H3K9me3對豬卵母細胞體外成熟和早期胚胎發育的影響
2022-03-26 07:48:08田釵張彩玉楊婷王婷黃時海石德順李湘萍
西南大學學報(自然科學版) 2022年3期
關鍵詞:實驗
田釵,張彩玉,楊婷,王婷,黃時海,石德順,李湘萍
廣西大學 亞熱帶農業生物資源保護與利用國家重點實驗室,南寧 530004
組蛋白修飾是最重要的表觀遺傳修飾標記之一,在控制基因表達和細胞譜系規范中起到關鍵作用[1-3].已有研究表明,組蛋白甲基化對早期胚胎發育和胚胎干細胞多能性的維持具有重要作用[4].組蛋白甲基化發生在組蛋白的賴氨酸和精氨酸殘基上,作用位點在其側鏈.
在N原子上,常見位點有H3K4me3,H3K9me3和H3K27me3[5-6].其中,組蛋白H3第9位賴氨酸三甲基化(Histone H3K9 trimethylation,H3K9me3)與維持異染色質穩定和沉默、X染色體失活、印記相關基因抑制及細胞特異性的鑒定有關[7],也被認為是胚胎發育的關鍵障礙[8].
H3K9me3被證明是小鼠[9-10]、人類[11]和牛[12]重新編程中關鍵的一個表觀遺傳調節因子.在小鼠GV卵母細胞中,H3K9me3的表達與組蛋白甲基轉移酶的活性密切相關.BHPF(Bateosi High Pass Filter)處理通過影響細胞骨架結構、線粒體功能、氧化應激、細胞凋亡提高了H3K9me3的表達,從而降低了小鼠卵母細胞的成熟效率和卵母細胞的質量[13].大劑量真菌毒素導致小鼠卵母細胞H3K9me3水平升高,這可能是導致卵母細胞發育能力下降的原因之一[14].以上研究表明,適當降低H3K9me3的表達有利于卵母細胞體外成熟和早期胚胎發育.
H3K9me3在基因表達調控和卵母細胞生長中起重要作用[15],目前其在體外成熟豬卵母細胞和孤雌胚胎中的表達模式還鮮有報道.毛殼素作為SUV39亞家族甲基轉移酶的抑制劑,能夠特異性抑制組蛋白甲基轉移酶SUV39H1/2和G9A的活性,通過競爭性結合SUV39H1/2[16],與關鍵殘基發生反應,從而調控H3K9me3水平,以此增強表觀遺傳重編程效率[17].因此,本實驗通過在豬卵母細胞體外成熟過程中添加毛殼素,調控卵母細胞H3K9me3及其相關基因的表達.研究結果有助于了解體外成熟卵母細胞和胚胎中H3K9me3表達的動態變化,并為改善豬卵母細胞成熟質量,提高胚胎發育效率奠定基礎.
1 材料與方法
1.1 材料和試劑
本實驗所用卵巢均來源于廣西南寧市屠宰場,實驗所用胚胎為孤雌激活胚胎.
1.2 豬卵巢的采集及體外成熟培養
將從屠宰場取得的卵巢置于37 ℃生理鹽水中,用生理鹽水清洗3遍,并用注射器采集卵巢表面直徑約3~10 mm的卵泡,挑選形態完好、胞質均勻,周圍帶有3層以上致密卵丘細胞的卵丘-卵母胞復合體(cumulus-oocyte complexes,COCs),用洗卵液清凈后將30~50個卵母細胞放入用礦物油覆蓋的一個胚胎培養液(PM)微滴中,每滴為150 μL培養液,[0~22)h將卵母細胞放入含有激素的培養液中進行培養,之后將卵母細胞移入另一個不含激素的培養液中繼續培養[22~44] h.實驗組在對照組的基礎上添加了不同濃度的毛殼素.
1.3 豬卵母細胞的孤雌激活
實驗前先打開熱臺、融合儀,并安裝激活盤.用提前平衡的電激活液清洗3遍融合槽,然后把新的電激活液放置于融合槽中.將卵母細胞在融合液中清洗3遍后放于激活槽中進行激活.將激活過的卵母細胞移入平衡好的胚胎培養液培養15 min,然后移入35 μL的胚胎培養液微滴中進行培養.24 h統計卵裂率,48 h統計4-細胞率,7 d統計囊胚率.
