18日 (2 061.51 cm2)分別達(dá)到第2峰。 Ygr與Nil單株角果面積(Silique area SA)的變化均表現(xiàn)為“單峰曲線”,兩者均在5月23日(分別為3 153.49、 3 176.11 cm2)達(dá)到峰值。
從圖11可看出,與Nil相比,Ygr的綠葉面積(LA)在10月28日~12月6日、2月28日~4月12日之間分別低5.03%~11.76%、5.33%~10.66%。Ygr與Nil間LA的差異在11月8日(11.76%)、3月29日(10.66%)相差較大。與Nil相比,Ygr的SA僅在4月5日、4月12日低6.92%、5.25%。
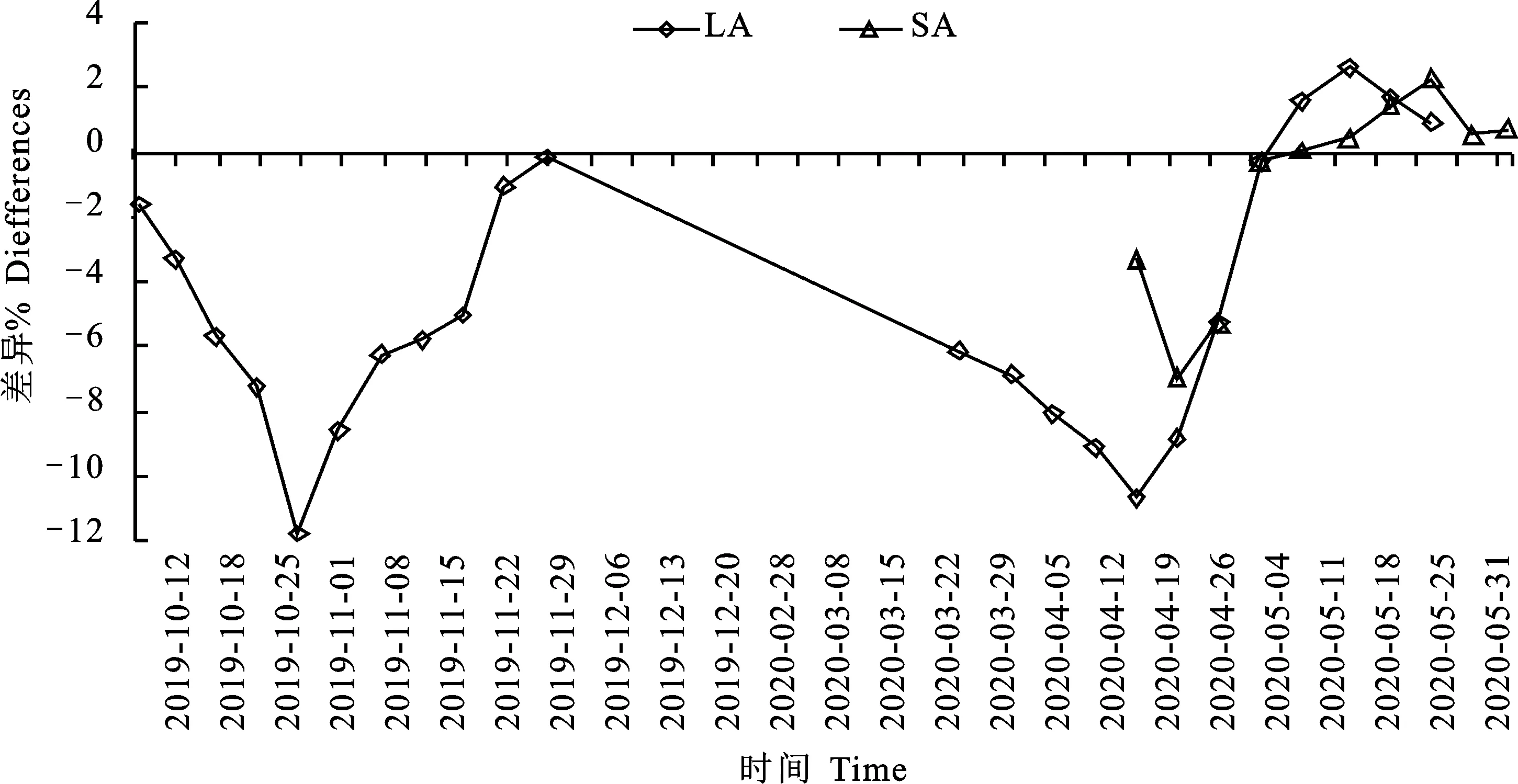
圖11 不同時間Ygr與Nil間單株綠葉面積和角果面積變化速率的比較Fig.11 Comparison of change rates of green leaf area and silique area per plant between Ygr and Nil at different time
2.5 農(nóng)藝性狀的比較
如圖11所示,與Nil相比,Ygr的葉片數(shù)(Leaf number LN)在10月25日-11月8日、3月8 -22日分別平均低6.47%、5.80%,角果數(shù)(Silique number SN)在3月29日低5.62%,株高(Plant height PH)在3月8日、3月15日分別低5.71%、5.46%,生物學(xué)產(chǎn)量(Biological yields BY)在10月25日-11月8日、11月29日-3月15日之間及4月19日分別平均低6.66%、 7.60%、5.76%。
從表3可看出,在成熟期與Nil相比,Ygr的分枝部位(BH)、主花序長度(MIL)低10.06%、3.47%而主花序角果數(shù)(SNMI)則高7.95%。
表3 Ygr和Nil成熟期主要農(nóng)藝性狀的比較Table 3 Comparison of main agronomic characters at maturity stage between Ygr and Nil
3 結(jié)論與討論
3.1 Ygr的黃化變異與油菜中其他黃化突變體的差異
與Nil相比,Ygr的5類光合器官(LPL、SPL、MSS、FB、SS)的4種光合色素(Chla、Chlb、Chl(a+b)、Car)含量均降低,且降低程度的大小為LPL>SPL>SS>FB>SS,表明Ygr的多個光合器官(葉片、莖稈、花蕾、角果)均會發(fā)生不同程度的黃化,這與對Cr3529[24]、NY[25]、L6382y[27]的研究結(jié)果相同。