1.4 囊胚細胞總數的鑒定
將收集的豬孤雌囊胚放入4%的多聚甲醛固定液中固定24 h,然后放入10 μg/mL細胞核染料(Hoechst)中,避光染色10 min,然后用磷酸鹽緩沖液(PBS)清洗3遍,每次5 min,染好的囊胚移入載玻片的抗淬滅劑中,用凡士林封片,在熒光顯微鏡下觀察并做好統計.
1.5 細胞免疫熒光
收集不同時期的卵母細胞和孤雌囊胚放入4%多聚甲醛中4 ℃固定24 h.固定之后,室溫條件下放入阻斷液中阻斷3次,每次5 min,然后放入1% Triton X-100中,透化30 min,再用阻斷液阻斷2次,每次5 min,然后用1%牛血清蛋白溶液(BSA)進行非特異性位點封閉1~2 h,封閉之后用T-BSA-PBS清洗3次,每次5 min,把處理好的樣品放入H3K9me3的一抗4 ℃過夜.第2 d把樣品放入培養箱孵育30 min,然后用T-BSA-PBS清洗3遍,每次5 min,清洗好之后把樣品放入裝有二抗的EP管中,避光室溫孵育1.5 h,然后用清洗液清洗3遍,每次5 min,最后用10 μg/mL Hoechst 33342復染10 min,用清洗液清洗兩遍放入滴有抗淬滅劑的玻片上,用凡士林封片鏡檢,記錄實驗數據.
1.6 反轉錄和qRT-PCR
收集不同時期的豬卵母細胞和孤雌囊胚,用PBS+PVA清洗3遍,洗去樣品所帶的培養液和血清等物質后將樣品放入裝有細胞裂解液的管中.微量反轉錄所有步驟均在冰上操作:裂解,將樣品放入聚合酶鏈式反應(PCR)儀中4 ℃裂解15 min;消化,裂解后每管加入1 μL DNase I和1.3 μL 10×DNA buffer,瞬時離心后放入PCR儀中37 ℃消化40 min;終止,消化完每管加入1 μL EDTA,2 μM Random Primer,1 μL mixture deoxynucleotides (dNTPs),瞬時離心后放入PCR儀中65 ℃10 min;反轉,終止后依次加入2 μL DTT,4 μL 5×First-Strand Buffer,0.5 μL RNase Inhibitor(RRI),0.25 μL SuperScriptTMⅡ Reverse Transcriptase (SSⅡ),瞬時離心,進行反轉錄,其條件是25 ℃ 10 min,42 ℃ 90 min,95 ℃ 10 min,4 ℃停止.將反轉錄得到的樣品存放于-20 ℃備用.
將微量反轉錄得到的產物作為模板,進行qRT-PCR檢測,觀察目的基因的表達情況.反應體系按說明書進行.
1.7 結果統計與分析
本實驗結果均使用SPSS 22.0軟件進行分析,早期胚胎發育率先通過反正弦變化后再進行分析.每個實驗重復至少3次,每次重復實驗中至少隨機挑選10個以上卵母細胞進行實驗.p<0.05表示差異具有統計學意義.
2 結果與分析
2.1 [0~22)h毛殼素處理對豬卵母細胞體外成熟和早期胚胎發育的影響
[0~22)h采用不同濃度毛殼素處理,發現2 nmol/L處理組的卵母細胞第一極體排出率顯著高于未處理組、5 nmol/L組和10 nmol/L組;4-細胞率顯著高于未處理組、5 nmol/L組和10 nmol/L組;2 nmol/L處理組的囊胚率顯著高于未處理組和10 nmol/L組(p<0.05);2 nmol/L處理組的囊胚細胞總數顯著高于5 nmol/L組和10 nmol/L組(p<0.05),與未處理組之間差異不具有統計學意義(p>0.05)(表1).
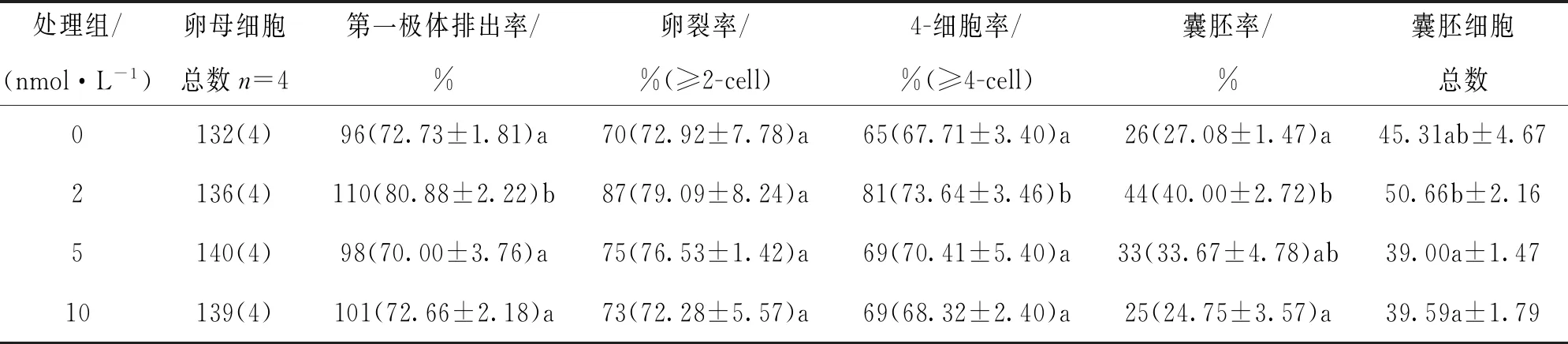
表1 毛殼素處理對豬卵母細胞體外成熟和早期胚胎發育的影響
2.2 [22~44] h毛殼素處理對豬卵母細胞體外成熟和早期胚胎發育的影響
[22~44] h采用不同濃度毛殼素處理時發現,與未處理組相比,5 nmol/L毛殼素處理組的第一極體排出率顯著提高(87.37% VS 71.91%,p<0.05),各處理組之間的卵裂率、4-細胞率、囊胚率和囊胚細胞總數差異不具有統計學意義(p>0.05)(表2).
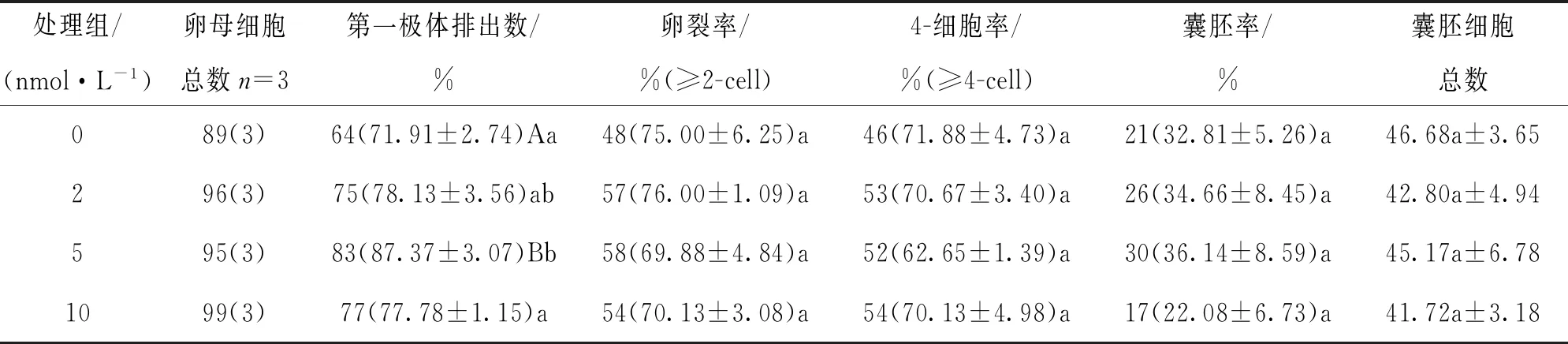
表2 [22~44] h毛殼素處理對豬卵母細胞體外成熟和早期胚胎發育的影響
基于以上實驗結果,后續實驗均采用體外成熟[0~22)h 添加2 nmol/L毛殼素處理方法.