與Nil相比,Ygr不同的光合色素組分的降低程度大小為Chlb>Chl(a+b)>Chla>Car,Chla/b的比值高6.82%~42.02%、Chlb/(a+b) 的比值低4.73%~22.75%,表明Ygr在黃化期間Chla、Chlb不是同步降低,即Chlb的降低程度大于Chla,這與NY[25]、L638-y[27]及董遵等[23]的黃化突變體的研究結(jié)果相同。與Nil相比,Ygr的Chl(a+b)/Car的比值低 11.89%~25.49%,表明Ygr同Cr3529[24,26]、NY[25]、L638-y[27]等突變體相同,均屬于葉綠素總量減少型突變體。Ygr的LPL、SPL葉綠素含量、單葉葉(角果)面積與NIL達(dá)到無顯著差異的時間不同,即不同光合器官葉綠素含量降低程度的差異不同,表明不同光合器官的黃化程度不同,即表現(xiàn)為SPL>LPL>GSS>FB>PS。Ygr的5類光合器官中Chl(a+b)/類胡蘿卜素的比值顯著比Nil降低,而L638-y與其原始材料相比葉綠素(a+b)/類胡蘿卜素的比值變化不大[27],可說明Ygr的黃化變異原因與L638-y有差別。
3.2 黃化對Ygr光合特性的影響
通過對基本光合參數(shù)比較,發(fā)現(xiàn)Ygr葉片(LPL、SPL)的光飽和點、光補(bǔ)償點、光量子效率、光呼吸速率、CO2飽和點、CO2補(bǔ)償點、暗呼吸速率顯著高于Nil;角果(SS)的光量子效率、暗呼吸速率、CO2飽和點顯著高于Nil,而CO2飽和點、羧化效率則極顯著低于Nil。Ygr葉片(LPL、SPL)的初始熒光、PSⅡ調(diào)節(jié)性能量耗散的量子產(chǎn)額、非光化學(xué)猝滅系數(shù)顯著高于Nil,而PSⅡ潛在活性、PSⅡ中光化學(xué)能量轉(zhuǎn)化的有效量子產(chǎn)量、光化學(xué)猝滅系數(shù)、表觀電子傳遞速率則顯著低于Nil。Ygr角果(SS)的PSⅡ調(diào)節(jié)性能量耗散的量子產(chǎn)額顯著高于Nil。以上試驗結(jié)果表明黃化對Ygr葉片、角果的基本光合特性均產(chǎn)生了明顯影響。在飽和光強(qiáng)點、CO2飽和點以上與Nil相比,Ygr的3類光合器官(LPL、SPL、SS)的平均Pn值分別降低5.73%~22.75%、4.93%~ 16.76%,不僅可與對Ygr的葉綠素含量的比較結(jié)果相印證,也說明葉綠素含量降低是Ygr在黃化期間Pn降低的主要原因。同時,在光飽和點、CO2飽和點以上,Ygr、Nil不同光合器官間Pn值的大小、降低程度均為SPL>LPL>SS,則可能是由于SPL所生長時期(抽薹期)的環(huán)境條件(光照、溫度等)的優(yōu)于LPL的(苗期)、SS(角果期)的生長時期,有利于光合能力發(fā)揮。與Nil相比,Ygr的氣體交換參數(shù)中氣孔導(dǎo)度(Cond)、氣孔限制值(Ls)顯著較低,胞間CO2濃度(Ci)、蒸騰速率(Trm)顯著較高,其原因可能在于Ygr在葉綠素含量降低的同時,葉片的組織結(jié)構(gòu)(薄厚、氣孔的大小與多少)也發(fā)生了改變,進(jìn)而導(dǎo)致光合性能發(fā)生改變,具體原因尚待進(jìn)一步研究。