2.3 毛殼素處理對豬卵母細胞和早期胚胎中H3K9me3表達的影響
與未處理組相比,H3K9me3在GV期2 nmol/L處理組有明顯的表達,差異不具有統計學意義(p>0.05).隨著卵母細胞的成熟進程,H3K9me3表達逐漸減弱,在MI和MⅡ期卵母細胞中未檢測到H3K9me3表達.與未處理組相比,2 nmol/L處理組的囊胚中H3K9me3表達顯著降低(p<0.05)(圖1、圖2).
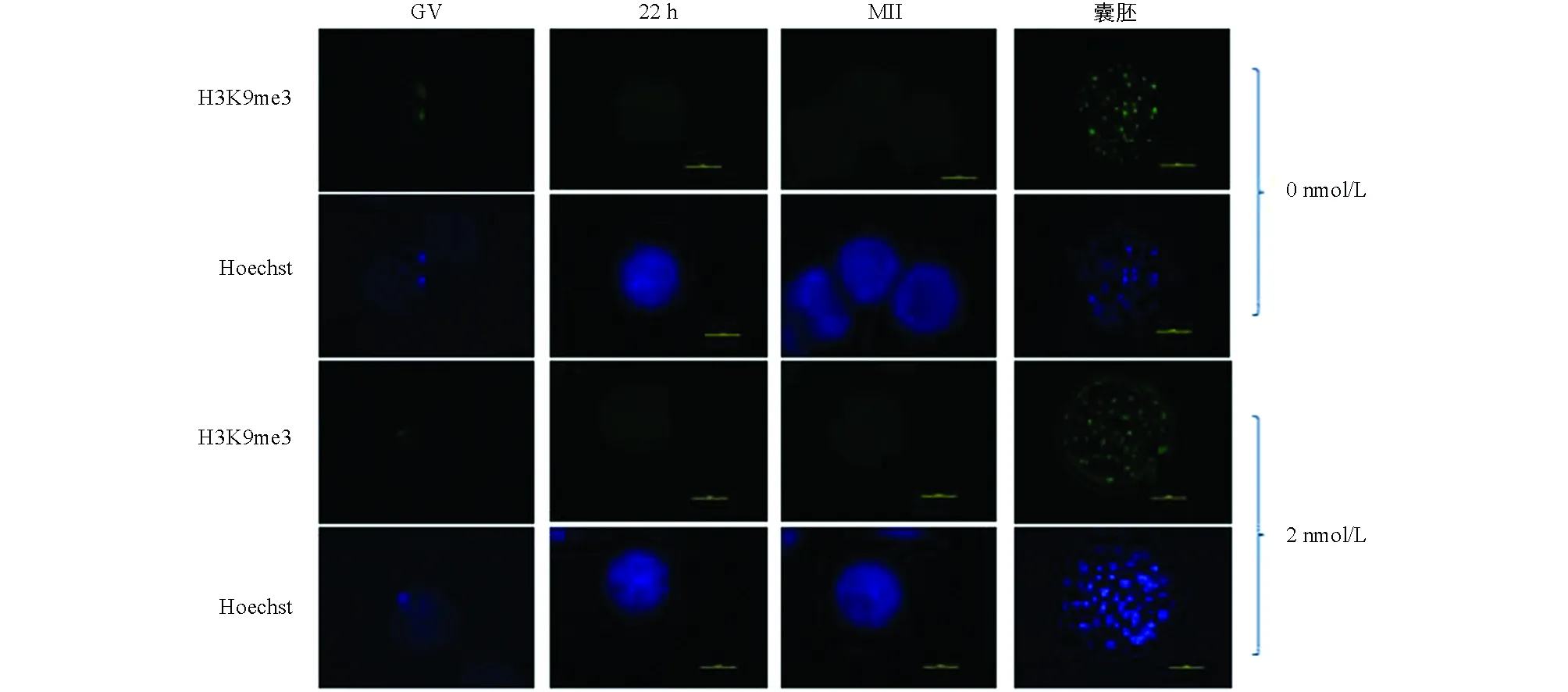
圖1 毛殼素處理對豬卵母細胞和囊胚中H3K9me3表達的影響
2.4 毛殼素處理對豬卵母細胞和早期胚胎中重要基因表達的影響
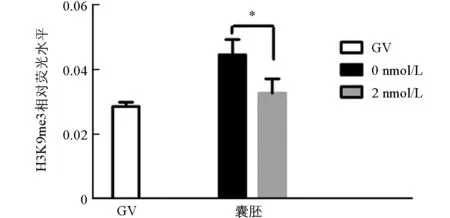
在相同時期各組之間*表示差異具有統計學意義(p<0.05).圖2 GV期豬卵母細胞和囊胚中H3K9me3的熒光強度分析
與未處理組相比,2 nmol/L處理組的MI期卵母細胞中的SUV39H1和SUV39H2表達顯著降低(p<0.05),兩組之間G9A的表達差異不具有統計學意義(p>0.05).與未處理組相比,2 nmol/L處理組的MⅡ期卵母細胞中SUV39H1,SUV39H2,G9A表達均顯著降低(p<0.05);囊胚中Nanog,Oct4,CDX2的表達顯著提高(p<0.05)(圖3).
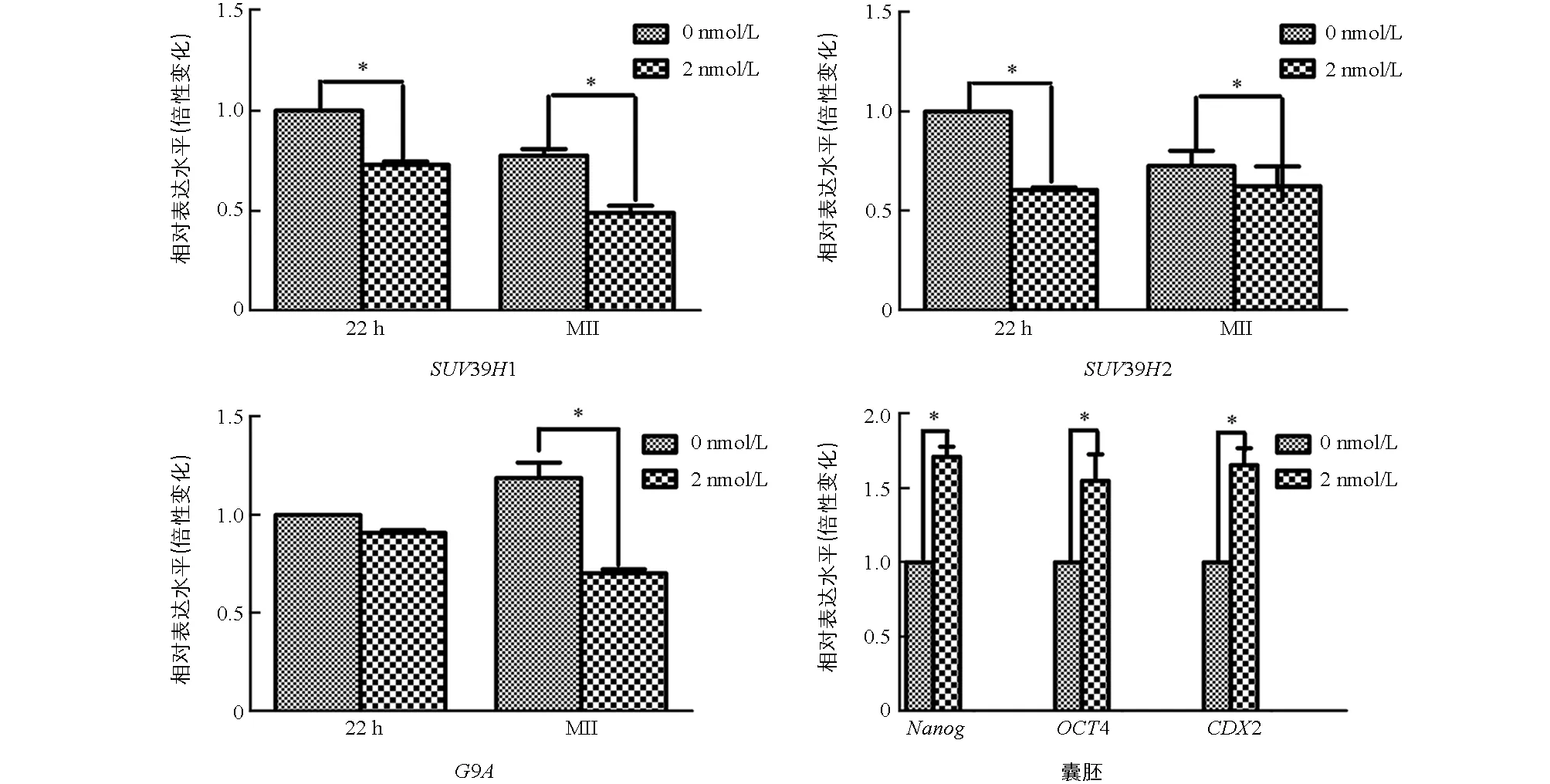
在相同時期各組之間*表示差異具有統計學意義(p<0.05).圖3 不同處理豬卵母細胞和早期胚胎中相關基因的表達
3 討論與結論
近年來的研究表明,胚胎中高表達的H3K9me3是阻礙胚胎發育的關鍵因素,特別是在合子基因激活(ZGA)過程中[18].在ZGA期間,卵母細胞中H3K9me3的異常變化會阻礙轉錄激活[19],且在核移植胚胎重編程抗性區域(RRR)中檢測到高水平的H3K9me3,而體外受精胚胎的RRR基因表達正常.這些RRRs區域的特征是富含SUV39H1/2并且缺乏DNasr1,這兩點都是異染色質的一般特征[20].由于H3K9的甲基化與基因的表達抑制密切相關,去除這些表觀遺傳標記有利于基因的重新激活和胚胎發育.以往的研究也表明,降低人和小鼠胚胎中H3K9me3水平能提高胚胎的發育能力[10].因此,適當降低H3K9me3的表達有利于卵母細胞體外成熟和早期胚胎的發育.
在本研究中,免疫熒光數據顯示GV期卵母細胞中H3K9me3的表達水平較高,但MI期和MⅡ期卵母細胞中未檢測到該蛋白的表達,發育到囊胚階段又表現出較高的表達水平,這意味著MII期卵母細胞中H3K9me3的缺失可能正在為通過其他修飾的細胞表觀遺傳重新編程做準備[21].H3K9me3的動態表達說明了其在卵母細胞體外成熟和早期胚胎發育過程中具有重要的調控作用.