3.3 Ygr是一種新型油菜葉色突變系
目前,在油菜中已發(fā)現(xiàn)的葉色突變體中L638-y屬于芥菜型油菜[27],Ygr與Cr3529[24]、NY[25]及董遵等[23]的黃化突變體同屬于甘藍(lán)型油菜。董遵等[23]的黃化突變體是用氮離子處理、Cr3529是利用快中子和硫酸二乙酯(DES)復(fù)合處理[24]油菜種子而來,而Ygr和NY[25]則來源于自然突變。前人按照葉色突變性狀轉(zhuǎn)化的時期將轉(zhuǎn)綠型突變體分為在特定階段突變和在整個生育期突變有2種類型[2-3],Ygr的葉色變化與大豆中的NJ89-3[15]、NTV1-M、NTV2-M[16],水稻中的grc2[9]、ygr[10,31-32],棉花中的浙12-12N[20]、中58vsp[21-22],油菜中的Cr3529[24]、NY[25]、L638-y[27]等均屬于在整個生育期突變型;而董遵等[23]的黃化突變體因僅在苗期出現(xiàn)黃化(持續(xù)時間為 7~10 d),屬于階段性表達(dá)突變。Ygr的子葉為綠色,而油菜中Cr3529[24]、NY[25]、L6382y[27]的子葉均為黃色。
3.4 Ygr具有較高的研究和利用價值
葉色突變體是研究植物光合機(jī)理的理想材料[1-2,32-33],因此Ygr可用于研究油菜葉綠體生長發(fā)育和光合特性研究。在油菜的葉色突變體中Cr3529的株高、分枝數(shù)、單株角果數(shù)、角粒數(shù)、單株產(chǎn)量等性狀顯著降低[24],NY的生育期延長12 d、初花期推遲約14 d、花期總?cè)諗?shù)縮短約6 d、株高降低30 cm、莖總?cè)~數(shù)減少6片,有效角果數(shù)、每角粒數(shù)、千粒質(zhì)量和單株籽粒產(chǎn)量僅為正常株系(NG)的61.52%、68.35%、75.76%和 57.09%[25],因此將兩者直接用于育種或生產(chǎn)尚有較大困難。而Ygr的LPL、SPL的葉綠素含量在15 d、12 d后,單葉葉面積在18 d、15 d后即與Nil間無差異,且對生長發(fā)育進(jìn)程和農(nóng)藝性狀無明顯影響,因而具有較高的生產(chǎn)利用價值。可將Ygr的黃化的性狀作為指示性狀用于鑒定雜交種子的純度鑒定,也可利用Ygr的階段性黃化性狀培育觀賞油菜。