本實驗通過在豬卵母細胞體外成熟過程中添加毛殼素,觀察其是否對豬卵母細胞體外成熟和早期胚胎發育產生影響.結果發現,與未處理組相比,2 nmol/L處理組顯著提高了卵母細胞的第一極體排出率、4-細胞率和囊胚率.在本研究中2 nmol/L處理組囊胚率增加12.92%,表明抑制劑只部分糾正了異常的表觀遺傳修飾.通過qRT-PCR和免疫熒光檢測發現,毛殼素處理之后顯著降低了卵母細胞中SUV39H1/2和G9A的表達,以及囊胚中H3K9me3的表達.這與Zhang等[22]在羊核移植胚胎中的研究一致.2 nmol/L毛殼素處理后顯著提高囊胚中多能性基因Nanog,Oct4和CDX2的表達,該結果與李海艷[23]對豬孤雌胚胎的研究結果一致.本實驗結果證明適量毛殼素處理有利于豬卵母細胞體外成熟和早期胚胎的發育.
豬卵母細胞中H3K9me3的去除對于正常的ZGA和表觀遺傳重編程至關重要[18],用毛殼素作為組蛋白甲基轉移酶SUV39H1/2和G9A特異性抑制劑,能有效降低H3K9me3的表達水平,最終提高豬體外成熟和胚胎發育效率.
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
